Pulsatile secretion
Pulsatile secretion is a biochemical phenomenon observed in a wide variety of cell and tissue types, in which chemical products are secreted in a regular temporal pattern. The most common cellular products observed to be released in this manner are intercellular signaling molecules such as hormones or neurotransmitters. Examples of hormones that are secreted pulsatilely include insulin, thyrotropin, TRH, gonadotropin-releasing hormone (GnRH) and growth hormone (GH). In the nervous system, pulsatility is observed in oscillatory activity from central pattern generators. In the heart, pacemakers are able to work and secrete in a pulsatile manner. A pulsatile secretion pattern is critical to the function of many hormones in order to maintain the delicate homeostatic balance necessary for essential life processes, such as development and reproduction. Variations of the concentration in a certain frequency can be critical to hormone function, as evidenced by the case of GnRH agonists, which cause functional inhibition of the receptor for GnRH due to profound downregulation in response to constant (tonic) stimulation. Pulsatility may function to sensitize target tissues to the hormone of interest and upregulate receptors, leading to improved responses. This heightened response may have served to improve the animal's fitness in its environment and promote its evolutionary retention.
Pulsatile secretion in its various forms is observed in:
- Hypothalamic-Pituitary-Gonadal axis (HPG) related hormones
- Glucocorticoids
- Insulin
- Growth Hormone
- Parathyroid hormone
Neuroendocrine Pulsatility
Nervous system control over hormone release is based in the hypothalamus, from which the neurons that populate the pariventricular and arcuate nuclei originate.[1] These neurons project to the median eminence, where they secrete releasing hormones into the hypophysial portal system connecting the hypothalamus with the pituitary gland. There, they dictate endocrine function via the four Hypothalamic-Pituitary-Glandular axes.[1] Recent studies have begun to offer evidence that many pituitary hormones which have been observed to be released episodically are preceded by pulsatile secretion of their associated releasing hormone from the hypothalamus in a similar pulsatile fashion. Novel research into the cellular mechanisms associated with pituitary hormone pulsatility, such as that observed for Leutinizing Hormone (LH) and Follicle Stimulating Hormone (FSH), have indicated similar pulses into the hypophyseal vessels of Gonadotropin Releasing Hormone (GnRH).[2][3]
Luteinizing Hormone & Follicle Stimulating Hormone (HPG axis)
LH is released from the pituitary gland along with FSH in response to GnRH release into the hypophyseal portal system.[4] Pulsatile GnRH release causes pulsatile LH and FSH release to occur, which modulates and maintains appropriate levels of bioavailable gonadal hormone: testosterone in males and estradiol in females subject to the requirements of a superior feedback loop.[3] In females the levels of LH is typically 1–20 IU/L during the reproductive period and is estimated to be 1.8–8.6 IU/L in males over 18 years of age.[5][6][7]
ACTH and Glucocorticoids (HPA axis)
Regular pulses of glucocorticoids, mainly cortisol in the case of humans, are released regularly from the adrenal cortex following a circadian pattern in addition to their release as a part of the stress response.[8][9] Cortisol release follows a high frequency of pulses forming an ultradian rhythm, with amplitude being the primary variation in its release, so that the signal is amplitude modulated.[8] Glucocorticoid pulsatlity has been observed to follow a circadian rhythm, with highest levels observed before waking and before anticipated mealtimes.[8][9] This pattern in amplitude of release is observed to be consistent across vertebrates.[9] Studies done in humans, rats, and sheep have also observed a similar circadian pattern of release of adrenocorticotropin (ACTH) shortly preceding the pulse in the resulting corticosteroid.[8] It is currently hypothesized that the observed pulsatility of ACTH and glucocorticoids is driven via pulsatility of corticotropin-releasing hormone (CRH), however there exists little data to support this due to difficulty in measuring CRH.[8]
