Tétrachromatisme
Le tétrachromatisme (ou tétrachromatie) est le traitement de l'information visuelle sur la couleur à l'aide de quatre canaux et quatre types de cônes photorécepteurs. Les organismes qui procèdent ainsi sont appelés tétrachromates.
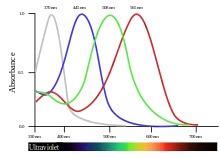
L'espace colorimétrique des organismes tétrachromates possède quatre dimensions. Par conséquent, la sensation spécifique à une onde électromagnétique donnée est susceptible d'être reconstituée à partir d'un mélange de quatre couleurs primaires (minimum). La plupart des oiseaux sont tétrachromates[2]. Plusieurs espèces de poissons, d'amphibiens, de reptiles, d'aranéides et d'insectes sont suspectés d'être tétrachromates[réf. nécessaire].
Physiologie
Le tétrachromatisme repose sur une palette de quatre sortes de photorécepteurs dans la rétine, dont la caractéristique est d'être stimulés par une intensité lumineuse plutôt élevée. Ces photorécepteurs sont connus sous le nom de cônes chez les vertébrés. On les différencie des bâtonnets, qui ont la caractéristique d'être stimulés en faible lumière. Chaque sorte de cônes a un spectre d'absorption propre. L'animal tétrachromate est susceptible de percevoir des longueurs d'onde que les humains (trichromates) ne voient pas, et pourrait être capable de distinguer des nuances de couleurs qui sembleraient identiques à un humain. Les espèces tétrachromates possèdent un léger avantage physiologique sur leurs espèces rivales[3].
Exemples
- Poissons
Le poisson zèbre (Danio rerio) est un exemple de tétrachromate, dont l’œil contient des cônes sensibles à la lumière rouge, verte, bleue et aux ultraviolets[4].
- Oiseaux
Certaines espèces d'oiseaux dont le Diamant mandarin fait partie ou les colombidés, se servent des ultraviolets (300 nm ≤ λ ≤ 400 nm) lors de la sélection des reproducteurs pour l'accouplement et lors de la recherche de nourriture[5]. Au cours de la sélection des reproducteurs, un plumage et une coloration de peau ultraviolets signalent un haut degré de sélection[6]. Typiquement, l’œil d'un oiseau est stimulé par des longueurs d'onde comprises entre 300 et 750 nm. Cela correspond à une plage de fréquences qui s'étend de 400 à 1000 THz.
- Insectes
Les insectes butineurs perçoivent les longueurs d'onde que les fleurs reflètent (de 300 nm à 700 nm[7])[réf. nécessaire]. La pollinisation est une relation mutualisée qui lie l'évolution des plantes à celle des insectes et inversement. L'une des conséquences de cette coévolution est d'avoir élargi le spectre visible des butineurs, ainsi que la gamme et les combinaisons de longueurs d'onde que reflètent les fleurs[3]. La sélection directionnelle a précisément conduit les plantes à développer davantage de variations colorées, notamment parmi les ultraviolets, attirant davantage de pollinisateurs[3]. Certains pollinisateurs pourraient utiliser le tétrachromatisme pour atteindre des performances plus élevées en butinage, que leurs concurrents trichromates[réf. nécessaire].
Tétrachromatisme éventuel des humains
Le tétrachromatisme des humains a été envisagé de deux façons différentes :
- la rétine des humains — et de leurs cousins proches (certains primates) — dispose généralement de trois types de cônes (ce sont des trichromates), mais, en faible lumière, les bâtonnets pourraient bien contribuer à la perception des couleurs. Ces conditions spéciales laissent donc une petite place au tétrachromatisme dans l'espace colorimétrique humain[8] ;
- chez les humains, deux des gènes spécifiques aux pigments des cônes, nommément OPN1MW et OPN1MW2 (opsines dites classiques de type 2) sont situés au niveau du chromosome X (déterminant sexuel). Parce que les femmes possèdent deux chromosomes X dans leurs cellules, il a été suggéré que certaines d'entre elles pouvaient porter des variations pigmentaires différentes susceptibles d'en faire des tétrachromates nées. Il faudrait néanmoins qu'elles possèdent quatre types de cônes, équipés de canaux distincts pour mener l'information jusqu'au cerveau, et qui répondent réellement à des longueurs d'onde différentes[9]. Une étude suggère qu'environ 2 à 3 % des femmes pourraient posséder un type de cône dont le spectre d’absorption se situerait entre celui du cône vert et du cône rouge habituel. Théoriquement, elles pourraient mieux différencier les couleurs[10]. Une autre étude suggère que la moitié des femmes et 8 % des hommes pourraient bien être pourvus de quatre photo-pigments[9].
