Vision des couleurs
La vision des couleurs est la capacité d'un animal ou d'une machine de distinguer des objets qui ne diffèrent que par le spectre de la lumière qu'ils réfléchissent.
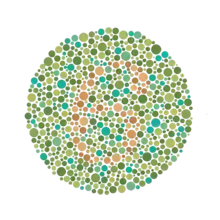
La notion de couleur dépend de la vision humaine, qui, sauf cas de daltonisme, connaît le rayonnement électromagnétique entre 400 et 700 nanomètres par un processus cognitif basé sur trois types de photorécepteurs dans la rétine. L'humain considère la couleur comme un attribut de l'objet, interprétant le rayonnement qu'il reçoit de cet objet en relation avec ce qui l'entoure. Cette caractéristique implique que le système visuel soit capable d'évaluer la lumière qu'il reçoit de l'objet en référence à celle qu'il reçoit de son environnement.
Les animaux sont également dotés de photorécepteurs utilisant plusieurs catégories de protéines pigmentaires, les opsines, qui détectent les changements d'intensité et de fréquence de la lumière. Selon le nombre de ces photorécepteurs, les biologistes distinguent les animaux monochromates, dichromates, trichromates, tétrachromates.
Le concept de couleur est souvent généralisé dans des applications scientifiques et techniques, qui ne visent pas toujours à produire des images en couleurs à l'usage des humains, mais, plus généralement, à distinguer des corps qui diffèrent principalement par la répartition spectrale de la lumière qu'ils renvoient. Pour distinguer des éléments chimiques par leur spectre d'émission ou d'absorption, il faut plusieurs dizaines de bandes. Ces bandes ne coïncident pas nécessairement avec celles de la vision humaine.
Vision animale
Parler de vision des couleurs pour les animaux implique des épreuves spécifiques, puisque ceux-ci ne peuvent répondre verbalement sur ce qu'ils voient. On doit leur appliquer la définition opérationnelle de la couleur, et déterminer s'ils sont capables de prendre une décision basée sur la caractéristique de réflectance spectrale des objets.
Les investigations portent en premier lieu sur les organes de la vision, dans lesquels il faut trouver des capteurs de sensibilité spectrale différente. Mais cette condition ne suffit pas à établir la vision en couleurs, qui est l'aptitude à distinguer des objets colorés. Certaines espèces ont une réaction invariable à la présence de certaines répartitions spectrales, on ne peut guère dire qu'elles voient. On peut vérifier la vision en couleurs pour les espèces capables d'apprentissage en faisant varier la couleur du stimulus qui déclenche une punition ou une récompense, et en s'assurant que l'attitude de l'animal se conforme à une variation de teinte, et non de luminosité. Cela ne prouve pas que les espèces qu'on ne peut soumettre à cette épreuve ne voient pas en couleurs, mais cela permet de l'affirmer, et de le préciser, pour celles qui le peuvent[1].
Les investigations ont montré que nombre d'espèces voient en couleurs avec les trois phénomènes qui caractérisent la vision en couleur humaine[2] :
- la reconnaissance de stimulus aussi bien achromatiques (pure variation de luminosité) que chromatiques (dominance d'une région spectrale) ;
- le contraste de couleur simultané et successif ;
- la constance de couleur dans des conditions d'éclairage variées.
Évolution de la vision des couleurs
 |
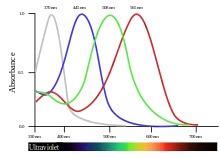 | |
Courbes de sensibilité spectrale des photorécepteurs chez les animaux dichromates et tétrachromates. | ||

L’évolution de la vision des couleurs chez les animaux est extrêmement complexe car elle reflète leur adaptation à des modes de vie (communications par des couleurs utilisées comme moyen de protection, mimétisme, pour les parades nuptiales, le fourrageage, etc.) et des environnements très divers.
La discrimination des spectres lumineux, qui peut présenter divers avantages, est possible en l'absence d'organe visuel. Le nématode Caenorhabditis elegans se nourrit de bactéries, et se détourne de celles qui sécrètent une toxine. Ce ver fuit les régions où une lumière bleuâtre réplique la couleur de la toxine que, sans lumière ou dans une lumière ambrée, il ne détecte que chimiquement et à une teneur plus élevée. Tous les variants de l'espèce n'ont pas la même discrimination, mais aucun ne possède de gènes codant une opsine, protéine fondamentale de la vision. La comparaison des gènes de lignées sensibles ou insensibles à la couleur a permis d'identifier deux gènes corrélés avec cette sensibilité spectrale[4],[5].
