Magnaporthe grisea
| Magnaporthe grisea | |
|---|---|
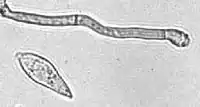 | |
| A conidium and conidiogenous cell of M. grisea | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | Ascomycota |
| Class: | Sordariomycetes |
| Order: | Magnaporthales |
| Family: | Magnaporthaceae |
| Genus: | Magnaporthe |
| Species: | M. grisea |
| Binomial name | |
| Magnaporthe grisea (T.T. Hebert) M.E. Barr | |
| Synonyms | |
|
Ceratosphaeria grisea T.T. Hebert, (1971) | |
Magnaporthe grisea, also known as rice blast fungus, rice rotten neck, rice seedling blight, blast of rice, oval leaf spot of graminea, pitting disease, ryegrass blast, Johnson spot,[1][2][3][4][5][6][7] neck blast,[8][9][10][11] and Imochi (Japanese:稲熱) is a plant-pathogenic fungus and model organism[12] that causes a serious disease affecting rice. It is now known that M. grisea consists of a cryptic species complex containing at least two biological species that have clear genetic differences and do not interbreed.[13] Complex members isolated from Digitaria have been more narrowly defined as M. grisea. The remaining members of the complex isolated from rice and a variety of other hosts have been renamed Magnaporthe oryzae, within the same M. grisea complex.[13] Confusion on which of these two names to use for the rice blast pathogen remains, as both are now used by different authors.
Members of the Magnaporthe grisea complex can also infect other agriculturally important cereals including wheat, rye, barley, and pearl millet causing diseases called blast disease or blight disease. Rice blast causes economically significant crop losses annually. Each year it is estimated to destroy enough rice to feed more than 60 million people. The fungus is known to occur in 85 countries worldwide[14] and as of 2003 was the most devastating fungal plant pathogen in the world.[12]
Hosts and symptoms


M. grisea is an ascomycete fungus. It is an extremely effective plant pathogen as it can reproduce both sexually and asexually to produce specialized infectious structures known as appressoria that infect aerial tissues and hyphae that can infect root tissues.
Rice blast has been observed on rice strains M-201, M-202, M-204, M-205, M-103, M-104, S-102, L-204, Calmochi-101, with M-201 being the most vulnerable.[15] Initial symptoms are white to gray-green lesions or spots with darker borders produced on all parts of the shoot, while older lesions are elliptical or spindle-shaped and whitish to gray with necrotic borders. Lesions may enlarge and coalesce to kill the entire leaf. Symptoms are observed on all above-ground parts of the plant.[16] Lesions can be seen on the leaf collar, culm, culm nodes, and panicle neck node. Internodal infection of the culm occurs in a banded pattern. Nodal infection causes the culm to break at the infected node (rotten neck).[17] It also affects reproduction by causing the host to produce fewer seeds. This is caused by the disease preventing maturation of the actual grain.[14]
Disease cycle
The pathogen infects as a spore that produces lesions or spots on parts of the rice plant such as the leaf, leaf collar, panicle, culm and culm nodes. Using a structure called an appressorium, the pathogen penetrates the plant. The appressorium cell wall is chitinous and its inner side contains melanin,[T 1] which is necessary to damage host structures.[T 2][12] The turgor pressure generated during this process is sufficient to penetrate the plants' cuticles routinely, and experimentally can penetrate Kevlar. This impressive turgor is produced by synthesis of glycerol and maintained by the aforementioned appressorial melanin.[12] The pathogen is able to move between the plant cells using its invasive hyphae to enter through plasmodesmata.[18] M. grisea then sporulates from the diseased rice tissue to be dispersed as conidiospores.[19] After overwintering in sources such as rice straw and stubble, the cycle repeats.[14]
A single cycle can be completed in about a week under favorable conditions where one lesion can generate up to thousands of spores in a single night. Disease lesions, however, can appear in three to four days after infection.[20] With the ability to continue to produce the spores for over 20 days, rice blast lesions can be devastating to susceptible rice crops.[21]
Environment
Rice blast is a significant problem in temperate regions and can be found in areas such as irrigated lowland and upland.[22] Conditions conducive for rice blast include long periods of free moisture and/or high humidity, because leaf wetness is required for infection.[22] Sporulation increases with high relative humidity and at 77–82 °F (25–28 °C), spore germination, lesion formation, and sporulation are at optimum levels.[14]
In terms of control, excessive use of nitrogen fertilization as well as drought stress increase rice susceptibility to the pathogen as the plant is placed in a weakened state and its defenses are low.[14] Flooding and draining fields is normal in rice growing, however leaving a field drained for extended periods also favors infection as that will aerate the soil, converting ammonium to nitrate and thus causing stress to rice crops, as well.[14]
