Lipoarabinomannan
Lipoarabinomannan, also called LAM, is a glycolipid, and a virulence factor associated with Mycobacterium tuberculosis, the bacteria responsible for tuberculosis. Its primary function is to inactivate macrophages and scavenge oxidative radicals.
The inactivation of macrophages allows for the dissemination of mycobacteria to other parts of the body. The destruction of oxidative radicals allows for the survival of the bacteria, as oxidative free radicals are an important mechanism by which our bodies try to rid ourselves of infection.
Background
Lipoarabinomannan is a lipoglycan and major virulence factor in the bacteria genus Mycobacterium. In addition to serving as a major cell wall component, it is thought to serve as a modulin with immunoregulatory and anti-inflammatory effects. This allows the bacterium to maintain survival in the human reservoir by undermining host resistance and acquired immune responses.[1] These mechanisms include the inhibition of T-cell proliferation and of macrophage microbicidal activity via diminished IFN-γ response. [2] Additional functions of Lipoarabinomannan are thought to include the neutralization of cytotoxic oxygen free radicals produced by macrophages, inhibition of protein kinase C, and induction of early response genes.[3]
Structure
Lipoarabinomannan is synthesized via addition of mannose residues to phosphoinositol by a series of mannosyltransferases to produce PIMs and lipomannan (LM).[4][5][6] PIM and LM are then glycosylated with arabinan to form LAM.[7] LAM is known to have three primary structural domains. These include a glycosylphosphatidyl anchor which attaches the molecule to the cell wall, a D-mannan core serving as a carbohydrate skeleton, and a terminal D-arabinan, also composing the carbohydrate skeleton.[7] Many arabinofuranosyl side chains branch off the mannose core.[8] It is the covalent modifications to this terminal D-arabinan that creates various LAM structures with their own unique functions to mediate bacterial survival within a host. The presence and the structure of capping allow classification of LAM molecules into three major classes.
ManLAM
Mannosylated LAMs (ManLAM) are characterized by the presence of mannosyl caps on the terminal D-arabinan. These types of LAMs are most commonly found in more pathogenic Mycobacterium species such as M. tuberculosis, M. leprae, and M. bovis. ManLAM has been shown to be an anti-inflammatory molecule that inhibits production of TNF-α and IL-12 production by human dendritic cells and human macrophages in vitro and to modulate M. tuberculosis-induced macrophage apoptosis via binding to host macrophage mannose receptors.[1][9] This is particularly important in deactivating host macrophages to allow the bacteria to survive and multiply within them.[2]
Proposed Mechanisms
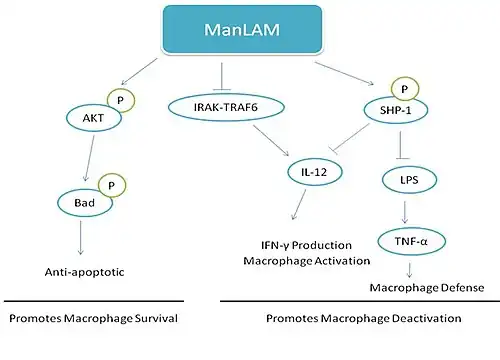
There are many proposed mechanisms behind ManLAM function. Activation of a PI3K pathway is sufficient to trigger phosphorylation of the Bcl-2 family member Bad by ManLAM. ManLAM is able to activate the serine/threonine kinase Akt via phosphorylation which is then able to phosphorylate Bad. Dephosphorylated Bad serves as a pro-apoptotic protein and its activation allows for cell survival. This demonstrates one virulence-associated mechanism by which bacteria are able to up-regulate signaling pathways to control host cell apoptosis.[8]
ManLAM may also directly activate SHP-1, a phosphotyrosine phosphatase known to be involved in terminating activation signals. SHP-1 negatively regulates pathways related to the actions of IFN-γ and insulin. LAM may regulate SHP-1 by multiple mechanisms including direct interactions, phosphorylation, and subcellular localization. Once activated, SHP-1 translocates from the cytosol to the membrane. By activating a phosphatase, LAM can inhibit LPS and IFN-γ induced protein tyrosine phosphorylation in monocytes. This decreases production of TNF-α, a molecule necessary in forming granulomas against M. tuberculosis and important in macrophage defense against bacterium via nitrogen oxide production. LAM's activation of SHP-1 also works to deactivate IL-12. IL-12 is important for innate resistance to M. tuberculosis infections. It activates natural killer cells which produce IFN-γ to activate macrophages. By impairing the function of these two molecules by SHP-1 activation, ManLAM may promote intracellular survival.[2]