Thyrotropin and thyroid hormones (HPT axis)
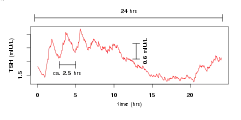
The secretion pattern of thyrotropin (TSH) is shaped by infradian, circadian and ultradian rhythms. Infradian rhythmis are mainly represented by circannual variation mirroring the seasonality of thyroid function.[10] Circadian rhythms lead to a peak secretion (acrophase) around midnight and nadir concentrations around noon and in the early afternoon.[11][12] A similar pattern is observed for triiodothyronine, however with a phase shift.[12] Pulsatile release contributes to the ultradian rhythm of TSH concentration with about 10 pulses per 24 hours.[13][14][15] The amplitude of the circadian and ultradian rhythms is reduced in severe non-thyroidal illness syndrome (TACITUS).[16][17]
Contemporary theories assume that autocrine and paracrine (ultrashort) feedback mechanisms controlling TSH secretion within the anterior pituitary gland are a major factor contributing to the evolution of its pulsatility.[18][19][20]
Insulin

Pulsatile insulin secretion from individual beta cells is driven by oscillation of the calcium concentration in the cells. In beta cells lacking contact (i.e. outside islet of Lagerhans), the periodicity of these oscillations is rather variable (2-10 min). However, within an islet of Langerhans, the oscillations become synchronized by electrical coupling between closely located beta cells that are connected by gap junctions, and the periodicity is more uniform (3-6 min).[21] In addition to gap junctions, pulse coordination is managed by ATP signaling. α and δ cells in the pancreas also share secrete factors in a similar pulsatile manner.[22]
References
- 1 2 Kandel ER, Jessell TM, Schwartz JH, Siegelbaum SA, Hudspeth AJ (2013). Principles of neural science (5th ed.). New York. ISBN 9780071390118. OCLC 795553723.
- ↑ Wetsel WC, Valença MM, Merchenthaler I, Liposits Z, López FJ, Weiner RI, Mellon PL, Negro-Vilar A (May 1992). "Intrinsic pulsatile secretory activity of immortalized luteinizing hormone-releasing hormone-secreting neurons". Proceedings of the National Academy of Sciences of the United States of America. 89 (9): 4149–53. Bibcode:1992PNAS...89.4149W. doi:10.1073/pnas.89.9.4149. PMC 525650. PMID 1570341.
- 1 2 Stamatiades GA, Kaiser UB (March 2018). "Gonadotropin regulation by pulsatile GnRH: Signaling and gene expression". Molecular and Cellular Endocrinology. 463: 131–141. doi:10.1016/j.mce.2017.10.015. PMC 5812824. PMID 29102564.
- ↑ Molina, Patricia E. (9 April 2018). Endocrine physiology. Preceded by: Molina, Patricia E. (Fifth ed.). [New York]. ISBN 978-1-260-01936-0. OCLC 1026417940.
- ↑ Mayo Medical Laboratories - Test ID: LH, Luteinizing Hormone (LH), Serum Archived 2016-09-25 at the Wayback Machine, retrieved December 2012
- ↑ World Health Organization Proposed International Standard for Luteinizing Hormone. WHO Expert Committee on Biological Standardization. World Health Organization. Geneva. 2003.
- ↑ WHO International Standard, Luteinizing Hormone, Human, Recombinant. National Institute for Biological Standards and Control.
- 1 2 3 4 5 Gjerstad JK, Lightman SL, Spiga F (September 2018). "Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility". Stress. 21 (5): 403–416. doi:10.1080/10253890.2018.1470238. PMC 6220752. PMID 29764284.
- 1 2 3 Kalsbeek A, van der Spek R, Lei J, Endert E, Buijs RM, Fliers E (February 2012). "Circadian rhythms in the hypothalamo-pituitary-adrenal (HPA) axis". Molecular and Cellular Endocrinology. 349 (1): 20–9. doi:10.1016/j.mce.2011.06.042. PMID 21782883. S2CID 33843620.
- ↑ Tendler, A; Bar, A; Mendelsohn-Cohen, N; Karin, O; Korem Kohanim, Y; Maimon, L; Milo, T; Raz, M; Mayo, A; Tanay, A; Alon, U (16 February 2021). "Hormone seasonality in medical records suggests circannual endocrine circuits". Proceedings of the National Academy of Sciences of the United States of America. 118 (7): e2003926118. doi:10.1073/pnas.2003926118. PMC 7896322. PMID 33531344.
