Ovarian follicle activation
Ovarian follicle activation can be defined as primordial follicles in the ovary moving from a quiescent (inactive) to a growing phase. The primordial follicle in the ovary is what makes up the “pool” of follicles that will be induced to enter growth and developmental changes that change them into pre-ovulatory follicles, ready to be released during ovulation. The process of development from a primordial follicle to a pre-ovulatory follicle is called folliculogenesis.
Activation of the primordial follicle involves the following: a morphological change from flattened to cuboidal granulosa cells, proliferation of granulosa cells, formation of the protective zona pellucida layer, and growth of the oocyte.[1]
It is widely understood that androgens act primarily on preantral follicles and that this activity is important for preantral follicle growth. Additionally, it is thought that androgens are involved in primordial follicle activation. However, the influence of androgens on primordial follicle recruitment and whether this response is primary or secondary is still uncertain.
Activation of Primordial Follicle Development
Primordial follicles are activated to grow into antral follicles. Communication between the oocytes and the surrounding somatic cells, such as the granulosa cells and the theca cells, is involved in the control of primordial follicle activation. There are various activator signalling pathways that are involved in the control of ovarian follicle activation, including: Neurotropin, nerve growth factor (NGF) and its tyrosine receptor kinase (NTRK1), neurotrophin 4 (NT4), brain-derived neurotrophic factor (BDNF) and their receptor NTRK2. Additional ligands have a role in facilitating primordial follicle activation such as transforming growth factor-beta (TGF-B), growth differentiation factor 9 (GDF9) and bone morphogenic protein 15 (BMP15).
GDF9
Follicular activation rate is increased in experiments where recombinant GDF9 is added. Additionally, in vitro addition of GDF9 to human ovarian cortical tissue causes enhanced activation and follicular survival. Removing GDF9 from mice, through knock-out experiments, halts follicle progression beyond the first stage, and prevents granulosa cell proliferation. However, these GDF9 null mice have accelerated oocyte growth, suggesting that GDF9 is partially responsible for granulosa cell recruitment, as well as inhibiting oocyte growth. GDF9 promotes follicular survival and growth as a result of dampened granulosa apoptosis and follicular atresia.[2]
TGF-β
As discussed above TGF-β ligands, for example BMP4 and 7 have a role in follicular activation. SMADS are downstream molecules of the TGF-β signalling pathway, hence rely on TGF-β for activation. In the absence of SMADs, mice have decreased folliculogenesis, with decreased quantities of primordial follicles, as well as developed adult follicles at both developmental stages. BMP15 has been shown to stimulate granulosa cell growth by encouraging proliferation of undifferentiated granulosa cells. This is not dependent on FSH. It was shown that two proliferation markers, Ki-67 and proliferating cell nuclear antigen (PCNA), are regulated by these factors. Additionally, PCNA has been suggested to act as a key regulator of ovarian follicle development. The temporal expression of PCNA in oocytes is coincident with the start of primordial follicle formation. PCNA promotes apoptosis of oocytes, which regulates primordial follicle assembly.