Ces hypothèses n'ont pas encore suffisamment fait l'objet de recherches pour qu'on puisse les considérer comme vérifiées. On aurait néanmoins identifié deux tétrachromates possibles en Angleterre[11],[10], mais ces dispositions naturelles n'ont pas encore fait l'objet d'une vérification complète.
Les variations génétiques concernant les photopigments sont courantes au sein de la plupart des populations humaines, mais le tétrachromatisme le plus à même de s'exprimer et qui serait le plus prononcé se manifesterait probablement chez les femmes porteuse d'une anomalie des pigments vert et rouge (protanopie ou deutéranopie). Cela résulterait de l'inactivation X des allèles hétérozygotes spécifiques aux gènes des pigments. C'est en tout cas le mécanisme qui fait s'exprimer le trichromatisme chez certains singes de sexe féminin du Nouveau Monde.
Chez les humains, le traitement de l'information visuelle commence dès la rétine, qui contient des neurones. Nous ne savons pas comment ces nerfs traiteraient un flux d'information supplémentaire sur la couleur, notamment s'ils l'achemineraient séparément jusqu'au cerveau ou pas. Il n'est pas certain que le nerf optique ait des ressources suffisantes pour un nouveau canal. De même, devant un canal supplémentaire, la réaction des différentes zones du cerveau concernées est inconnue.
Des expérimentations animales ont été menées sur des souris qui normalement n'ont que deux types de photopigments. Or les manipulations génétiques ont exprimé un troisième type de photopigment chez ces souris qui auraient fait montre d'une capacité de différenciation des couleurs comparativement plus élevée[12]. Toutefois, l'objet de la publication qui portait sur la plasticité du nerf optique a fait l'objet d'une controverse[13].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Tetrachromacy » (voir la liste des auteurs).
- Les données du graphique trouvent leur origine dans les courbes d'absorbance non corrigées publiées par : Hart NS, Partridge JC, Bennett ATD and Cuthill IC (2000) "Visual pigments, cone oil droplets and ocular media in four species of estrildid finch." Journal of Comparative Physiology A186 (7-8): 681-694.
- (en) Susan E. Wilkie, « The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus) », Biochemical Journal, vol. 330, no Pt 1, , p. 541–47 (PMID 9461554, PMCID 1219171)
- (en) W. Backhaus, R. Kliegl et J.S. Werner, Color vision : perspective from different disciplines, , p. 163-182
- (en) J. Robinson, E.A. Schmitt, F.I. Harosi, R.J. Reece et J.E. Dowling, « Zebrafish ultraviolet visual pigment: absorption spectrum, sequence, and localization », Proc. Natl. Acad. Sci. U.S.A., vol. 90, no 13, , p. 6009–6012 (PMID 8327475, PMCID 46856, DOI 10.1073/pnas.90.13.6009)
- (en) Bennett, A.T.D., Cuthill, I.C., Partridge, J.C., Maier, E.J., « Ultraviolet vision and mate choice in zebra finches », Nature, vol. 380, no 6573, , p. 433–435 (DOI 10.1038/380433a0)
- (en) Bennett, A.T.D., Cuthill, I.C, « Avian color vision and coloration: multidisciplinary », American Naturalist, vol. 169, , p. 1–6
- (en) Markham, K.R., Bloor, S.J., R. Nicholson, R. Rivera, M. Shemluck, P.G. Kevan, C. Michener, « Black flower coloration in wild Lisianthius nigrescens: its chemistry and ecological consequences », Zeitschrift für Naturforschung C, vol. 59c, , p. 625–630
- (en) Hansjochem Autrum et Richard Jung, Integrative Functions and Comparative Data, vol. 7 (3), Springer-Verlag, (ISBN 978-0-387-05769-9, lire en ligne), p. 226
- (en) Jameson, K. A., Highnote, S. M., & Wasserman, L. M., « Richer color experience in observers with multiple photopigment opsin genes », Psychonomic Bulletin and Review, vol. 8, no 2, , p. 244–261 (PMID 11495112, DOI 10.3758/BF03196159, lire en ligne [PDF])
- (en) Mark Roth, « Some women may see 100,000,000 colors, thanks to their genes », Pittsburgh Post-Gazette,
- (en) « You won't believe your eyes: The mysteries of sight revealed », The Independent, (lire en ligne)
- (en) Jacobs et al., GA Williams, H Cahill et J Nathans, « Emergence of Novel Color Vision in Mice Engineered to Express a Human Cone Photopigment », Science, vol. 315, no 5819, , p. 1723–1725 (PMID 17379811, DOI 10.1126/science.1138838, lire en ligne)
- (en) W. Makous, « Comment on "Emergence of Novel Color Vision in Mice Engineered to Express a Human Cone Photopigment" », Science, vol. 318, no 5848, , p. 196 (PMID 17932271, DOI 10.1126/science.1146084, lire en ligne)
Articles connexes
 Portail de la biologie
Portail de la biologie  Portail des couleurs
Portail des couleurs