La vision des poissons est adaptée à leur environnement lumineux : les espèces des profondeurs ont une vision monochrome grâce à leurs cônes sensibles au bleu, les espèces vivant dans les eaux peu profondes peuvent être tétrachromates, c'est-à-dire avec quatre types de cônes[6].
La majorité des poissons téléostéens, des oiseaux, reptiles et thérapsides (« reptiles mammaliens ») jouissent aussi d’une vision probablement tétrachromate, un des types de cône étant sensible aux ultraviolets, utiles pour différentes fonctions et actif dans la sélection sexuelle[7]. Les premiers mammifères sont dichromates. Leurs deux sortes de cônes sont sensibles au bleu et vert, la perte de deux pigments correspondant à leur mœurs de chasseurs nocturnes insectivores au Trias[8]. La vision devient également monochromatique (cône sensibles au bleu) ou dichromatique (cônes sensibles au bleu et vert) par perte d'opsines chez les mammifères aquatiques[6],[9]. La plupart des mammifères actuels (bovidés, équidés, canidés, félidés, singes du nouveau monde) sont aujourd'hui considérés comme des dichromates, héritage des ancêtres nocturnes[10].
Chez les primates, la perte de plusieurs gènes de récepteurs olfactifs associés à des opsines coïncide avec l'acquisition de gènes de photorécepteurs impliqués dans la vision trichromatique[11]. Chez ces animaux, la tendance évolutive vers la disparition du museau et la régression du prognathisme facial est liée à la réduction de l'appareil olfactif au profit du système tactile et visuel (adaptation à la vie arboricole diurne grâce à des mains et pieds préhensiles, développement d'aires visuelles assurant une vision binoculaire stéréoscopique et une meilleure coordination dans l'obscurité de la forêt[alpha 1]), et un trichromatisme lié à l'évolution d'un régime alimentaire exclusivement insectivore à un régime de plus en plus omnivore (fruits colorés, jeunes pousses, noix, insectes présentant une taille et une consistance très variées, etc.)[13]. Cette vision trichromatique (vision rouge-vert supplémentaire) serait un avantage sélectif permettant de distinguer tout ce qui n'a pas la même couleur que celle d'un feuillage mature (fruits et jeunes pousses de la végétation)[14]. Une autre hypothèse propose d'envisager que la détection du rougissement de l'épiderme (et des humeurs qui l'accompagnent) pourrait avoir influencé le développement du trichromatisme de la vision chez les primates. La psychologie des couleurs attribue à la couleur rouge des effets sur le comportement des primates, humains compris[15].
Chez les primates haplorrhiniens (Singes et humains), le gène codant le pigment visuel absorbant les longueurs d'onde moyennes donne naissance par duplication à deux gènes. Cette mutation génétique leur fait gagner une couleur mais aussi une acuité visuelle bien supérieure à celles des autres mammifères[16].
| Désignation | nombre de types de cônes | Espèces concernées |
|---|---|---|
| Monochromatisme | 1 | Mammifère marin, douroucouli, lion de mer australien, primate achromate. |
| Dichromatisme | 2 | La plupart des mammifères en dehors des primates, les primates daltoniens. |
| Trichromatisme | 3 | La plupart des primates, tels que les grands singes (dont les êtres humains), les marsupiaux, certains insectes (tels que les abeilles). |
| Tetrachromatisme | 4 | La plupart des reptiles, les amphibiens, les oiseaux et les insectes, rarement les humains. |
| Pentachromatisme | 5 | Certaines espèces de papillons et autres insectes ; certains oiseaux comme les pigeons. |
Le nombre de ces types donne un indice du nombre de dimensions de l'espace de couleurs de l'animal ; mais dans certaines espèces, comme le ouistiti, un tiers le la population est trichromate, les autres, dichromates ; dans d'autres, il y a plusieurs variantes de pigments détecteurs dans la rétine[18]. La vision d'oiseaux diurnes comme les pigeons et les canards est encore plus complexe. Leur rétine a au moins quatre types de cônes, et des gouttelettes d'huile colorées ou non par des carotènes s'interposent au dessus de ceux-ci, comme des filtres. Le pigeon a deux régions fovéales distinctes, capables de percevoir la couleur avec une sensibilité et une discrimination différentes[19]. Le système visuel de ces oiseaux traite bien plus de données brutes que l'humain, avec ses trois types sans filtre, et il est préférable d'éviter de se l'expliquer en termes de couleurs, telles que nous les comprenons[20].
