Scolytinae
Les scolytes (Scolytinae) forment une sous-famille d'insectes coléoptères[1] de la famille des Curculionidae.
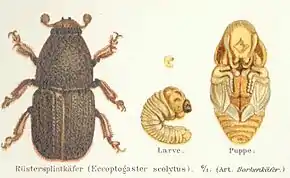
Description

Les scolytes sont de petits insectes de l'ordre des coléoptères. Ils font partie d'une grande famille renfermant des insectes dits « ravageurs », les curculionidés. Leur corps de 2 à 5 mm est cylindrique et court, de couleur brun foncé à rougeâtre et recouvert par les élytres qui protègent leurs ailes. Leur nom vient du grec skôlêx, terme des auteurs antiques désignant un ver amis aussi une larve d'insecte, celle des scolytes étant caractéristique (larve apode, molle, blanchâtre et arquée)[2].
Certaines espèces de scolytes (par exemple : Dendroctonus ponderosae) sont à l'état adulte attirées par certaines hormones de stress (phytohormones) émises par des arbres malades ou déshydratés[3], les autres le sont par l'odeur du bois mort.
Sur les quelque 6 000 espèces connues dans 247 genres, la plupart sont phytophages, se nourrissant principalement ou uniquement sur des angiospermes. Pas plus de 10 % sont xylophages (les femelles pondent sous l'écorce des arbres et les larves se nourrissent de leur sève, ce qui peut mener à la mort de l'arbre, moins de 1 % de ces espèces pouvant tuer des arbres vivants et sains)[4].
Comme beaucoup d'insectes xylophages ou saproxylophages (comme les fourmis et les termites), le scolyte ne peut directement digérer le bois en raison de la présence de lignine et de cellulose. Il emporte donc avec lui, de l'arbre où il est né, une provision de spores et/ou de mycélium d'un champignon symbiotique capable de digérer ces polymères. Le transport se fait grâce aux mycangia (sing. mycangium), invaginations de la cuticule souvent garnie de glandes qui semblent nourrir et/ou préserver les spores ou fragments de mycélium. Des spores de champignons peuvent être aussi captées, transportées et inoculées grâce aux poils microscopiques qui garnissent la carapace et les pattes du scolyte[5]. Ce champignon symbiote attaque la lignine et la cellulose, qui sont ainsi comestibles pour la larve du scolyte. Chez certaines espèces de Scolytinae et chez les Platypodinae, ce ne sont pas les fibres végétales prédigérées qui sont ingérées par la larve, mais plutôt le champignon lui-même. Certaines espèces de scolytes (e.g. Dendroctonus frontalis) n'ont pas de mycangium, mais elles transportent sur leur corps des acariens qui eux possèdent une sorte de mycangium (qu'on appelle sporothèque chez les acariens pour des raisons historiques de vocabulaire taxonomique).
De nombreuses espèces de scolytes sont, comme les hyménoptères, haplodiploïdes : les femelles sont diploïdes (provenant d'œufs fécondés) et les mâles haploïdes (provenant d'œufs non fécondés et ils sont donc parthénogénétiques).
