Genetics, Histocompatibility Antigen
- Article Author:
- Sarah Kamal
- Article Author:
- Connor Kerndt
- Article Editor:
- Sarah Lappin
- Updated:
- 8/29/2020 9:42:16 PM
- For CME on this topic:
- Genetics, Histocompatibility Antigen CME
- PubMed Link:
- Genetics, Histocompatibility Antigen
Introduction
Genetically encoded molecules, known as major histocompatibility complex (MHC) molecules or human leukocyte antigens (HLA), determine an individual’s ability to produce an immune response to different antigens. These MHC molecules are responsible for determining if a molecule is histocompatible with the individual and for initiating an immune response if it is deemed foreign.[1] They are found in all nucleated cells and primarily function in antigen presentation to T cells. For an antigen presenting cell (APC) and a T cell to interact and initiate an immune response, the T cell must bind to the APC as well as the antigen bound to an MHC molecule. A major histocompatibility complex is a set of genes that code these MHC molecules/HLA antigens. Multiple genes code for different parts of the molecules and several possible combinations of gene products exist. As a result, the chance of two individuals expressing identical MHC molecules is tremendously low. Clinically, MHC molecules are significant as they are the principle mediators involved in transplant rejection.[2]
Development
The human major histocompatibility complex or human leukocyte antigen system exhibits polymorphism. It is comprised of over 200 genes, encoding for several different molecules involved in immune defense.[1] All human genes encoding MHC molecules are present on the short arm of chromosome 6. Two principal types of MHC molecules involved in antigen presentation and the associated immune response are class I and class II MHC molecules. Three unique loci contain genes that code for Class I MHC molecules, defined as A, B, and C. Class II MHC genes are at loci called DR, DQ, and DP. Multiple alleles are possible at each of these loci, and an individual can be either homozygous or heterozygous for each gene, thus creating the polymorphism (Figure 1). Genes of the MHC system are codominant, and therefore all inherited alleles code for specific MHC molecules to express on a cell surface. As a consequence, every human inherits a set of genes for A, B, C, DR, DP, and DQ. Remarkable variability exists ensuring that a person’s MHC type is unique.[3]
Biochemical
The following discusses the structures of Class I and II MHC molecules (Figure 2):
Class I MHC molecules are glycoprotein dimers, composed of two polypeptide chains, including a heavy chain and a light chain. The lighter chain is call beta2–microglobulin and is encoded by a single non-polymorphic gene located on chromosome 15. The heavy chain inserts into the cell membrane by a hydrophobic transmembrane segment. Beta2–macroglobulin functions in folding of the heavy chain, but does not penetrate the cell membrane. The heavy chain involves three alpha-helix domains, named alpha-1, alpha-2, and alpha-3. Alpha-1 and alpha-2 form the peptide-binding site that is used in antigen recognition and exhibits the most structural variation.[4]
Class II MHC molecules are also composed of two polypeptide chains; however in contrast to Class I molecules both chains are encoded by polymorphic MHC complex genes on chromosome 6. Genes at the DR locus are most highly expressed and the most polymorphic. Both the heavy and light chains insert into the cell membrane. The heavy chain has two domains named alpha-1 and alpha-2, and the light chain has domains beta-1 and beta-2. Alpha-1 and beta-1 form the peptide-binding cleft.[5]
Function
When antigens are inside of cells, they are cleaved into peptides, and those peptides bind to MHC class I or class II molecules. MHC molecules then carry the antigen to the plasma membrane and form a complex that can be recognized by T cells. Each MHC molecule can bind to various peptides and present those antigens to T cells. Class I MHC molecules primarily present peptide products that are synthesized inside of the cell to cytotoxic (CD8) T cells.[6] In contrast, class II MHC molecules primarily present exogenous proteins that were taken up and degraded by the cell to helper (CD4) T cells.[7] Therefore, class I molecules are responsible for presenting viral, tumor, or parasitic antigens and class II molecules are responsible for initiating the immune response in bacterial infections.