Geographical distribution
Wheat blast was found in the 2017-2018 rainy season in Zambia, in the Mpika district of the Muchinga Province.[23][24]
In February 2016 a devastating wheat epidemic struck Bangladesh.[25][26] Transcriptome analysis showed this to be an M. grisea lineage most likely from Minas Gerais, São Paulo, Brasília, and Goiás states of Brazil and not from any geographically proximate strains.[25][26] This successful diagnosis shows the ability of genetic surveillance to untangle the novel biosecurity implications of transcontinental transportation[25][26] and allows the Brazilian experience to be rapidly applied to the Bangladeshi situation.[25][26] To that end the government has set up an early warning system to track its spread through the country.[26]
Management

The fungus has been able to establish both resistance to chemical treatments and virulence to genetic resistance in some types of rice developed by plant breeders. It is thought that the fungus can achieve this by genetic change through mutation. In order to most effectively control infection by M. grisea, an integrated management program should be implemented to avoid overuse of a single control method and fight against genetic resistance. For example, eliminating crop residue could reduce the occurrence of overwintering and discourage inoculation in subsequent seasons. Another strategy would be to plant resistant rice varieties that are not as susceptible to infection by M. grisea.[14] Knowledge of the pathogenicity of M. grisea and its need for free moisture suggest other control strategies such as regulated irrigation and a combination of chemical treatments with different modes of action.[14] Managing the amount of water supplied to the crops limits spore mobility thus dampening the opportunity for infection. Chemical controls such as Carpropamid have been shown to prevent penetration of the appressoria into rice epidermal cells, leaving the grain unaffected.[27] Papajani et al 2015 finds the essential oils of both Origanum vulgare and Rosmarinus officinalis to be effective in vitro, and provides treatment thresholds.[28]: 107–108
Importance
Rice blast is the most important disease concerning rice crops in the world. Since rice is an important food source for much of the world, its effects have a broad range. It has been found in over 85 countries across the world and reached the United States in 1996. Every year the amount of crops lost to rice blast could feed 60 million people. Although there are some resistant strains of rice, the disease persists wherever rice is grown. The disease has never been eradicated from a region.[29]
Strains
Three strains, albino (defined by a mutation at the ALB1 locus), buff (BUF1), and rosy (RSY1) have been extensively studied because they are nonpathogenic. This has been found to be due to nonuse of melanin, which is a virulence factor in M. grisea.[T 2]
Genetics
Whole-genome sequences were just becoming possible, and being made available, in 2003.[12]
A mitogen-activated protein kinase (MAPK) called pmk1 is genetically close to one necessary for mating and cell morphology in yeasts, FUS3/KSS1. Defective mutant yeast are somewhat or entirely restored in mating function if they are given a copy of pmk1. It was therefore assumed that this must only be a mating and development gene in M. grisea, however it turns out to be both vital to the female mating process and in appressorium function and pathogenicity as a whole.[12]
Because signal links between MAPKs and cyclic adenosine monophosphates were shown to be required for mating in several other models, including Ustilago maydis and several others, this was assumed to be true for M. grisea, and yet that was then shown to be unnecessary in this model. This demonstrates significant variety in cellular function within fungi.[12]
The transaminase alanine: glyoxylate aminotransferase 1 (AGT1) has been shown to be crucial to the pathogenicity of M. grisea through its maintenance of redox homeostasis in peroxisomes. Lipids transported to the appressoria during host penetration are degraded within a large central vacuole, a process that produces fatty acids. β-Oxidation of fatty acids is an energy producing process that generates Acetyl-CoA and the reduced molecules FADH2 and NADH, which must be oxidized in order to maintain redox homeostasis in appressoria. AGT1 promotes lactate fermentation, oxidizing NADH/FADH2 in the process.[30]
M. grisea mutants lacking the AGT1 gene were observed to be nonpathogenic through their inability to penetrate host surface membranes. This indicates the possibility of impaired lipid utilization in M. grisea appressoria in the absence of the AGT1 gene. [31]
See also
- Corn grey leaf spot, a similar disease in maize/corn
- Gray leaf spot, a similar disease in other grasses
References
- ↑ Talbot, N. J. (2003). "ON THE TRAIL OF A CEREAL KILLER: Exploring the Biology of Magnaporthe grisea". Annual Review of Microbiology. 57: 177–202. doi:10.1146/annurev.micro.57.030502.090957. PMID 14527276.