Système de galeries
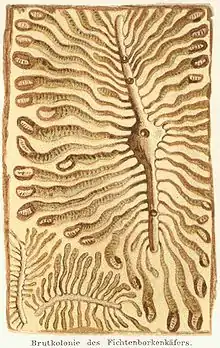
Ce sont généralement les femelles qui, grâce à leurs mandibules en forme de gouge, forent les orifices d'entrée dans l'écorce d'arbres de ponte, les mâles collaborant en dégageant les débris. Ils produisent des phéromones agrégatives, qui attirent des congénères des deux sexes. Le taraudage de la galerie de pénétration forme un canal d'abord radial ou oblique, avant de s'infléchir pour suivre une zone de moindre résistance (courbure d'un cerne de croissance). La littérature scientifique décrit aussi le forage de galerie alimentaire ou de galerie de maturation. Pour se reproduite, les mâles, parfois aidés des femelles, forent une chambre nuptiale. Lors d'une colonisation réussie, une ou plusieurs femelles (selon que l'espèce est monogame ou polygame) pénètrent dans la chambre nuptiale aménagée où se déroule l'accouplement avec le mâle. Depuis la chambre nuptiale, chaque femelle fore une galerie maternelle (appelée aussi galerie de ponte) de longueur variable (généralement quelques centimètres) dans le sens des fibres du bois (verticalement si l’arbre est encore debout), ce qui donne plusieurs branches partant de la chambre nuptiale (le nombre de branches correspondant au nombre de femelles fécondées). Elle y aménage des trous d'aération et dépose ses œufs (d'une dizaine à plusieurs centaines selon l'espèce) dans des encoches de ponte de part et d'autre de la galerie. Dès l'éclosion, la larve creuse à partir de l'encoche une galerie légèrement sinueuse (appelée galerie larvaire) qui s'élargit progressivement jusqu'à son extrémité où elle forme une niche appelée berceau de nymphose. Après la mue de la larve en imago, le jeune scolyte poursuit le forage cortical en creusant de nombreux couloirs et perçant un trou dans l'écorce pour partir essaimer. L'essence de l'arbre, la forme et la taille des galeries, permettent à un œil averti de déterminer l'espèce de scolyte responsable. À mesure que la densité des galeries augmente, une phase de dépérissement des arbres commence. L'écorce perd alors ses capacités de défense physique et chimique. Elle se détache sous l'effet de ces forages larvaires et finit par tomber, révélant le réseau de galeries qui dessinent de grands motifs rappelant des graffitis sous l'écorce[6].
Dégâts


D'autres espèces de scolytes ou d'insectes xylophages peuvent creuser plus profondément le bois, favorisant la pénétration de champignons qui le dégradent plus rapidement (certains scolytes ne consomment que le bois d'arbres tombés).

En temps normal, le scolyte joue un rôle utile pour la régénération forestière. Dans un cas particulier, des scolytes ont colporté d'orme en orme un nouveau variant — extrêmement phytopathogène — du champignon Ophiostoma ulmi ; ce champignon ayant peut-être été favorisé par la forte sécheresse de 1975-1976 et par l'homogénéité génétique des ormes d'alignement et du bocage.
Un arbre en bonne santé dispose pour se défendre d'une batterie de molécules naturellement insecticides, bactéricides et fongicides et de défenses physiques (lignine dure, capacité à immobiliser, engluer et noyer dans la sève ou de la résine tout insecte s'introduisant ou se développant dans la partie superficielle de son tronc). Ce n'est plus le cas chez un arbre fendu, blessé ou déshydraté.
Des successions de canicules ou d'étés chauds et secs suivant des hivers anormalement doux semblent avoir favorisé — dans tout l'hémisphère nord — des pullulations de défoliateurs et de scolytes, que les monocultures équiennes de résineux semblent aussi favoriser.
Après de grandes tempêtes ou une sécheresse anormalement longue et/ou importante, le scolyte peut se reproduire massivement sur les chablis, les arbres déshydratés, mutilés, blessés ou affaiblis. Les galeries de ces scolytes sont réputées être autant de « portes d'entrée » à d'autres organismes saproxyliques (qui consomment le bois mort ou vivant). Les scolytes ont longtemps été considérés comme des « nuisibles ». Pourtant, lors des années de sécheresse intense, parfois après une ou plusieurs vagues de défoliation par des insectes défoliateurs (également attirés par les hormones de stress émises par les arbres), les scolytes jouent un rôle majeur dans la résilience écologique de la forêt.
Si d'autres insectes ont d'abord défolié, éventuellement plusieurs années de suite, les arbres les plus fragiles ou incapables de diminuer leur évapotranspiration, bloquant leur photosynthèse et leur croissance, les scolytes attirés par les hormones de stress de l'arbre vont, eux, les tuer si la sécheresse s'aggrave ou se prolonge. En contribuant à tuer les arbres qui sont en état de stress hydrique aigu, des scolytes tels que Dendroctonus ponderosae jouent un rôle important dans l'écosystème forestier, en supprimant l'évapotranspiration et la respiration de ces arbres. Les scolytes limitent ainsi le pompage de l'eau dans le sol et la nappe superficielle par les arbres adultes, au profit des graines et des jeunes plants, tout en diminuant le risque d'incendie. Les scolytes accélèrent ensuite fortement la décomposition du bois mort, notamment de résineux, accélérant ainsi la formation d'un humus forestier favorable à la pousse des futurs arbres et à une meilleure rétention de l'eau dans le sol[réf. nécessaire].