Foxl2
Another molecule that has been implicated in the activation of oocyte follicles is Forkhead boxL2 (Foxl2). In knock out studies, it has been shown that Foxl2 may be responsible for the cuboidal transition of the pre-granulosa cells. Hence, when Foxl2 is removed, the primordial follicles are unable to develop into secondary follicles.[2]
Sohlh1
Spermatogenesis-and-oogenesis-specific basic helix-loop-helix containing protein 1 (Sohlh1) is expressed within germ cell clusters and in new primordial follicles. Knock out studies of this protein in mice show a reduced number of oocytes present at 7 weeks post birth and a malfunction in the transition from primordial to primary follicle.[2]
Repression of Primordial Follicle Activation
PTEN
Phosphatase and tensin homolog (PTEN) is a tumour suppressor gene whose actions directly affect the activation of primordial follicles. It does this by negatively controlling the PI3K/AKT/mTOR pathway.[2] This particular action of PTEN was initially discovered in an experiment using PTEN knockout mice.[2] The absence of PTEN within the primordial follicles lead to an increase in AKT phosphorylation. This then creates a subsequent rise in FOXO3 export, as AKT is no longer inhibiting its production.[3] This led to over-activation of the primordial follicles, which resulted in a premature decline of the primordial follicle pool.[2]
Foxo3
When Foxo3 is KO in mice models a huge uncontrolled activation of follicles is seen thus the mouse ovaries are deficient of the entire pool of primordial follicles because they have been prematurely activated.[2] This action is regulated by phosphorylation, the unphosphorylated form is transcriptionally active in the nucleus. However, when phosphorylation occurs the protein is transported to the cytoplasm and loses its transcriptional activity. Pelosi et al. noted that the timing and level of the Foxo3 expression is very important to regulate ovarian follicle activation.[4]
AKt- PTEN-AKt and Foxo3 are all involved in the same pathway. PTEN is situated upstream of AKt. Therefore, if PTEN is deleted specifically from an oocyte this causes an increase in AKt activity resulting in large numbers of dormant ovarian follicles resuming their growth and differentiation. The TSC complex also plays an important role in this pathways by suppressing the activity of mTOR which has been proven to be essential for maintaining dormancy.[5]
TSC and mTOR
Tuberin/tuberous sclerosis complex is also thought to be important in the regulation of primordial follicle activation. TSC negatively controls the function of mTOR (mammalian target of rapamycin). TSC knockout mice have a raised level of mTORC1 activity.[6] Suppressing mTORC1 is a necessary process to prevent primordial follicles from being prematurely activated and therefore premature ovarian insufficiency.[7]
AMH
AMH (Anti-mullerian Hormone) is a member of the transforming growth factor beta (TGF-b), that has a very important role in regulating both testicular and ovarian function. In the first instance AMH inhibits the initial enrollment of the resting primordial follicles. Secondly AMH prevents the regulation of preantral/small antral follicle growth by reducing their responsiveness to FSH.[8]
Cyclin-dependent kinase (Cdk) inhibitor p27
P27 inhibits cell cycle progression at the G1 phase[9] by preventing the action of cyclin E-Cdk2.[3] Due to its important role in the cell cycle, it is found within the nucleus of mice oocytes in primordial and primary follicles. During puberty of p27 knock out mice, all primordial follicles are activated and leads to POF. This indicates that p27 is a vital regulator in maintaining a quiescent state in primordial follicles.[6]
Medical Consequences
Premature Ovarian Failure (POF)
Premature ovarian failure (POF), or premature ovarian insufficiency (POI), is a female reproductive disorder characterised by at least 4 months of primary or secondary amenorrhea, before the age of 40.[10] It is caused by either a decrease in the primordial follicle pool, accelerated atresia of follicles or altered maturation or recruitment of primordial follicles and is associated with menopausal levels of follicle stimulating hormones, exceeding 40 Ul/L.[10],[11] Specific activator and suppressor genes are implicated in ovarian follicle activation and recent research suggests that POF may be the consequence of a genetic mutation in one or more of these genes.