Other models suggest that ManLAM acts to mediate immunosuppressive effects through suppression of LPS-induced IL-12 p40 protein production. ManLAM is thought to inhibit the IL-1 receptor-associated kinase (IRAK)-TRAF6 interaction, IκB-α phosphorylation, and nuclear translocation of c-Rel and p50 which causes reduced IL-12 p40 production.[10]
PILAM
LAMS capped with phosphoinositol are typically found in nonpathogenic species including M. smegmatis. In contrast to ManLAMs, PILAMs are pro-inflammatory. CD14, a recognition receptor present on macrophages, associate with toll-like receptor 2 (TLR2) is described to be a receptor for PILAM.[11] Binding of PILAM to the receptor elicits the activation of an intracellular signaling cascade which activates transcription factors that initiate transcription of proinflammatory cytokine genes. This may lead to TNF-α, IL-8, and IL-12 activation and apoptosis of macrophages.[1][12]
AraLAM (CheLAM)
Certain species of rapid-growing bacterium such as M. chelonae and laboratory strains (H37Ra) contain LAMs that are absent of both mannose and phosphoinosital caps.[1] This form of LAM is characterized by 1,3 –mannosyl side chains instead of the 1,2 commonly found in others mycobacterial species.[12] These forms are considered to be more potent than the mannose-capped ManLAM in inducing functions associated with macrophage activation.[9] In addition to stimulation of early genes such as c-fos, KC, and JE, AraLAM induces transcription of the mRNA for cytokines (such as TNF-α, IL 1-α, IL 1-β, IL-6, IL-8, and IL-10) characteristically produced by macrophages.[2][9] Proto-oncogenes c-fos and c-myc are involved in the regulation of gene transcription while JE and KC are peptide cytokines that serve as specific chemoattractants for neutrophils and monocytes.[13] Activation of TNF-α creates pathologic manifestations of disease such as tissue necrosis, nerve damage, and protective immunity. [14] O-acyl groups of the arabinomannan moiety may be responsible for TNF-inducing activity which causes the tuberculosis symptoms of fever, weight loss, and necrosis.[15] However, the presence of ManLAMs decreases AraLAM activity, suppressing an immune response.[9]
Point-of-care TB Diagnosis
Fujifilm SILVAMP TB LAM is a LAM based urine point-of-care test, using silver halide amplification technology.[16] Up to 60% of people with HIV are unable to produce a sputum sample, leading to delays in TB diagnosis for these patients, which often proves deadly.[17] Foundation for Innovative New Diagnostics (FIND) and Fujifilm developed the test, which is particularly useful in low-income settings, where the burden of HIV and TB is the highest. It takes an hour, doesn’t rely on electricity, and requires limited training for health workers.[18] A study with 968 HIV+ hospital inpatients found the Fujifilm SILVAMP TB LAM test to have a 28.1% higher sensitivity than the Alere Determine TB LAM Ag and the Fujifilm SILVAMP TB LAM could diagnose 65% of patients with active TB within 24 h.[19] A meta-analysis with 1,595 inpatients and outpatients showed 70.7% sensitivity and 90.9% specificity for TB diagnosis in people living with HIV for Fujifilm SILVAMP TB LAM. Further, FujiLAM showed good sensitivity for the detection of extrapulmonary TB (EPTB) ranging from 47 to 94% across different forms of ETB [20] and could have rapidly diagnosed TB in up to 89% of HIV-positive inpatients who died within 12 weeks.[21] The test showed a high positive predictive value (95.2%) in HIV-negative outpatients and has the potential to improve rapid, urine-based TB diagnosis in general populations at the point-of-care.[22] Large prospective studies are on the way.[23]
References
- 1 2 3 4 Guérardel Y, Maes E, Briken V, Chirat F, Leroy Y, Locht C, et al. (September 2003). "Lipomannan and lipoarabinomannan from a clinical isolate of Mycobacterium kansasii: novel structural features and apoptosis-inducing properties". The Journal of Biological Chemistry. 278 (38): 36637–51. doi:10.1074/jbc.M305427200. PMID 12829695.