Néanmoins, c'est surtout le risque phytosanitaire et la perte apparente de revenus que voit le sylviculteur ou le propriétaire forestier qui craignent toujours une pullulation d'insectes mangeurs de bois et/ou des champignons qui les accompagnent, pouvant engendrer une catastrophe plus étendue et plus dommageable que la tempête elle-même. Ces atteintes suivent souvent les périodes de fortes sécheresses ou de sécheresses répétées, et elles peuvent accroître de 30 % le volume d'arbres, morts ou mourants, renversés par les tempêtes qui suivront, et jusqu'à 50 voir localement 100 % si le vent est très violent[réf. nécessaire].
Une espèce de scolyte, Tomicus piniperda, a un imago qui achève sa maturation en se nourrissant non pas dans l'écorce de l'arbre où il a effectué son développement larvaire, mais dans la moelle des pousses de pins, en limitant l'efficacité de la régénération.
Des programmes d'incendies contrôlés sont encouragés au Canada et localement obligatoires en Suède[réf. nécessaire]. Paradoxalement, la lutte contre les incendies a favorisé à la fois les scolytes et des incendies de plus en plus difficiles à maîtriser.
 Traces de galeries de scolytes, Saint-Guilhem-le-Désert, Hérault, France.
Traces de galeries de scolytes, Saint-Guilhem-le-Désert, Hérault, France. Traces de galeries, avec quelques trous de sortie.
Traces de galeries, avec quelques trous de sortie. Certaines espèces pénètrent plus profondément le bois mort, avec leur champignon symbiote, accélérant la décomposition du bois.
Certaines espèces pénètrent plus profondément le bois mort, avec leur champignon symbiote, accélérant la décomposition du bois. Bleuissement du bois causé par des champignons transmis par des scolytes.
Bleuissement du bois causé par des champignons transmis par des scolytes.
 Un variant exceptionnellement pathogène d'un champignon transporté par des scolytes a décimé les ormes à plusieurs reprises dans l'hémisphère nord.
Un variant exceptionnellement pathogène d'un champignon transporté par des scolytes a décimé les ormes à plusieurs reprises dans l'hémisphère nord. Sur la presqu'île de Crozon.
Sur la presqu'île de Crozon. Forte régénération naturelle après sécheresse de 1976 puis mortalité par les scolytes.
Forte régénération naturelle après sécheresse de 1976 puis mortalité par les scolytes.
Moyens de lutte


La lutte contre les scolytes est délicate, d'abord du fait de leur mode de vie : ils passent en effet la plus grande part de leur vie sous l'écorce (où on leur connaît peu de prédateurs, hormis des bactéries ou acariens qui peuvent attaquer ses œufs[11]) ; ensuite parce qu'en les éliminant (par des insecticides), on permettrait aux arbres stressés de survivre, c'est-à-dire d'évapotranspirer plus longtemps, en continuant donc à épuiser la ressource en eau en temps de sécheresse (risque accru de maladies et de défoliation plus grave, mais aussi d'incendies aggravés).
Plusieurs modes de lutte sont possibles :
- Lutte préventive : elle vise à ne pas encourager la pullulation de scolytes ; elle visera surtout à limiter tant que faire se peut le stress hydrique des arbres qui semble être une condition de pullulation du scolyte. Selon une étude slovène, l'application d'une dose précise, et dans certaines conditions de deux couches d'acide salicylique semble aussi renforcer les défenses immunitaires de l'arbre[12].
- Limitation des « effets de lisières » et donc de la fragmentation des forêts par des routes et grandes coupes rases uniformes ; on a par exemple constaté que les épicéas poussant sur les lisières naturelles (falaises) ou artificielles (de coupes rases, routes, layons...) ou sur des zones sèches, notamment sur pentes, semblent présenter une sensibilité exacerbée au stress hydrique et aux scolytes et peut-être à des infestations par des défoliateurs (les perturbations micro-climatiques liées aux effets de lisières sont mesurables avec un simple thermo-hygromètre et visibles en photo-infrarouges. Dans ces zones, les attaques de scolytes semblent favorisées, selon des études qui demandent encore à être affinées[13].
- Meilleure protection du sol, et si nécessaire sa restauration.
