B-cell receptor

The B cell receptor (BCR) is a transmembrane protein on the surface of a B cell. A B cell receptor is composed of a membrane-bound immunoglobulin molecule and a signal transduction moiety. The former forms a type 1 transmembrane receptor protein, and is typically located on the outer surface of these lymphocyte cells.[1] Through biochemical signaling and by physically acquiring antigens from the immune synapses, the BCR controls the activation of the B cell.[2] B cells are able to gather and grab antigens by engaging biochemical modules for receptor clustering, cell spreading, generation of pulling forces, and receptor transport, which eventually culminates in endocytosis and antigen presentation.[1] B cells' mechanical activity adheres to a pattern of negative and positive feedbacks that regulate the quantity of removed antigen by manipulating the dynamic of BCR–antigen bonds directly.[3] Particularly, grouping and spreading increase the relation of antigen with BCR, thereby proving sensitivity and amplification.[4] On the other hand, pulling forces delinks the antigen from the BCR, thus testing the quality of antigen binding.
The receptor's binding moiety is composed of a membrane-bound antibody that, like all antibodies, has two identical paratopes that are unique and randomly determined. The BCR for an antigen is a significant sensor that is required for B cell activation, survival, and development. A B cell is activated by its first encounter with an antigen (its "cognate antigen") that binds to its receptor, resulting in cell proliferation and differentiation to generate a population of antibody-secreting plasma B cells and memory B cells.[1][4] The B cell receptor (BCR) has two crucial functions upon interaction with the antigen. One function is signal transduction, involving changes in receptor oligomerization.[1] The second function is to mediate internalization for subsequent processing of the antigen and presentation of peptides to helper T cells.
Development and structure of the B cell receptor
The first checkpoint in the development of a B cell is the production of a functional pre-BCR, which is composed of two surrogate light chains and two immunoglobulin heavy chains, which are normally linked to Ig-α (or CD79A) and Ig-β (or CD79B) signaling molecules.[1][5] Each B cell, produced in the bone marrow, is highly specific to an antigen.[1][3] The BCR can be found in a number of identical copies of membrane proteins that are exposed at the cell surface.[1][3][6]
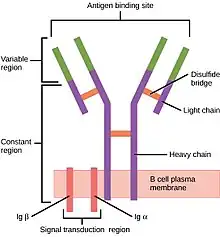
The B cell receptor is composed of two parts:
- A membrane-bound immunoglobulin molecule of one isotype (IgD, IgM, IgA, IgG, or IgE). With the exception of the presence of an integral membrane domain, these are identical to a monomeric version of their secreted forms.
- Signal transduction moiety: a heterodimer called Ig-α/Ig-β (CD79), bound together by disulfide bridges. Each member of the dimer spans the plasma membrane and has a cytoplasmic tail bearing an immunoreceptor tyrosine-based activation motif (ITAM).[6][7]
More analytically, the BCR complex consists of an antigen-binding subunit known as the membrane immunoglobulin (mIg), which is composed of two immunoglobulin light chains (IgLs) and two immunoglobulin heavy chains (IgHs) as well as two heterodimer subunits of Ig-α and Ig-β. In order for mIgM molecules to transport to the surface of the cell, there must be a combination of Ig-α and Ig-β with the mIgM molecules. Pre-B cells that do not generate any Ig molecule normally carry both Ig-α and Ig-β to the cell surface.[1][7]
Heterodimers may exist in the B cells as either an association or combination with another pre B cell-specific proteins or alone, thereby replacing the mIgM molecule. Within the BCR, the part that recognizes antigens is composed of three distinct genetic regions, referred to as V, D, and J.[1][4][8] All these regions are recombined and spliced at the genetic level in a combinatorial process that is exceptional to the immune system. There are a number of genes that encode each of these regions in the genome and can be joined in various ways to generate a wide range of receptor molecules.[1][4][7][8] The production of this variety is crucial since the body may encounter many more antigens than the available genes. Through this process, the body finds a way of producing multiple different combinations of antigen-recognizing receptor molecules. Heavy chain rearrangement of the BCR entails the initial steps in the development of B cell. The short JH (joining) and DH (diversity) regions are recombined first in early pro-B cells in a process that is dependent on the enzymes RAG2 and RAG1.[8][9] After the recombination of the D and J regions, the cell is now referred to as a “late pro-B” cell and the short DJ region can now be recombined with a longer segment of the VH gene.[7][8]
BCRs have distinctive binding sites that rely on the complementarity of the surface of the epitope and the surface of the receptor, which often occurs by non-covalent forces. Mature B cells can only survive in the peripheral circulation for a limited time in when there is no specific antigen. This is because when cells do not meet any antigen within this time, they will go through apoptosis.[6] It is notable that in the peripheral circulation, apoptosis is important in maintaining an optimal circulation of B-lymphocytes.[8][9] In structure, the BCR for antigens are almost identical to secreted antibodies.[1][5] However, there is a distinctive structural dissimilarity in the C-terminal area of the heavy chains, as it consists of a hydrophobic stretch that is short, which spreads across the lipid bilayer of the membrane.