- 1 2 3 4 Knutson KL, Hmama Z, Herrera-Velit P, Rochford R, Reiner NE (January 1998). "Lipoarabinomannan of Mycobacterium tuberculosis promotes protein tyrosine dephosphorylation and inhibition of mitogen-activated protein kinase in human mononuclear phagocytes. Role of the Src homology 2 containing tyrosine phosphatase 1". The Journal of Biological Chemistry. 273 (1): 645–52. doi:10.1074/jbc.273.1.645. PMID 9417127.
- ↑ Chan J, Fan XD, Hunter SW, Brennan PJ, Bloom BR (May 1991). "Lipoarabinomannan, a possible virulence factor involved in persistence of Mycobacterium tuberculosis within macrophages". Infection and Immunity. 59 (5): 1755–61. doi:10.1128/IAI.59.5.1755-1761.1991. PMC 257912. PMID 1850379.
- ↑ Korduláková J, Gilleron M, Mikusova K, Puzo G, Brennan PJ, Gicquel B, Jackson M (August 2002). "Definition of the first mannosylation step in phosphatidylinositol mannoside synthesis. PimA is essential for growth of mycobacteria". The Journal of Biological Chemistry. 277 (35): 31335–44. doi:10.1074/jbc.m204060200. PMID 12068013.
- ↑ Lea-Smith DJ, Martin KL, Pyke JS, Tull D, McConville MJ, Coppel RL, Crellin PK (March 2008). "Analysis of a new mannosyltransferase required for the synthesis of phosphatidylinositol mannosides and lipoarbinomannan reveals two lipomannan pools in corynebacterineae". The Journal of Biological Chemistry. 283 (11): 6773–82. doi:10.1074/jbc.m707139200. PMID 18178556.
- ↑ Morita YS, Sena CB, Waller RF, Kurokawa K, Sernee MF, Nakatani F, et al. (September 2006). "PimE is a polyprenol-phosphate-mannose-dependent mannosyltransferase that transfers the fifth mannose of phosphatidylinositol mannoside in mycobacteria". The Journal of Biological Chemistry. 281 (35): 25143–55. doi:10.1074/jbc.m604214200. PMID 16803893.
- 1 2 Guerardel Y, Maes E, Elass E, Leroy Y, Timmerman P, Besra GS, et al. (August 2002). "Structural study of lipomannan and lipoarabinomannan from Mycobacterium chelonae. Presence of unusual components with alpha 1,3-mannopyranose side chains". The Journal of Biological Chemistry. 277 (34): 30635–48. doi:10.1074/jbc.M204398200. PMID 12063260.
- 1 2 Maiti D, Bhattacharyya A, Basu J (January 2001). "Lipoarabinomannan from Mycobacterium tuberculosis promotes macrophage survival by phosphorylating Bad through a phosphatidylinositol 3-kinase/Akt pathway". The Journal of Biological Chemistry. 276 (1): 329–33. doi:10.1074/jbc.M002650200. PMID 11020382.
- 1 2 3 4 Gilleron M, Himoudi N, Adam O, Constant P, Venisse A, Rivière M, Puzo G (January 1997). "Mycobacterium smegmatis phosphoinositols-glyceroarabinomannans. Structure and localization of alkali-labile and alkali-stable phosphoinositides". The Journal of Biological Chemistry. 272 (1): 117–24. doi:10.1074/jbc.272.1.117. PMID 8995236.
- ↑ Pathak SK, Basu S, Bhattacharyya A, Pathak S, Kundu M, Basu J (December 2005). "Mycobacterium tuberculosis lipoarabinomannan-mediated IRAK-M induction negatively regulates Toll-like receptor-dependent interleukin-12 p40 production in macrophages". The Journal of Biological Chemistry. 280 (52): 42794–800. doi:10.1074/jbc.M506471200. PMID 16263713.