FOXL2 - FOXL2 knockout mouse models showed failure of granulosa cell differentiation, which led to the premature activation and depletion of primordial follicles, characteristic of POF. Two different variations of mutations in the FOXL2 gene, which cause different forms of POF, one with earlier onset and the other with later onset and incomplete penetrance, have been identified.[12] Additionally, mutations in the FOXL2 gene have been found in approximately 5% of nonsyndromic POF patients, which suggests that FOXL2 mutations are also associated with idiopathic POF.[10]
BMP15 and GDF9 - Mutations in BMP15 and GDF9 genes can be involved in POF, but are not major causes of the disease. For example, low GDF9 mutation frequency has been found in a large cohort of Indian cases of POF.[13]
SOHLH1 - Little is known about the causative association of SOHLH1 and POF, however three novel SOHLH1 variants have been found to potentially cause the disease and when studied, they were absent in controls.[12]
AMH - A decrease in AMH expression in POF antral follicles leads to defective antral development.[14]
mTORC1 and PI3K - Deregulation of mTORC1 and PI3K signaling pathways in oocytes results in ovarian pathological conditions, including POF and subsequent infertility.[15]
PTEN - Studies of mice with a deletion in PTEN in the oocytes showed early activation of the entire pool of primordial follicles, leading to a lack of primordial follicles in adulthood, resulting in a POF phenotype.[16]
Foxo3a - Studies into mice with complete and partial Foxo3a deletions also showed premature activation of the entire primordial follicle pool, destroying the ovarian reserve and leading to oocyte death. This led to a POF phenotype, seen in studies in a range of countries.[17],[18]
TSC - In the oocytes of Tsc2 knockout mice, elevated mTORC1 activity causes the pool of primordial follicles to become activated prematurely. This results in follicle depletion in early adulthood, causing POF.[7]
Chemotherapy and Ovarian Follicle Activation
As well as having many genetic causes, premature ovarian failure has been shown to be a side effect of many chemotherapeutic agents.[19] The damage suffered by ovaries appears to be dose-dependent, and a class of chemotherapy drugs known as alkylating agents, seem to cause the most damage to the ovary and follicles. There are two ways in which this damage occurs:
- By direct damage to the primordial follicle, causing cell death by toxicity
- By indirect damage to the stromal cells which surround the follicle and support it, to allow it to grow. Loss of these supporting cells then leads to death of the follicle.
Chemotherapeutic agents, such as Cyclophosphamide, have been shown to activate the PI3K/PTEN/Akr pathway, which is the main pathway involved in keeping the follicles dormant and permitting them to grow - activation of this pathway encourages more primordial follicles to grow and develop.[19] These growing follicles may then be destroyed in subsequent rounds of chemotherapy, which often target growing cells, which will then cause more primordial follicles to differentiate and grow to replace the destroyed cells. This concept, known as burnout, leads to a depletion of the ovarian reserve and results in premature ovarian failure.
Oocyte cryopreservation
Oocyte cryopreservation is a preservative process which can be used as a way of preserving fertility in children treated for childhood or adolescent cancer, and to avoid the damage caused to the ovaries caused by cytotoxic drugs often used in chemotherapy.[20] There are several methods of cryopreservation, each with different levels of effectiveness. After cryopreservation, the ovarian tissue must be placed back into the patient, in order to allow the ovary to function normally again and regain fertility.
Restoration of Ovarian Activity after Cryopreservation
Restoration of the ovarian function occurs in almost all cases of cryopreservation, but it takes a while for the ovarian follicles to regain full function. In all cases of successful function restoration, it took 3.5-6.5 months after reimplantation before a rise in oestrogen, a key hormone produced by the ovary, and a decrease in follicle stimulating hormone (FSH) were detected. The variation in time difference may be due to differences in the follicular reserves in the women at the time of cryopreservation.[20]
Inducing follicle activation
In vitro follicle activation describes the process by which primordial follicles are deliberately activated. Oocytes from these activated follicles can be used to establish a pregnancy.[21]This has potential for widespread use in restoring fertility in women suffering with fertility issues such as early or medically-induced menopause.[22]
Medical Applications of Inducing Ovarian Follicle Growth
Diminished Ovarian Reserve/ Premature Ovarian Insufficiency