Signaling pathways of the B cell receptor
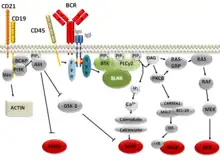
There are several signaling pathways that the B cell receptor can follow through. The physiology of B cells is intimately connected with the function of their B cell receptor. The BCR signaling pathway is initiated when the mIg subunits of the BCR bind a specific antigen. The initial triggering of the B cell receptor is similar for all receptors of the non-catalytic tyrosine-phosphorylated receptor family.[11] The binding event allows phosphorylation of immunoreceptor tyrosine-based activation motifs (ITAMs) in the associated Igα/Igβ heterodimer subunits by the tyrosine kinases of the Src family, including Blk, Lyn, and Fyn. Multiple models have been proposed how BCR-antigen binding induces phosphorylation, including conformational change of the receptor and aggregation of multiple receptors upon antigen binding.[12] Tyrosine kinase Syk binds to and is activated by phosphorylated ITAMs and in turn phosphorylates scaffold protein BLNK on multiple sites. After phosphorylation, downstream signalling molecules are recruited to BLNK, which results in their activation and the transduction of the signal to the interior.[13]
- IKK/NF-κB Transcription Factor Pathway: CD79 and other proteins, microsignalosomes, go to activate PLC-γ after antigen recognition by the BCR and before it goes to associate into the c-SMAC. It then cleaves PIP2 into IP3 and DAG (diacylglycerol). IP3 acts as a second messenger to dramatically increase ionic calcium inside the cytosol (via release from the endoplasmic reticulum or influx from the extracellular environment via ion channels). This leads to eventual activation of PKCβ from the calcium and DAG. PKCβ phosphorylates (either directly or indirectly) the NF-κB signaling complex protein CARMA1 (the complex itself comprising CARMA1, BCL10, and MALT1). These result in recruitment and summoning of the IKK (IkB kinase), TAK1, by several ubiquitylation enzymes also associated with the CARMA1/BCL10/MALT1 complex. MALT1 itself is a caspase-like protein that cleaves A20, an inhibitory protein of NF-κB signaling (which acts by deubiquitylating NF-κB's ubiquitylation substrates, having an inhibitory effect). TAK1 phosphorylates the IKK trimer after it too has been recruited to the signaling complex by its associated ubiquitylation enzymes. IKK then phosphorylates IkB (an inhibitor of and bound to NF-κB), which induces its destruction by marking it for proteolytic degradation, freeing cytosolic NF-κB. NF-κB then migrates to the nucleus to bind to DNA at specific response elements, causing recruitment of transcription molecules and beginning the transcription process.
- Ligand binding to the BCR also leads to the phosphorylation of the protein BCAP. This leads to the binding and activation of several proteins with phosphotyrosine-binding SH2 domains. One of these proteins is PI3K. Activation of PI3K leads to PIP2 phosphorylation, forming PIP3. Proteins with PH (Pleckstrin homology) domains can bind to the newly created PIP3 and become activated. These include proteins of the FoxO family, which stimulate cell cycle progression, and protein kinase D, which enhances glucose metabolism. Another important protein with a PH domain is Bam32. This recruits and activates small GTPases such as Rac1 and Cdc42. These, in turn, are responsible for the cytoskeletal changes associated with BCR activation by modifying actin polymerisation.
The B cell receptor in malignancy
The B cell receptor has been shown to be involved in the pathogenesis of various B cell derived lymphoid cancers. Although it may be possible that stimulation by antigen binding contributes to the proliferation of malignant B cells,[14] increasing evidence implicates antigen-independent self-association of BCRs as a key feature in a growing number of B cell neoplasias[15][16][17][18] B cell receptor signalling is currently a therapeutic target in various lymphoid neoplasms.[19]
See also
References
- 1 2 3 4 5 6 7 8 9 10 11 Owen, J.; Punt, J.; Stranford, S; Jones, P.; Kuby, J. (2013). Kuby Immunology (Seventh ed.). New York: W.H. Freeman and Company. pp. 102–104. ISBN 978-1429219198.
- ↑ Saito, Batista; Saito, Takashi; Facundo, D. (2010). Immunological Synapse (Current Topics in Microbiology and Immunology, 340). Berlin: Heidelberg: Springer Berlin Heidelberg. ISBN 978-3642038570.
- 1 2 3 Merlo, Lauren M. F.; Mandik-Nayak, Laura (2013). Cancer Immunotherapy: Chapter 3-Adaptive Immunity: B Cells and Antibodies. London: Academic Press; 2 edition. pp. 25–40. ISBN 978-0-12-394296-8.