- Meilleure gestion de l'eau, de manière à mieux la retenir en forêt, dès le haut des bassins-versant (pour limiter l'impact des sécheresses), alors que depuis le Moyen Âge on a surtout drainé et asséché les forêts.
- Biodiversité élevée des arbres, ce qui implique l'usage de sujets issus de souches locales et adaptées au substrat, nés de graines et non clonés ou bouturés) et leur plantation en mélange. Il semble qu'elle soit favorable à moins de pullulations et à une meilleure résilience, mais de nombreuses inconnues persistent en raison du réchauffement climatique attendu ; dans les forêts polonaises et bélarusses de Białowieskiej, les pullulations semblent toutes corrélées à des stress climatiques, mais elles restent localisées.
- L'écorçage des grumes' abattues, ou des arbres tombés après une tempête, permet d'éviter que des scolytes ne s'y installent et y pondent, leurs larves ne pouvant en effet vivre et grandir que sous l'écorce d'arbres fraîchement abattus ou tombés. Les adultes issus de ces larves pourront alors ensuite s'attaquer à des arbres proches et assez affaiblis pour y être vulnérables. En écorçant les grumes après abattage ou à la suite d'une tempête, on évite ou au moins on limite le développement des populations de scolytes.
- Le débardage des arbres qui sont tombés en grande quantité, après une tempête ou des chutes de neige par exemple.
- Le piégeage des adultes. Les adultes sont en effet attirés par les odeurs des arbres auxquels ils sont attachés (et en particulier des arbres malades ou en déficience physiologique, par exemple du fait d'une sécheresse). On peut donc attirer les adultes vers des pièges à phéromones ou qui reproduisent le spectre d'odeurs d'arbres malades. Mais, compte tenu du nombre très important d'insectes pouvant sortir de chaque arbre (jusqu'à plusieurs dizaines de milliers), et du nombre beaucoup plus restreint capté par ces pièges, cette méthode s'avère coûteuse car elle demanderait la pose de plusieurs pièges par arbre pour être efficace - ce qui est inenvisageable en forêt. En revanche, un tel piégeage peut être utilisé pour tenter de réduire les populations dans les parcs ou dans les vergers ou pour détecter un éventuel début de « pullulation ».
Quelques espèces de scolytes
.jpg.webp)
- Alniphagus aspericollis ou scolyte de l'aulne
- Bostrychus capucinus ou bostryche capucin. Élytres rouges et tête noire.
- Conophthorus resinosae Hopkins ou scolyte des cônes du pin rouge.
- Dryocoetes betulae ou scolyte brun du bouleau
- Hylesinus californicus ou scolyte du frêne de l'ouest, présent dans la moitié occidentale de l'Amérique du Nord sur les frênes[14].
- Hylurgopinus rufipes ou scolyte américain de l'orme, originaire d'Amérique du Nord.
- Hypothenemus hampei Ferr. (syn. Stephanoderes hampei) ou scolyte des cerises du caféier, originaire d'Afrique, mais présent dans la plupart des régions tropicales.
- Ips cembrae ou grand scolyte du mélèze, en Europe.
- Ips typographus ou bostryche typographe, ou scolyte de l'épicéa.
- Ips sexdentatus ou sténographe.
- Myelophilus piniperda ou scolyte du pin.
- Phloeosinus canadensis ou scolyte du thuya de l'est, en Amérique du Nord, attaque les thuyas et les genévriers.
- Pityogenes chalcographus ou chalcographe, présent dans le houppier des vieux épicéas, en Europe.
- Pityokteines curvidens ou scolyte curvidenté, dans les sapins blancs, en Europe.
- Ruguloscolytus amygdali ou scolyte de l'amandier.
- Ruguloscolytus rugulosus ou petit scolyte des arbres fruitiers.
- Scolytus mali ou scolyte du pommier ou bostryche disparate, attaque les arbres fruitiers et la vigne, en Europe.
- Scolytus multistriatus ou petit scolyte de l'orme, originaire d'Europe.
- Scolytus rugulosus ou scolyte rugueux ou petit scolyte des arbres fruitiers en Amérique du Nord.
- Scolytus scolytus ou grand scolyte de l'orme, vecteur de la graphiose, maladie qui a décimé les ormes d'Europe.