Conventional IVA or drug free IVA can be used to preserve fertility in patients with Diminished Ovarian Reserve (see Poor Ovarian Reserve) or Premature Ovarian Insufficiency (POI).[23]
Drug free IVA is more beneficial in patients with DOR or POI with recent menses cessation than conventional IVA. This is because it reduces the invasiveness of surgical approaches and circumvents adverse effects of tissue culture on follicles.[24] Traditional IVA is recommended in patients with low ovarian reserve like in patients with POI with prolonged menopause or patients looking for immediate outcomes.[23]
Using drug free IVA, at least a total of 18 healthy live births and three more ongoing pregnancies in POI and DOR patients charted in papers, with additional unpublished deliveries and ongoing pregnancies reported in scientific conferences.[23]
Resistant Ovary Syndrome
Patients with Resistant Ovary Syndrome (ROS) are a subgroup of patients with POI who have multiple antral follicles present in the ovary as shown by ultrasound monitoring. These follicles secrete low levels of oestrogen but are resistant to exogenous follicle stimulating hormone treatment so cannot develop to maturity.[24]
Like patients with POI, ROS patients can respond to incision of the ovarian cortex to cause Hippo signalling disruption. Additionally, laparoscopic ovarian incision (LOI) can be performed in vivo to successfully promote follicle growth without removing tissue outside of the body.[24]
Post-cancer treatment
Oocyte cryopreservation is widely used to protect primordial follicles from gonadotoxic treatment but some patients are not eligible for this, IVA could be a beneficial option for fertility preservation in cancer patients. The IVA approach could maximise the number of secondary oocytes in infertile woman since transplanting ovarian tissue successfully activates the dormant follicle pool.[23]
The IVA method focuses on the short term goal of generating a large number of activated follicles immediately following transplantation of tissue back into the body. [24] This means that the IVA method has a reduced graft lifespan compared to the 4-5 years that can be offered in the cryopreservation method. The graft lifespan relates to the length of time following transplantation back into he body that pregnancies can occur within.[25] This is because IVA promotes loss of dormant follicles to activation so fewer dormant follicles will remain in the graft. This, as a result, reduces graft lifespan.[25] However, this can be beneficial in patients looking for immediate outcomes or to improve her chance of conceiving as fast as possible due to advancing age.[23]
So far, there have been no reported live births but progress is being made in in-vitro research.[23]
Obtaining the ovaries
Surgically removing ovaries
Patients are usually treated with synthetic oestrogen and progesterone for at least a month in order to suppress their gonadotrophin levels which has been shown to promote ovulation.[26] A laparoscopic approach is used to remove the ovary. The outer part of the ovary (the cortex) is removed using scissors as this minimises any damage to the sensitive inner ovarian tissue. This tissue is then analysed to assess the presence of existing follicles before further manipulation.[27]
Artificial ovaries
Artificial ovaries can be used to provide follicles for artificial activation.[2] Follicles from these synthetic ovaries have been shown to support successful pregnancies in mouse models and show promise for potential for future treatments in humans.[2]
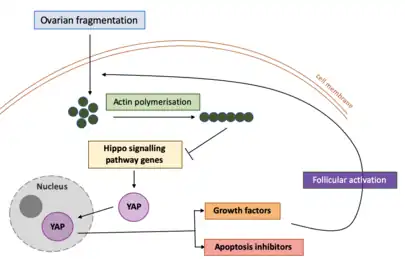
Ovarian Fragmentation and Hippo Signalling Disruption
The Hippo signalling pathway is an essential pathway involving several components that regulate cell growth, survival and stem cell renewal.[28] For this reason, the Hippo signalling pathway has been implicated in the maintenance and control of organ size in organisms.
In recent years, ovarian fragmentation has surfaced as an effective strategy to promote ovarian follicular activation and growth.[2] Studies undertaken on cultured human ovarian cortical fragments have demonstrated that the mechanical damage resulting from ovarian tissue fragmentation disrupts the Hippo signalling pathway. This has been found to accelerate follicle activation. Mechanical damage to the ovaries through fragmentation have been found to induce a transient elevation in actin polymerisation - a process where small actin molecules (proteins) combine to form a large, chain-like structure with repeating actin units.[2] Mechanical damage to the ovaries has also decreased phosphorylation of Yes-associated protein (YAP), an essential compound in the Hippo Signalling pathway.[29] Phosphorylation refers to the addition of phosphate groups to molecules. Consequently, this leads to an increase in the levels of YAP in the nucleus. This in turn triggers a downstream ripple effect to increase the expression of growth factors and apoptosis inhibitors, therefore promoting cell growth and survival.