- 1 2 3 4 Dal Porto, JM; Gauld, SB (2014). Merrell KT, Mills D, Pugh-Bernard AE. "B cell antigen receptor signaling 101". Mol Immunol. 41: 599–613. doi:10.1016/j.molimm.2004.04.008. PMID 15219998.
- 1 2 Brenzski, Randall J.; Monroe, John G. (2010). "Chapter 2: B cell Receptor". In Sigalov, Alexander B. (ed.). Multichain Immune Recognition Receptor Signaling: From Spatiotemporal Organization to Human Disease (Advances in Experimental Medicine and Biology). Springer; Softcover reprint of hardcover 1st ed. 2008 edition (November 23, 2010). pp. 12–21. ISBN 978-1441918871.
- 1 2 3 Janeway, CA Jr; Travers, P.; Walport, M. (2015). Immunobiology: The Immune System in Health and Disease. New York: Garland Science (5th edition). ISBN 978-0815341017.
- 1 2 3 4 Pier, Gerland B.; Lyczak, Jeffrey B.; Wetzler, Lee M. (2005). Immunology, Infection, and Immunity. Washington D.C.: ASM Press. pp. 234–247. ISBN 978-1555812461.
- 1 2 3 4 5 Hoehn, Kenneth B.; Fowler, Anna; Lunter, Gerton; Pybus, Oliver G. (2016). "The Diversity and Molecular Evolution of B Cell Receptors during Infection". Mol Biol Evol. 33: 1147–57. doi:10.1093/molbev/msw015. PMC 4839220. PMID 26802217.
- 1 2 Alberts, Bruce (2014). Molecular Biology of the Cell. Garland Science; 6th edition. ISBN 978-0815344322.
- ↑ Wan, Leo D.; Clark, Marcus R. (2003). "B cell antigen-receptor signaling in lymphocyte development". Immunology. 110: 411–20. doi:10.1111/j.1365-2567.2003.01756.x. PMC 1783068. PMID 14632637.
- ↑ Dushek O, Goyette J, van der Merwe PA (November 2012). "Non-catalytic tyrosine- phosphorylated receptors". Immunological Reviews. 250 (1): 258–276. doi:10.1111/imr.12008. PMID 23046135.
- ↑ Treanor B (2012). "B cell receptor: from resting state to activated". Immunology. 136 (1): 21–27. doi:10.1111/j.1365-2567.2012.03564.x. PMC 3372753. PMID 22269039.
- ↑ Janeway's immunobiology (8th ed.). Garland Science. 2011. pp. 258–260. ISBN 0815342438.
- ↑ Daneshek W, Schwartz RS (1959). "Leukemia and auto-immunization- some possible relationships" (PDF). Blood. 14: 1151–8. PMID 13813891.
- ↑ Corcos D (1990). "Oncogenic potential of the B cell antigen receptor and its relevance to heavy chain diseases and other B-cell neoplasias: a new model". Research in Immunology. 141 (6): 543–53. doi:10.1016/0923-2494(90)90022-Q. PMID 2284498.
- ↑ Corcos D, Dunda O, Butor C, Cesbron JY, Lorès P, Bucchini D, Jami J (October 1995). "Pre-B-cell development in the absence of lambda 5 in transgenic mice expressing a heavy-chain disease protein". Current Biology. 5 (10): 1140–8. doi:10.1016/S0960-9822(95)00230-2. PMID 8548286.
- ↑ Davis RE, Ngo VN, Lenz G, Tolar P, Young RM, Romesser PB, et al. (January 2010). "Chronic active B-cell-receptor signalling in diffuse large B-cell lymphoma". Nature. 463 (7277): 88–92. Bibcode:2010Natur.463...88D. doi:10.1038/nature08638. PMC 2845535. PMID 20054396.
- ↑ Dühren-von Minden M, Übelhart R, Schneider D, Wossning T, Bach MP, Buchner M, Hofmann D, Surova E, Follo M, Köhler F, Wardemann H, Zirlik K, Veelken H, Jumaa H (September 2012). "Chronic lymphocytic leukaemia is driven by antigen-independent cell-autonomous signalling". Nature. 489 (7415): 309–12. Bibcode:2012Natur.489..309M. doi:10.1038/nature11309. PMID 22885698.
- ↑ Woyach JA, Johnson AJ, Byrd JC (August 2012). "The B-cell receptor signaling pathway as a therapeutic target in CLL". Blood. 120 (6): 1175–84. doi:10.1182/blood-2012-02-362624. PMC 3418714. PMID 22715122.
External links
- B-Cell+Antigen+Receptors at the US National Library of Medicine Medical Subject Headings (MeSH)