- Taphrorychus bicolor ou petit scolyte du hêtre.
- Tomicus piniperda ou blastophage du pin
- Trypodendron lineatum ou scolyte birayé, se reproduit normalement dans les conifères morts ou moribonds de l'Amérique du Nord, mais attaque aussi les érables et les aubiers[15].
- Xyleborus dispar ou xylébore disparate.
- Xyleborus saxeseni ou petit bostryche.
- Xylosandrus germanus ou bostryche noir du Japon.
Liste des tribus
Amphiscolytini - Bothrosternini - Cactopinini - Carphodicticini - Coptonotini - Corthylini - Cryphalini - Crypturgini - Diamerini - Dryocoetini - Hexacolini - Hylastini - Hylesinini - Hylurgini - Hyorrhynchini - Hypoborini - Ipini - Micracidini - Phloeosinini - Phloeotribini - Phrixosomatini - Polygraphini - Premnobiini - Scolytini - Scolytoplatypodini - Xyleborini - Xyloctonini - Xyloterini - †Cylindrobrotini
Liste des genres
- Alniphagus Swaine, 1918.
- Ambrosiodmus Hopkins, 1915.
- Araptus Eichhoff, 1878.
- Cactopinus Schwarz, 1899.
- Carphobius Blackman, 1943.
- Carphoborus Eichhoff, 1864.
- Chaetophloeus Leconte, 1876.
- Chramesus LeConte, 1868.
- Cnemonyx Eichhoff, 1868.
- Cnesinus LeConte, 1868.
- Coccotrypes Eichhoff, 1878.
- Conophthorus Hopkins, 1915.
- Corthylus Erichson, 1836.
- Cryphalus Erichson, 1836.
- Cryptocarenus Eggers, 1937.
- Crypturgus Erichson, 1836.
- Dendrocranulus Schedl, 1937.
- Dendroctonus Erichson, 1836.
- Dendrosinus Chapuis, 1869.
- Dendroterus Blandford, 1904.
- Dolurgus Eichhoff, 1868.
- Dryocoetes Eichhoff, 1864.
- Ericryphalus.
- Ernoporicus Berger, 1917.
- Euwallacea.
- Gnathotrichus Eichhoff, 1869.
- Hylastes Erichson, 1836.
- Hylastinus Bedel, 1888.
- Hylesinus Fabricius, 1801.
- Hylocurus Eichhoff, 1872.
- Hylurgopinus Swaine, 1918.
- Hylurgops LeConte, 1876.
- Hypocryphalus Hopkins, 1915.
- Hypothenemus Westwood, 1836.
- Ips De Geer, 1775.
- Liparthrum Wollaston, 1864.
- Lymantor Lovendal, 1889.
- Micracis LeConte, 1868.
- Micracisella Blackman, 1928.
- Monarthrum Kirsch, 1866.
- Orthotomicus Ferrari, 1867.
- Pagiocerus Eichhoff, 1868.
- Phloeosinus Chapuis, 1869.
- Phloeotribus Latreille, 1804.
- Pityoborus Blackman, 1922.
- Pityogenes Bedel, 1888.
- Pityokteines Fuchs, 1911.
- Pityophthorus Eichhoff, 1864.
- Pityotrichus Wood, 1962.
- Polygraphus Erichson, 1836.
- Premnobius Eichhoff, 1878.
- Procryphalus Hopkins, 1915.
- Pseudohylesianus Swaine, 1917.
- Pseudohylesinus Swaine, 1917.
- Pseudopityophthorus Swaine, 1918.
- Pseudothysanoes Blackman, 1920.
- Ptilopodius.
- Pycnarthrum Eichhoff, 1878.
- Scierus LeConte, 1876.
- Scolytodes Ferrari, 1867.
- Scolytogenes Eichhoff, 1878.
- Scolytus Geoffroy, 1762.
- Stenocleptus Blackman, 1943.
- Thysanoes LeConte, 1876.
- Tomicus Latreille 1803.
- Trischidias Hopkins, 1915.
- Trypodendron Stephens, 1830.
- Trypophloeus Fairmaire, 1868.
- Xyleborinus Reitter, 1913.
- Xyleborus Eichhoff, 1864.
- Xylechinus Chapuis, 1869.