PI3K/AKT/MTOR Signalling Pathway Activation
Another key pathway in ovarian follicle activation is the PI3K-PTEN-AKT-FOXO3 pathway.
The PI3K-PTEN-AKT-FOXO3 pathway is a complex signal transduction pathway involving a cascade of several molecules. This pathway is involved in promoting growth, survival and proliferation of cells in response to certain factors.[30] The pathway is therefore implicated in the activation of ovarian primordial follicles. In in vitro activation of ovarian follicles, drugs such as inhibitors of PTEN or activators of PI3K can be administered to stimulate follicle activation.
In vitro activation (IVA)
Stimulation of the Atk pathway activates dormant primordial follicles. The hippo signalling pathway regulates growth by inhibiting cell proliferation and promoting apoptosis, disrupting this pathway stimulates cell growth and proliferation.[2] In vitro activation (IVA) consists of the combination of disrupting the hippo signalling pathway and stimulation of Akt signalling to activate ovarian follicle growth and maturation.[2][3]Atk pathway activation can be artificially induced by Atk stimulating drugs such as PTEN inhibitors and PI3K activators and hippo signalling disruption is achieved through fragmentation of ovarian cortex via various methods.[2] Growth of follicles can be seen in subsequent autografts, mature eggs can be retrieved and IVF carried out making it a good candidate for fertility preservation. Drug free IVA may alternatively be used. This is the same as IVA however no drugs are used to stimulate the Atk pathway, only ovarian cortex fragmentation is used.[3]
References
- ↑ Braw-Tal R (February 2002). "The initiation of follicle growth: the oocyte or the somatic cells?". Molecular and Cellular Endocrinology. 187 (1–2): 11–8. doi:10.1016/s0303-7207(01)00699-2. PMID 11988306. S2CID 1417914.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Kim JY (March 2012). "Control of ovarian primordial follicle activation". Clinical and Experimental Reproductive Medicine. 39 (1): 10–4. doi:10.5653/cerm.2012.39.1.10. PMC 3341446. PMID 22563545.
- 1 2 3 4 Hsueh AJ, Kawamura K, Cheng Y, Fauser BC (February 2015). "Intraovarian control of early folliculogenesis". Endocrine Reviews. 36 (1): 1–24. doi:10.1210/er.2015.36.issue-1.edboard. PMC 4309737. PMID 25202833.
- ↑ Pelosi E, Omari S, Michel M, Ding J, Amano T, Forabosco A, et al. (2013-05-14). "Constitutively active Foxo3 in oocytes preserves ovarian reserve in mice". Nature Communications. 4: 1843. Bibcode:2013NatCo...4.1843P. doi:10.1038/ncomms2861. PMC 4504230. PMID 23673628.
- ↑ Zhang X, Tang N, Hadden TJ, Rishi AK (November 2011). "Akt, FoxO and regulation of apoptosis". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1813 (11): 1978–86. doi:10.1016/j.bbamcr.2011.03.010. PMID 21440011.
- 1 2 Reddy P, Zheng W, Liu K (February 2010). "Mechanisms maintaining the dormancy and survival of mammalian primordial follicles". Trends in Endocrinology and Metabolism. 21 (2): 96–103. doi:10.1016/j.tem.2009.10.001. PMID 19913438. S2CID 27867193.
- 1 2 Adhikari D, Flohr G, Gorre N, Shen Y, Yang H, Lundin E, et al. (December 2009). "Disruption of Tsc2 in oocytes leads to overactivation of the entire pool of primordial follicles". Molecular Human Reproduction. 15 (12): 765–70. doi:10.1093/molehr/gap092. PMID 19843635.
