Cysteine-rich secretory protein
| Crisp domain (CRD) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| Symbol | Crisp | ||||||||
| Pfam | PF08562 | ||||||||
| Pfam clan | CL0213 | ||||||||
| InterPro | IPR013871 | ||||||||
| |||||||||
Cysteine-rich secretory proteins, often abbreviated as CRISPs, are a group of glycoproteins.[2] They are a subgroup of the CRISP, antigen 5 and Pr-1 (CAP) protein superfamily and also contain a domain related to the ShK toxins.[1] They are substantially implicated in the functioning of the mammalian reproductive system.[3] CRISPs are also found in a variety of snake venoms where they inhibit both smooth muscle contraction and cyclic nucleotide-gated ion channels.[4]
Structure
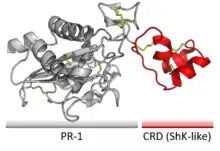
CRISPs contain two domains joined by a hinge region. The larger domain is a CAP-like 'Pathogenesis-related 1' domain (PR-1), followed by the smaller ShK-like 'Cysteine-Rich Domain' (CRD).[1]
CRISPs are glycoproteins, with a number of carbohydrate glycans covalently attached to amino acid side-chains on their surface via glycosylation.[5] The primary structure is also rich in cysteine that form disulfide bonds, particularly in the hinge region and CRD.[1]
Mammalian reproduction
CRISPs are found in the testes and epididymis of mammals, and are also involved in the process of fertilisation.[2] In the spermatogenesis process (development of the spermatozoa in the testis), the CRISP2 protein is incorporated into the acrosome where it is believed to be involved in the adhesion of germ cells with Sertoli cells. CRISP2 also forms part of the sperm tail where it is thought to be involved in regulating flagellar beating. Proteins CRISP1 and CRISP4 are both found in the epididymis where they are also incorporated within the spermatozoa as it matures. Protein CRISP3 is found in seminal fluid, excreted from the prostate although its function is unknown.[3]
During capacitation, the penultimate stage of spermatozoa maturation, the acrosomal sperm head membrane is destabilised to allow greater binding between oocyte and sperm. CRISP1 binds to surface of the sperm leading to a quiescent state of storage prior to capacitation. The mechanism is believed to involve inhibition of ion channel activity, similar to the mechanism of action of the other major function of CRISPs in snake venom.[3] Research also suggests that CRISPs are involved in the oocyte-sperm binding needed for fertilisation.[2] Given the involvement of CRISPs in several stages of human reproduction, it is unsurprising that applications in treatment of infertility and as contraceptives are being actively investigated.[3]
Snake venom
.jpg.webp)
CRISPs are found in the venom of a wide variety of snake species.[4] Examples include ablomin from the Japanese Mamushi snake (Gloydius blomhoffii, formerly Agkistrodon blomhoffi),[6] latisemin from the Erabu sea snake (Laticauda semifasciata),[7] ophanin from the King Cobra (Ophiophagus hannah),[8] piscivorin from the Eastern Cottonmouth (Agkistrodon piscivorus)[8] and triflin from the Habu snake (Trimeresurus flavoviridis)[9] – each of these proteins is named for the snake species in which it was discovered. These venoms are toxic due to their blocking of calcium channels and also because they reduce potassium-induced smooth muscle contraction.[6] Among the four CRISPs isolated from the Monocled Cobra (Naja kaouthia) and the three from the Egyptian Cobra (Naja haje), ion channel activity occurred by blocking of cyclic nucleotide-gated ion channels. One of the N. haje CRISPs was the first example of an acidic CRISP in reptilian venom. The selective ion channel activity of snake CRISPs, coupled with the variety of CRISPs available as the pool of venom proteins appears highly variable between (at least) cobra species, provide a valuable tool for probing the mechanisms of ion channel activity.[10]
References
- 1 2 3 4 Guo M, Teng M, Niu L, Liu Q, Huang Q, Hao Q (April 2005). "Crystal structure of the cysteine-rich secretory protein stecrisp reveals that the cysteine-rich domain has a K+ channel inhibitor-like fold". The Journal of Biological Chemistry. 280 (13): 12405–12. doi:10.1074/jbc.M413566200. PMID 15596436.
- 1 2 3 Cammack R, Attwood TK, Campbell PN, Parish JH, Smith AD, Stirling JL, Vella F, eds. (2006). Oxford Dictionary of Biochemistry and Molecular Biology (2nd ed.). New York: Oxford University Press. p. 150. ISBN 0-19-852917-1. Retrieved October 27, 2010.
- 1 2 3 4 Koppers AJ, Reddy T, O'Bryan MK (January 2011). "The role of cysteine-rich secretory proteins in male fertility". Asian Journal of Andrology. 13 (1): 111–7. doi:10.1038/aja.2010.77. PMC 3739402. PMID 20972450.
- 1 2 Yamazaki Y, Morita T (September 2004). "Structure and function of snake venom cysteine-rich secretory proteins". Toxicon. 44 (3): 227–31. doi:10.1016/j.toxicon.2004.05.023. PMID 15302528.
- ↑ Cammack R, Attwood TK, Campbell PN, Parish JH, Smith AD, Stirling JL, Vella F, eds. (2006). Oxford Dictionary of Biochemistry and Molecular Biology (2nd ed.). New York: Oxford University Press. p. 286. ISBN 0-19-852917-1. Retrieved October 28, 2010.
- 1 2 Yamazaki Y, Koike H, Sugiyama Y, Motoyoshi K, Wada T, Hishinuma S, Mita M, Morita T (June 2002). "Cloning and characterization of novel snake venom proteins that block smooth muscle contraction". European Journal of Biochemistry. 269 (11): 2708–15. doi:10.1046/j.1432-1033.2002.02940.x. PMID 12047379.
- ↑ Yamazaki Y, Morita T (September 2004). "Structure and function of snake venom cysteine-rich secretory proteins". Toxicon. 44 (3): 227–31. doi:10.1016/j.toxicon.2004.05.023. PMID 15302528.
- 1 2 Yamazaki Y, Hyodo F, Morita T (April 2003). "Wide distribution of cysteine-rich secretory proteins in snake venoms: isolation and cloning of novel snake venom cysteine-rich secretory proteins". Archives of Biochemistry and Biophysics. 412 (1): 133–41. doi:10.1016/S0003-9861(03)00028-6. PMID 12646276.
- ↑ Shikamoto Y, Suto K, Yamazaki Y, Morita T, Mizuno H (July 2005). "Crystal structure of a CRISP family Ca2+ -channel blocker derived from snake venom". Journal of Molecular Biology. 350 (4): 735–43. doi:10.1016/j.jmb.2005.05.020. PMID 15953617.
- ↑ Osipov AV, Levashov MY, Tsetlin VI, Utkin YN (March 2005). "Cobra venom contains a pool of cysteine-rich secretory proteins". Biochemical and Biophysical Research Communications. 328 (1): 177–82. doi:10.1016/j.bbrc.2004.12.154. PMID 15670767.