- ↑ Yu W, Soprana E, Cosentino G, Volta M, Lichenstein HS, Viale G, Vercelli D (October 1998). "Soluble CD14(1-152) confers responsiveness to both lipoarabinomannan and lipopolysaccharide in a novel HL-60 cell bioassay". Journal of Immunology. 161 (8): 4244–51. PMID 9780199.
- 1 2 Vignal C, Guérardel Y, Kremer L, Masson M, Legrand D, Mazurier J, Elass E (August 2003). "Lipomannans, but not lipoarabinomannans, purified from Mycobacterium chelonae and Mycobacterium kansasii induce TNF-alpha and IL-8 secretion by a CD14-toll-like receptor 2-dependent mechanism". Journal of Immunology. 171 (4): 2014–23. doi:10.4049/jimmunol.171.4.2014. PMID 12902506.
- ↑ Roach TI, Barton CH, Chatterjee D, Blackwell JM (March 1993). "Macrophage activation: lipoarabinomannan from avirulent and virulent strains of Mycobacterium tuberculosis differentially induces the early genes c-fos, KC, JE, and tumor necrosis factor-alpha". Journal of Immunology. 150 (5): 1886–96. PMID 8436823.
- ↑ Barnes PF, Chatterjee D, Brennan PJ, Rea TH, Modlin RL (April 1992). "Tumor necrosis factor production in patients with leprosy". Infection and Immunity. 60 (4): 1441–6. doi:10.1128/IAI.60.4.1441-1446.1992. PMC 257016. PMID 1548069.
- ↑ Moreno C, Taverne J, Mehlert A, Bate CA, Brealey RJ, Meager A, et al. (May 1989). "Lipoarabinomannan from Mycobacterium tuberculosis induces the production of tumour necrosis factor from human and murine macrophages". Clinical and Experimental Immunology. 76 (2): 240–5. PMC 1541837. PMID 2503277.
- ↑ "Fujifilm and FIND* Sign Development Contract to Develop Highly Sensitive Rapid Tuberculosis Diagnosis Kit for Developing Countries". Fujifilm Corporation. Archived from the original on 20 December 2018.
- ↑ "Historic UN TB declaration 'falls short' say activists". Health24. 27 September 2018.
- ↑ "Fujifilm SILVAMP TB LAM test procedure".
{{cite web}}: CS1 maint: url-status (link) - ↑ Broger T, Sossen B, du Toit E, Kerkhoff AD, Schutz C, Ivanova Reipold E, et al. (August 2019). "Novel lipoarabinomannan point-of-care tuberculosis test for people with HIV: a diagnostic accuracy study". The Lancet. Infectious Diseases. 19 (8): 852–861. doi:10.1016/s1473-3099(19)30001-5. PMC 6656794. PMID 31155318.
- ↑ Kerkhoff AD, Sossen B, Schutz C, Reipold EI, Trollip A, Moreau E, et al. (February 2020). "Diagnostic sensitivity of SILVAMP TB-LAM (FujiLAM) point-of-care urine assay for extra-pulmonary tuberculosis in people living with HIV". The European Respiratory Journal. 55 (2): 1901259. doi:10.1183/13993003.01259-2019. PMC 7002975. PMID 31699835.
- ↑ Sossen B, Broger T, Kerkhoff AD, Schutz C, Trollip A, Moreau E, et al. (January 2020). "'SILVAMP TB LAM' rapid urine tuberculosis test predicts mortality in hospitalized HIV patients in South Africa". Clinical Infectious Diseases. doi:10.1093/cid/ciaa024. PMC 8240995. PMID 31917832.
- ↑ Broger T, Nicol M, Sigal G, Gotuzzo E, Zimmer AJ, Surtie S, et al. (July 2020). "Diagnostic accuracy of three urine lipoarabinomannan tuberculosis assays in HIV-negative outpatients". The Journal of Clinical Investigation. doi:10.1172/JCI140461. PMID 32692731.