- ↑ Chang HM, Klausen C, Leung PC (August 2013). "Antimüllerian hormone inhibits follicle-stimulating hormone-induced adenylyl cyclase activation, aromatase expression, and estradiol production in human granulosa-lutein cells". Fertility and Sterility. 100 (2): 585–92.e1. doi:10.1016/j.fertnstert.2013.04.019. PMID 23663993.
- ↑ Møller MB (September 2000). "P27 in cell cycle control and cancer". Leukemia & Lymphoma. 39 (1–2): 19–27. doi:10.3109/10428190009053535. PMID 10975380. S2CID 41764123.
- 1 2 3 Chapman C, Cree L, Shelling AN (2015-09-23). "The genetics of premature ovarian failure: current perspectives". International Journal of Women's Health. 7: 799–810. doi:10.2147/IJWH.S64024. PMC 4590549. PMID 26445561.
- ↑ Persani L, Rossetti R, Cacciatore C (November 2010). "Genes involved in human premature ovarian failure". Journal of Molecular Endocrinology. 45 (5): 257–79. doi:10.1677/JME-10-0070. PMID 20668067.
- 1 2 Harris SE, Chand AL, Winship IM, Gersak K, Aittomäki K, Shelling AN (August 2002). "Identification of novel mutations in FOXL2 associated with premature ovarian failure". Molecular Human Reproduction. 8 (8): 729–33. doi:10.1093/molehr/8.8.729. PMID 12149404.
- ↑ Laissue P, Christin-Maitre S, Touraine P, Kuttenn F, Ritvos O, Aittomaki K, et al. (May 2006). "Mutations and sequence variants in GDF9 and BMP15 in patients with premature ovarian failure". European Journal of Endocrinology. 154 (5): 739–44. doi:10.1530/eje.1.02135. PMID 16645022.
- ↑ Méduri G, Massin N, Guibourdenche J, Bachelot A, Fiori O, Kuttenn F, et al. (January 2007). "Serum anti-Müllerian hormone expression in women with premature ovarian failure". Human Reproduction. 22 (1): 117–23. doi:10.1093/humrep/del346. PMID 16954410.
- ↑ Adhikari D, Zheng W, Shen Y, Gorre N, Hämäläinen T, Cooney AJ, et al. (February 2010). "Tsc/mTORC1 signaling in oocytes governs the quiescence and activation of primordial follicles". Human Molecular Genetics. 19 (3): 397–410. doi:10.1093/hmg/ddp483. PMC 2798719. PMID 19843540.
- ↑ Reddy P, Liu L, Adhikari D, Jagarlamudi K, Rajareddy S, Shen Y, et al. (February 2008). "Oocyte-specific deletion of Pten causes premature activation of the primordial follicle pool". Science. 319 (5863): 611–3. doi:10.1126/science.1152257. PMID 18239123. S2CID 31102184.
- ↑ Watkins WJ, Umbers AJ, Woad KJ, Harris SE, Winship IM, Gersak K, Shelling AN (November 2006). "Mutational screening of FOXO3A and FOXO1A in women with premature ovarian failure". Fertility and Sterility. 86 (5): 1518–21. doi:10.1016/j.fertnstert.2006.03.054. PMID 16979636.
- ↑ Wang B, Mu Y, Ni F, Zhou S, Wang J, Cao Y, Ma X (April 2010). "Analysis of FOXO3 mutation in 114 Chinese women with premature ovarian failure". Reproductive Biomedicine Online. 20 (4): 499–503. doi:10.1016/j.rbmo.2010.01.008. PMID 20219431.