Mechanism
Major histocompatibility complex molecules are initially synthesized in and anchored to the rough endoplasmic reticulum (ER). After MHC class I chains are synthesized, they are first bound to calnexin, a membrane-bound molecule in the rough ER that stabilizes the chain until it can bind to beta2-microglobulin. After synthesis, chaperone proteins calreticulin, tapasin, and ERp57 additionally stabilize the molecule until an antigenic peptide binds. Proteins synthesized in the cytoplasm are then cleaved into peptides about 8 to 10 amino acids long and transported to the ER by ATP-dependent transporter proteins TAP1 or TAP2, where they can bind to MHC class I molecules. Any antigen that produces intracellular proteins, whether viral, bacterial, or tumor in etiology, could bind to MHC. Also, defective proteins that did not fold correctly and are digested by proteasomes, also known as defective ribosomal products (DRiPs) could bind to MHC. Tapasin is a molecule involved in stabilizing the transfer of peptides from TAP transporters to MHC molecules. Once the MHC I-peptide complex forms, it is transported to the cell surface, where it could potentially bind to a cytotoxic T cell and induce a response. In most healthy cells, if the complex contains a self-antigen, no immune response will be triggered. However, if the CD8 T cell identifies a non-self antigen, then it will trigger cell lysis.[1]
The mechanism of class MHC molecule function is similar; however, class II molecules first get transported to an endosome before binding to a peptide. In the endoplasmic reticulum, they are bound to an invariant chain that prevents them from binding to any peptides inside the ER. After binding to the invariant chain, the MHC-invariant chain complex is transported to an endosome. Most of the invariant chain is cleaved by a protease, and a small molecule named class II invariant chain peptide (CLIP) remains at the peptide-binding cleft. Meanwhile, the antigen-presenting cell, such as a dendritic cell, will phagocytize an exogenous peptide antigen, creating another endosome. The endosome containing the digested peptide will then fuse with the MHC-CLIP endosome. After fusion, preferential binding of the exogenous peptide displaces CLIP. Additionally, HLA-DM molecules help remove CLIP. After the MHC-peptide complex forms, it is transported to the cell surface. It can then bind to the T-cell receptor on a CD4 T cell, which will eventually result in antibody formation by B cells. Note that binding of the MHC-peptide complex to the T cell receptor is one of two interactions required for a successful immune response. The other is the binding of the B7 receptor on the antigen-presenting cell to the CD28 receptor on the CD4 T cell.[2]
(Figures 3 and 4)
Clinical Significance
Both major histocompatibility complex class I and class II molecules are responsible for the acute immune response that progresses to graft rejection. Because of the significant genetic variability in HLA antigens between individuals, donor organs can contain highly immunogenic antigens. It is the donor’s HLA molecules themselves that get recognized as non-self HLA, which leads to the generation of anti-HLA antibodies and subsequent graft damage.[8] In fact, antibody-mediated rejection is the most common cause of kidney or heart transplant loss. These antibodies can cause necrotic cell death in endothelial cells. The mechanism of cell damage is via both complement-dependent and complement-independent pathways. The complement-dependent antibody response involves the classical complement cascade while the complement-independent response involves intracellular signal transduction cascades.[9]
Donor organs are often preferentially taken from siblings or close family members and tested for HLA compatibility to minimize this risk. Patients typically undergo crossmatching to identify any preexisting HLA antibodies. However, post-transplant monitoring for anti-HLA antibodies is still required as they can develop even years after the transplant.[10]
(Click Image to Enlarge)
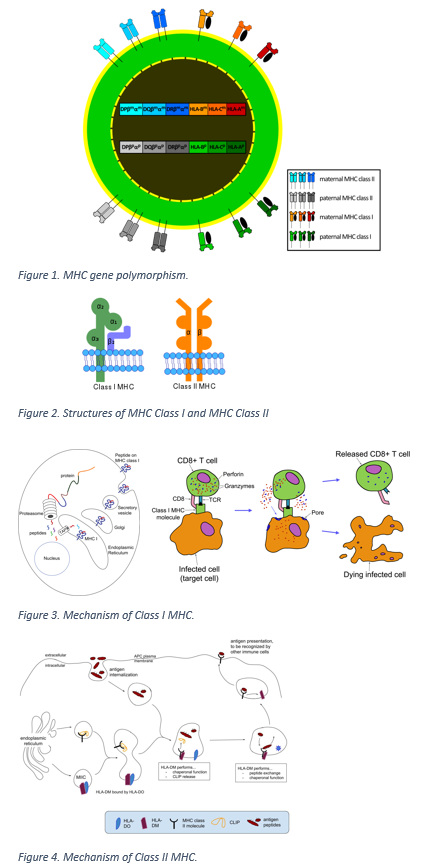
HLA/MHC gene polymorphism, structure, and mechanism
Contributed from the Wikimedia Gallery (CC BY-2.5 https://creativecommons.org/licenses/by/2.5/)