Certaines espèces de papillons comme Graphium sarpedon ont des photorécepteurs avec au moins 15 sensibilités spectrales différentes, divisés en 6 classes de l'ultraviolet (300 nm) au rouge, répartis différemment dans les facettes des yeux de ces insectes. Des pigments fluorescents ou non et ces effet de filtrage par réflexion multiples peuvent modifier la sensibilité spectrale des opsines qui permettent la conversion de la lumière en influx nerveux. Cette diversités de récepteurs n'implique pas que leur vision des couleurs soit décapentochromatique. On ne pourrait arriver à cette conclusion que par des expériences comportementales. L'étude d'autre papillons montre que certains photorécepteurs n'interviennent que dans la détection de signaux particuliers, ou dans la vision en basse lumière, ou la détection de la couleur d'objet en déplacement rapide, tandis que la vision est tétrachromatique[21].
Vision humaine

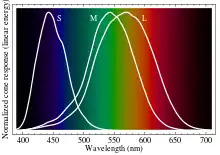
L'humain a dans la rétine trois types de cellules sensibles à la lumière. Ces cellules appelées cônes contiennent des pigments que l'exposition à une partie spécifique du spectre lumineux décompose, créant un influx nerveux avant leur reconstitution par l'organisme. La vision diurne humaine est donc trichromatique. La vision nocturne utilise un type unique de récepteur plus sensible que les cônes appelé bâtonnet, et ne permet pas de distinguer des couleurs.
Les trois types de cônes ne correspondent pas à des couleurs particulières. Ils sont conventionnellement désignés selon la longueur d'onde du maximum de leur sensibilité spectrale : court (S, pour short), moyen (M), et long (L). La présence de récepteurs dont la sensibilité spectrale recouvre les mêmes régions du spectre, mais dont les maxima ne coïncident pas, ne suffit pas à constituer la vision des couleurs. Il faut un réseau neuronal capable de comparer les influx nerveux qu'ils produisent[22]. Un processus complexe produit la perception de la couleur. La concordance entre les réponses des cônes M et L produit la sensation de clarté ; leur différence établit un axe rouge-vert, tandis que la différence entre l'ensemble M et L avec les cônes S établit un axe jaune-bleu[23],[24].
Ces deux axes différentiels constituent la différence entre une vision achromatique et la vision des couleurs. Les oppositions qu'ils créent n'appartiennent pas au domaine physique ; dans le cadre de la physique, il n'y a aucune raison d'opposer les rayonnements où dominent une longueur d'onde de 470 nm à ceux où dominent une longueur d'onde de 573 nm[alpha 3]. La notion humaine de couleur inclut ces oppositions[26].
Variations de la vision humaine
Certains humains sont dichromates, avec seulement deux des trois types de cônes L, M, S. Cette condition est généralement considérée comme un handicap, puisqu'elle gêne la réalisation de certaines tâches ; mais on lui a trouvé, quelquefois, des avantages, dans la mesure où la vision exploite mieux la plus faible variété des signaux issus de la rétine.
Des études indiquent que les pigments des cônes varient dans l'espèce humaine. La sensibilité d'un des pigments, généralement celui d'un des cônes L ou M, est décalée sur une position intermédiaire entre les deux. Ces personnes se détectent surtout par le fait que dans la tâche consistant à mélanger deux lumières, une rouge et une verte, pour égaliser la teinte avec un jaune, elles donnent des proportions différentes de celles de la population générale. Une étude plus complète des résultats de cette épreuve psychophysique montre une que la population se répartit, de ce point de vue, en deux groupes ayant une sensibilité différente au rouge. Ces faits ne se détectent pas dans les tâches quotidiennes, mais ont une importance considérable pour la compréhension de la vision des couleurs[27].
Constance de couleur
La vision des couleurs est un processus d'interaction avec l'environnement, et non une mesure du rayonnement visible. L'humain associe une couleur stable aux objets, compensant les différences d'éclairement par comparaison avec l'entourage. L'échiquier d'Adelson met ce processus en évidence. L'adaptation visuelle chromatique compense les variations d'ensemble, la loi du contraste simultané des couleurs résulte de la comparaison locale[28]. Il en résulte une certaine constance de la couleur d'un objet dans diverses circonstances. La colorimétrie échappe à cette complexité en comparant les couleurs une à une et dans un environnement uniforme[29].