- ↑ Clinical trial number NCT04089423 for "FujiLAM Prospective Evaluation Trial" at ClinicalTrials.gov
Further reading
- Sibley LD, Adams LB, Krahenbuhl JL (April 1990). "Inhibition of interferon-gamma-mediated activation in mouse macrophages treated with lipoarabinomannan". Clinical and Experimental Immunology. 80 (1): 141–8. doi:10.1111/j.1365-2249.1990.tb06454.x. PMC 1535223. PMID 2138940.
- Ellner JJ, Daniel TM (February 1979). "Immunosuppression by mycobacterial arabinomannan". Clinical and Experimental Immunology. 35 (2): 250–7. PMC 1537646. PMID 108041.
- Chatterjee D, Khoo KH (February 1998). "Mycobacterial lipoarabinomannan: an extraordinary lipoheteroglycan with profound physiological effects". Glycobiology. 8 (2): 113–20. doi:10.1093/glycob/8.2.113. PMID 9451020.
- Chattree V, Khanna N, Bisht V, Rao DN (February 2008). "Inhibition of apoptosis, activation of NKT cell and upregulation of CD40 and CD40L mediated by M. leprae antigen(s) combined with Murabutide and Trat peptide in leprosy patients". Molecular and Cellular Biochemistry. 309 (1–2): 87–97. doi:10.1007/s11010-007-9646-8. PMID 18008143. S2CID 20518106.
- Tapping RI, Tobias PS (2003). "Mycobacterial lipoarabinomannan mediates physical interactions between TLR1 and TLR2 to induce signaling". Journal of Endotoxin Research. 9 (4): 264–8. doi:10.1177/09680519030090040801. PMID 12935358. S2CID 208323243.
- Patel VB, Bhigjee AI, Paruk HF, Singh R, Meldau R, Connolly C, et al. (November 2009). "Utility of a novel lipoarabinomannan assay for the diagnosis of tuberculous meningitis in a resource-poor high-HIV prevalence setting". Cerebrospinal Fluid Research. 6 (13): 13. doi:10.1186/1743-8454-6-13. PMC 2777116. PMID 19878608.
- Nigou J, Gilleron M, Puzo G (February 1999). "Lipoarabinomannans: characterization of the multiacylated forms of the phosphatidyl-myo-inositol anchor by NMR spectroscopy". The Biochemical Journal. 337 ( Pt 3) (3): 453–60. doi:10.1042/0264-6021:3370453. PMC 1219996. PMID 9895288.
- Venisse A, Berjeaud JM, Chaurand P, Gilleron M, Puzo G (June 1993). "Structural features of lipoarabinomannan from Mycobacterium bovis BCG. Determination of molecular mass by laser desorption mass spectrometry". The Journal of Biological Chemistry. 268 (17): 12401–11. PMID 8509380.
- Gilleron M, Garton NJ, Nigou J, Brando T, Puzo G, Sutcliffe IC (February 2005). "Characterization of a truncated lipoarabinomannan from the Actinomycete Turicella otitidis". Journal of Bacteriology. 187 (3): 854–61. doi:10.1128/JB.187.3.854-861.2005. PMC 545729. PMID 15659663.
- Prinzis S, Chatterjee D, Brennan PJ (November 1993). "Structure and antigenicity of lipoarabinomannan from Mycobacterium bovis BCG". Journal of General Microbiology. 139 (11): 2649–58. doi:10.1099/00221287-139-11-2649. PMID 8277248.
- Bhattacharjee S, Majumder N, Bhattacharyya P, Bhattacharyya S, Majumdar S (October 2007). "Immunomodulatory role of arabinosylated lipoarabinomannan on Leishmania donovani infected murine macrophages". Indian Journal of Biochemistry & Biophysics. 44 (5): 366–72. PMID 18341212.
- Garton NJ, Sutcliffe IC (February 2006). "Identification of a lipoarabinomannan-like lipoglycan in the actinomycete Gordonia bronchialis". Archives of Microbiology. 184 (6): 425–7. doi:10.1007/s00203-005-0050-z. PMID 16320036. S2CID 19740898.