- Xylosandrus Reitter, 1913.
- Xyloterinus Swaine, 1918.
Notes et références
- Insectes dont une des deux paires d'ailes est transformée en élytres protecteurs, comme les scarabées, les coccinelles, etc.
- (en) Ray F. Smith, Thomas E. Mittler, Carroll Newton Smith, History of Entomology, Annual Reviews, , p. 4.
- Source : Wikipedia anglais.
- (en) Lawrence R. Kirkendall, Peter H.W. Biedermann, Bjarte H.Jordal, « Evolution and diversity of bark and ambrosia beetles », dans Fernando E. Vega , Richard W. Hofstetter, Bark Beetles : Biology and Ecology of Native and Invasive Species, Elsevier Science, , p. 85-156.
- Francke-Grossmann, H. (1967). « Ectosymbiosis in wood inhabitinginsects ». In: M. Henry (ed.) Simbiosis, Vol. 2. Academic Press, New York. p. 141-205.
- Vincent Albouy, Eric Darrouzet, Architectes du monde animal, Quæ, , p. 43-44.
- Appelé Scolyte liseré ou Scolyte birayé.
- Appelé Scolyte liseré ou Scolyte birayé.
- Tanins de l'écorce externe, reliés de façons différentes aux composants de la subérine. Cf Marc-André Selosse, Les Goûts et les couleurs du monde. Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud Nature, , p. 222
- Tanins de l'écorce externe, reliés de façons différentes aux composants de la subérine. Cf Marc-André Selosse, Les Goûts et les couleurs du monde. Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud Nature, , p. 222
- (de) Article suisse allémanique sur l'importance de la température pour le développement du scolyte, avec allusion aux acariens prédateurs de ses œufs.
- Krajnc A.U., Kristl J., Ivancic A. [2011]. Application of salicylic acid induces antioxidant defense responses in the phloem of Picea abies and unhibits colonization by Ips typographus. Forest Ecology and Management 261(3) : 416-426 (11 p., 6 fig., 73 réf.).
- « Integrated risk assessment and new pest management technology in ecosystems affected by forest decline and bark beetle outbreaks ». Programme TATRY ; IC15-CT98-0151 (Voir photos infrarouges et légendes (en bas de page) (en).
- « Scolyte du frêne de l'Ouest », Ressources naturelles Canada (consulté le ).
- « Scolyte rayé », Ressources naturelles Canada (consulté le ).
Voir aussi
Bibliographie
- (en) Jakus, R., « A method for the protection of spruce stands against Ips typographus by the use of barriers of pheromone traps in north-eastern Slovakia », 1998, Anz. Schädlingskde., Pflanzenschutz, Umweltschutz, 71, 152-158
- Beaulieu, Marie-Ève., « Caractérisation moléculaire des champignons ophiostomatoïdes associés à quatre espèces de scolytes de l'écorce colonisant l'épinette blanche au Québec et phylogénie multigénique d'une nouvelle espèce de Leptographium », , université Laval (télécharger)
- Balachowsky A., Coléoptères Scolytides, 1949, 320 p., 345 fig. (faunedefrance.org)
- Warzée N. & Grégoire J.C., « Biodiversité forestière et ennemis naturels des scolytes - Le cas exemplaire de Thanasimus formicarius », 2003, Forêt wallonne n° 66
- (en) Fernando E. Vega, Richard W. Hofstetter, Bark beetles. Biology and ecology of native and invasive species, Elsevier Science, , 640 p. (lire en ligne)
Articles connexes
- Scolytes de l'orme
 : différentes espèces.
: différentes espèces. - Crise des scolytes, épiphytie de Scolytinae dans les forêts d'épicéas communs du Nord-Est de la France à partir de 2018.
Liens externes
- (en) Référence BioLib : Scolytinae Latreille, 1804
- (en) Référence Fauna Europaea : Scolytinae (consulté le )
- (fr+en) Référence ITIS : Scolytinae Latreille, 1804
- (en) Référence NCBI : Scolytinae (taxons inclus)
- (fr+en) Référence EOL : Scolytinae Latreille 1804 : liste de genres
 Portail du bois et de la forêt
Portail du bois et de la forêt  Portail de l’entomologie
Portail de l’entomologie  Portail de la protection des cultures
Portail de la protection des cultures