- 1 2 Kalich-Philosoph L, Roness H, Carmely A, Fishel-Bartal M, Ligumsky H, Paglin S, et al. (May 2013). "Cyclophosphamide triggers follicle activation and "burnout"; AS101 prevents follicle loss and preserves fertility". Science Translational Medicine. 5 (185): 185ra62. doi:10.1126/scitranslmed.3005402. PMID 23677591. S2CID 206681545.
- 1 2 Donnez J, Dolmans MM, Pellicer A, Diaz-Garcia C, Sanchez Serrano M, Schmidt KT, et al. (May 2013). "Restoration of ovarian activity and pregnancy after transplantation of cryopreserved ovarian tissue: a review of 60 cases of reimplantation". Fertility and Sterility. 99 (6): 1503–13. doi:10.1016/j.fertnstert.2013.03.030. PMID 23635349.
- ↑ Devenutto L, Quintana R, Quintana T (2020). "In vitro activation of ovarian cortex and autologous transplantation: A novel approach to primary ovarian insufficiency and diminished ovarian reserve". Human Reproduction Open. 2020 (4): hoaa046. doi:10.1093/hropen/hoaa046. PMC 7668396. PMID 33225075.
- ↑ Niederberger C, Pellicer A, Cohen J, Gardner DK, Palermo GD, O'Neill CL, et al. (July 2018). "Forty years of IVF". Fertility and Sterility. 110 (2): 185–324.e5. doi:10.1016/j.fertnstert.2018.06.005. PMID 30053940. S2CID 51725993.
- 1 2 3 4 5 6 Vo KC, Kawamura K (April 2021). "In Vitro Activation Early Follicles: From the Basic Science to the Clinical Perspectives". International Journal of Molecular Sciences. 22 (7): 3785. doi:10.3390/ijms22073785. PMC 8038686. PMID 33917468.
- 1 2 3 4 Hsueh AJ, Kawamura K (September 2020). "Hippo signaling disruption and ovarian follicle activation in infertile patients". Fertility and Sterility. 114 (3): 458–464. doi:10.1016/j.fertnstert.2020.07.031. PMID 32782158. S2CID 221108206.
- 1 2 Meirow D, Roness H, Kristensen SG, Andersen CY (November 2015). "Optimizing outcomes from ovarian tissue cryopreservation and transplantation; activation versus preservation". Human Reproduction. 30 (11): 2453–6. doi:10.1093/humrep/dev210. PMID 26345687.
- ↑ Bidet M, Bachelot A, Touraine P (August 2008). "Premature ovarian failure: predictability of intermittent ovarian function and response to ovulation induction agents". Current Opinion in Obstetrics & Gynecology. 20 (4): 416–20. doi:10.1097/GCO.0b013e328306a06b. PMID 18660695. S2CID 20196412.
- ↑ Fabregues F, Ferreri J, Calafell JM, Moreno V, Borrás A, Manau D, Carmona F (August 2018). "Pregnancy after drug-free in vitro activation of follicles and fresh tissue autotransplantation in primary ovarian insufficiency patient: a case report and literature review". Journal of Ovarian Research. 11 (1): 76. doi:10.1186/s13048-018-0447-3. PMC 6119245. PMID 30170634.
- ↑ Ma S, Meng Z, Chen R, Guan KL (June 2019). "The Hippo Pathway: Biology and Pathophysiology". Annual Review of Biochemistry. 88 (1): 577–604. doi:10.1146/annurev-biochem-013118-111829. PMID 30566373. S2CID 58642425.
- ↑ Vo KC, Kawamura K (2021). "Ovarian Fragmentation and AKT Stimulation for Expansion of Fertile Lifespan". Frontiers in Reproductive Health. 3: 3. doi:10.3389/frph.2021.636771. ISSN 2673-3153.
- ↑ Hsueh AJ, Kawamura K, Cheng Y, Fauser BC (February 2015). "Intraovarian control of early folliculogenesis". Endocrine Reviews. 36 (1): 1–24. doi:10.1210/er.2014-1020. PMC 4309737. PMID 25202833.