- ↑ Lucke, C; Hehrmann, R; von Mayersbach, K; von zur Mühlen, A (September 1977). "Studies on circadian variations of plasma TSH, thyroxine and triiodothyronine in man". Acta Endocrinologica. 86 (1): 81–8. doi:10.1530/acta.0.0860081. PMID 578614.
- 1 2 Russell, W; Harrison, RF; Smith, N; Darzy, K; Shalet, S; Weetman, AP; Ross, RJ (June 2008). "Free triiodothyronine has a distinct circadian rhythm that is delayed but parallels thyrotropin levels". The Journal of Clinical Endocrinology and Metabolism. 93 (6): 2300–6. doi:10.1210/jc.2007-2674. PMID 18364382.
- ↑ Greenspan, SL; Klibanski, A; Schoenfeld, D; Ridgway, EC (September 1986). "Pulsatile secretion of thyrotropin in man". The Journal of Clinical Endocrinology and Metabolism. 63 (3): 661–8. doi:10.1210/jcem-63-3-661. PMID 3734036.
- ↑ Brabant, G; Prank, K; Ranft, U; Schuermeyer, T; Wagner, TO; Hauser, H; Kummer, B; Feistner, H; Hesch, RD; von zur Mühlen, A (February 1990). "Physiological regulation of circadian and pulsatile thyrotropin secretion in normal man and woman". The Journal of Clinical Endocrinology and Metabolism. 70 (2): 403–9. doi:10.1210/jcem-70-2-403. PMID 2105332.
- ↑ Samuels, MH; Veldhuis, JD; Henry, P; Ridgway, EC (August 1990). "Pathophysiology of pulsatile and copulsatile release of thyroid-stimulating hormone, luteinizing hormone, follicle-stimulating hormone, and alpha-subunit". The Journal of Clinical Endocrinology and Metabolism. 71 (2): 425–32. doi:10.1210/jcem-71-2-425. PMID 1696277.
- ↑ Adriaanse, R; Romijn, JA; Brabant, G; Endert, E; Wiersinga, WM (November 1993). "Pulsatile thyrotropin secretion in nonthyroidal illness". The Journal of Clinical Endocrinology and Metabolism. 77 (5): 1313–7. doi:10.1210/jcem.77.5.8077326. PMID 8077326.
- ↑ Chatzitomaris, A; Hoermann, R; Midgley, JE; Hering, S; Urban, A; Dietrich, B; Abood, A; Klein, HH; Dietrich, JW (2017). "Thyroid Allostasis-Adaptive Responses of Thyrotropic Feedback Control to Conditions of Strain, Stress, and Developmental Programming". Frontiers in Endocrinology. 8: 163. doi:10.3389/fendo.2017.00163. PMC 5517413. PMID 28775711.
- ↑ Dietrich, J. W.; Tesche, A.; Pickardt, C. R.; Mitzdorf, U. (June 2004). "Thyrotropic Feedback Control: Evidence for an Additional Ultrashort Feedback Loop from Fractal Analysis". Cybernetics and Systems. 35 (4): 315–331. doi:10.1080/01969720490443354. S2CID 13421388.
- ↑ Dietrich, JW; Landgrafe, G; Fotiadou, EH (2012). "TSH and Thyrotropic Agonists: Key Actors in Thyroid Homeostasis". Journal of Thyroid Research. 2012: 351864. doi:10.1155/2012/351864. PMC 3544290. PMID 23365787.
- ↑ Hoermann, R; Midgley, JE; Larisch, R; Dietrich, JW (2015). "Homeostatic Control of the Thyroid-Pituitary Axis: Perspectives for Diagnosis and Treatment". Frontiers in Endocrinology. 6: 177. doi:10.3389/fendo.2015.00177. PMC 4653296. PMID 26635726.
- 1 2 Hellman B, Gylfe E, Grapengiesser E, Dansk H, Salehi A (August 2007). "[Insulin oscillations--clinically important rhythm. Antidiabetics should increase the pulsative component of the insulin release]". Lakartidningen. 104 (32–33): 2236–9. PMID 17822201.
- ↑ Hellman B (2009). "Pulsatility of insulin release--a clinically important phenomenon". Upsala Journal of Medical Sciences. 114 (4): 193–205. doi:10.3109/03009730903366075. PMC 2852781. PMID 19961265.