Selon une approche écologique de la perception visuelle, ces particularités, qui font, dans une certaine mesure, diverger la vision des couleurs de l'analyse spectrale du rayonnement, ont triomphé parce qu'elles favorisent ceux qui en sont dotés dans leur interaction avec leur environnement, tant animé qu'inanimé. La stabilité de la perception de couleur, dans des environnements lumineux très variés, leur permet de repérer les objets intéressants dans tous les cas [30].
Vision en couleurs des machines
Pour qu'on puisse parler de vision en couleurs pour une machine, il faut que celle-ci puisse fonder une décision sur une différence de réponse entre des capteurs de sensibilité spectrale différente.
Pour évaluer la température de couleur d'un objet irradiant par l'effet de sa chaleur, on utilise le plus souvent deux capteurs, dont au moins un est sensible au rayonnement infrarouge, invisible pour l'humain.
La machine recherche la température du corps noir qui donnerait la même différence relative entre les deux plages. Cette évaluation de température sans contact peut ensuite déclencher une action.
Une machine peut évaluer sur le même modèle, par la couleur de leur peau, le degré de maturité d'un agrume ou d'une banane, et lancer une récolte. Comme l'évolution de la couleur suit un parcours spectral bien défini du vert au jaune ou à l'orange, il suffit pour cela de deux capteurs. La machine doit repérer le fruit par sa forme, ce qui peut se faire en noir et blanc, situer sa couleur, indépendamment de l'éclairage ou en l'éclairant avec une lumière connue, et déterminer sa place sur l'échelle de maturité.
Lorsque le système doit identifier des spectres dont l'évolution n'est pas, comme dans les deux cas précédents, régie par une loi, il faut que les sensibilités spectrales des capteurs se recouvrent. La luminosité s'obtient par l'addition pondérée des résultats des deux types de capteurs, la « couleur » par leur différence. La meilleure discrimination des « couleurs » se produit alors dans la région où la sensibilité d'un des types de capteurs croît rapidement, alors que l'autre décroît rapidement. Le nombre de types de capteurs donne autant de variables indépendantes pour définir une « couleur », ce qu'on exprime généralement par le nombre de dimensions de l'espace de couleur[31].
Le problème de la vision des couleurs pour une machine est identique à celui qui se pose aux animaux. Le spectre de la lumière qui arrive de l'objet est le produit de la distribution spectrale de l'éclairant par celle de la réflexion de l'objet. Il faut évaluer celui de l'illuminant, à partir de l'image de l'environnement, pour connaître celui lié à l'objet, qui est la grandeur intéressante[32].
Annexes
Bibliographie
- (en) Evan Thompson, Colour Vision : A Study in Cognitive Science and Philosophy of Science, Routledge, (présentation en ligne)
- Claude Romano, De la couleur : cours, Paris, Éditions de la Transparence, coll. « Philosophie », .
- (en) F. Crescitelli, The Visual System in Vertebrates, Springer Science & Business Media, (lire en ligne)
- Yves Le Men, La vision dans le monde animal, AlterPublishing, (lire en ligne)
Articles connexes
Références
- La migration des orbites vers l'avant assure en outre une meilleure vision binoculaire stéréoscopique. Il existe au sein des primates une grande variabilité sur le degré de convergence orbitale et la taille du champ binoculaire mais il semble que cette convergence et la vision binoculaire correspondent aux ancêtres des primates. Ces derniers étaient des insectivores avec cette vision adaptée pour mieux discerner le camouflage de leurs proies habitant un environnement nocturne[12].
- La crevette-mante (Stomatopoda) a un système visuel unique dans le règne animal, avec de 12 à 16 récepteurs de sensibilité différente, une vision stéréoscopique dans chacun de leurs deux yeux, et certaines espèces distinguent la polarisation de la lumière[17].
- Avec une apparence respectivement bleue et jaune, couleurs complémentaires pour l'illuminant D relevée dans le diagramme de chromaticité CIE xy[25].
- Thompson 1995, p. 141-142.
- Thompson 1995, p. 145.
- (en) Rui Borges, Warren E. Johnson, Stephen J. O’Brien, Cidália Gomes, Christopher P. Heesy & Agostinho Antunes, « Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments », BMC Genomics, vol. 19, no 1, , p. 21 (DOI 10.1017/S0952523805226159).
- (en) Lauren A. Neal1 et Leslie B. Vosshall, « Eyeless worms detect color », Science, vol. 371, no 6533, , p. 995 (DOI 10.1126/science.abg5201).
- (en) D. Dipon Ghosh, Dongyeop Lee, Xin Jin, H. Robert Horvitz et Michael N. Nitabach, « C. elegans discriminates colors to guide foraging », Science, vol. 371, no 6533, , p. 1059-1063 (DOI 10.1126/science.abd3010).
- (en) L. Peichl, G. Behrmann et R. H. Kröger, « For whales and seals the ocean is not blue: a visual pigment loss in marine mammals », Eur. J. Neurosci., vol. 13, no 8, , p. 1520–1528 (DOI 10.1046/j.0953-816x.2001.01533.x)
- (en) Justin Marshall et Kentaro Arikawa, « Unconventional colour vision », Current Biology, vol. 24, no 24, , p. 1150-1154 (DOI 10.1016/j.cub.2014.10.025).
- (en) G. H. Jacobs, M. Neitz et J. Neitz, « Mutations in S-cone pigment genes and the absence of colour vision in two species of nocturnal primates », Proc. R. Soc. Lond. B., vol. 263, no 1371, , p. 705–710 (DOI 10.1098/rspb.1996.0105).
- (en) L. A. Newman et P. R. Robinson, « Cone visual pigments of aquatic mammals », Vis. Neurosci., vol. 22, no 6, , p. 873-879 (DOI 10.1017/S0952523805226159).
- (en) Michael H. Rowe, « Trichromatic color vision in primates », News in Physiological Sciences, no 17 (3), , p. 93-98 (lire en ligne).
- (en) Yoav Gilad, Victor Wiebe, Molly Przeworski, Doron Lancet, Svante Pääbo, « Loss of Olfactory Receptor Genes Coincides with the Acquisition of Full Trichromatic Vision in Primates », PLoS Biol, vol. 2, , p. 120-125 (DOI 10.1371/journal.pbio.0020005).
- (en) Callum F. Ross, « Allometric and functional influences on primate orbit orientation and the origins of the anthropoidea », Journal of Human Evolution, vol. 29, no 3, , p. 201–227 (DOI 10.1006/jhev.1995.1057).
- Robert Boyd et Joan Silk, L'aventure humaine. Des molécules à la culture, De Boeck Supérieur, , p. 264.
- (en) Sharpe et al., « Advantages and disadvantages of human dichromacy », Journal of Vision, no 6, , p. 213-223 « Copie archivée » (version du 10 octobre 2014 sur l'Internet Archive).
- Diana Widermann, Robert A. Barton, and Russel A. Hill. Evolutionary perspectives on sport and competition. In DOI:10.1093/acprof:oso/9780199586073.001.0001
- (en) Kanwaljit S. Dulai, Miranda von Dornum, John D. Mollon & David M. Hunt, « Duplication in New World and Old World Primates », Genome Res., vol. 9, , p. 629-638 (DOI 10.1101/gr.9.7.629).
- (en) « Secrets of the stomatopod » (consulté le ).
- Thompson 1995, p. 163sq.
- Thompson 1995, p. 149.
- Thompson 1995, p. 151.
- (en) Chen, Pei-Ju, Awata, Hiroko, Atsuko, Matsushita, Yang, En-Cheng et Arikawa, Kentaro, « Extreme spectral richness of the eye of the Common Bluebottle, Graphium sarpedon », Frontiers in Physiology, (ISSN 1664-042X, lire en ligne)
- Thompson 1995, p. 56.
- Robert Sève, « 1. Vision des couleurs : Aspects physiques et perceptifs », dans Science de la couleur, Marseille, Chalagam, , p. 27
- Thompson 1995, p. 38sq.
- Kodak-Pathé, Filtres Kodak : pour usages scientifiques et techniques, , p. 18.
- Thompson 1995, p. 186.
- Thompson 1995, p. 166.
- Thompson 1995, p. 57.
- Thompson 1995, p. 40.
- Romano 2010, p. 72sq.
- Thompson 1995, p. 146-148.
- Thompson 1995, p. 45.
 Portail des couleurs
Portail des couleurs  Portail de la philosophie
Portail de la philosophie  Portail de la physiologie
Portail de la physiologie  Portail de l’œil et de la vue
Portail de l’œil et de la vue