- ↑ Zeigler, RS; Leong, SA; Teeng, PS (1994). "Rice Blast Disease." Wallingford: CAB International.
- ↑ Wilson, R. A.; Talbot, N. J. (2009). "Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae". Nature Reviews Microbiology. 7 (3): 185–95. doi:10.1038/nrmicro2032. PMID 19219052. S2CID 42684382.
- ↑ Sesma, A.; Osbourn, A. E. (2004). "The rice leaf blast pathogen undergoes developmental processes typical of root-infecting fungi". Nature. 431 (7008): 582–6. Bibcode:2004Natur.431..582S. doi:10.1038/nature02880. PMID 15457264. S2CID 549194.
- ↑ Dean, R. A.; Talbot, N. J.; Ebbole, D. J.; Farman, M. L.; Mitchell, T. K.; Orbach, M. J.; Thon, M; Kulkarni, R; Xu, J. R.; Pan, H; Read, N. D.; Lee, Y. H.; Carbone, I; Brown, D; Oh, Y. Y.; Donofrio, N; Jeong, J. S.; Soanes, D. M.; Djonovic, S; Kolomiets, E; Rehmeyer, C; Li, W; Harding, M; Kim, S; Lebrun, M. H.; Bohnert, H; Coughlan, S; Butler, J; Calvo, S; et al. (2005). "The genome sequence of the rice blast fungus Magnaporthe grisea". Nature. 434 (7036): 980–6. Bibcode:2005Natur.434..980D. doi:10.1038/nature03449. PMID 15846337.
- ↑ Couch, B. C.; Kohn, L. M. (2002). "A multilocus gene genealogy concordant with host preference indicates segregation of a new species, Magnaporthe oryzae, from M. Grisea". Mycologia. 94 (4): 683–93. doi:10.2307/3761719. JSTOR 3761719. PMID 21156541.
- ↑ Magnaporthe grisea Archived 2007-10-12 at the Wayback Machine at Crop Protection Compendium Archived 2007-07-16 at the Wayback Machine, CAB International
- ↑ Te Beest (2007). "Rice Blast". The Plant Health Instructor. American Phytopathological Society. doi:10.1094/phi-i-2007-0313-07. ISSN 1935-9411.
- ↑ Khan, Mohammad Ashik Iqbal; Rejwan Bhuiyan, Mohammad; Hossain, Mohammad Shahadat; Pratim Sen, Partha; Ara, Anjuman; Abubakar Siddique, Md; Ansar Ali, Md (2014). "Neck blast disease influences grain yield and quality traits of aromatic rice". Comptes Rendus Biologies. Elsevier Masson. 337 (11): 635–641. doi:10.1016/j.crvi.2014.08.007. ISSN 1631-0691. PMID 25444707.
- ↑ Roumen, E. C. (1992). "Partial resistance to neck blast influenced by stage of panicle development and rice genotype". Euphytica. Springer Science and Business Media LLC. 64 (3): 173–182. doi:10.1007/bf00046046. ISSN 0014-2336. S2CID 45126761.
- ↑ Titone, Patrizia; Mongiano, Gabriele; Tamborini, Luigi (2015-01-04). "Resistance to neck blast caused by Pyricularia oryzae in Italian rice cultivars". European Journal of Plant Pathology. Springer Science and Business Media LLC. 142 (1): 49–59. doi:10.1007/s10658-014-0588-1. ISSN 0929-1873. S2CID 14478689.
- 1 2 3 4 5 6 7 Yarden, O.; Ebbole, D. J.; Freeman, S.; Rodriguez, R. J.; Dickman, M. B. (2003). "Fungal Biology and Agriculture: Revisiting the Field". Molecular Plant-Microbe Interactions. American Phytopathological Society. 16 (10): 859–866. doi:10.1094/mpmi.2003.16.10.859. ISSN 0894-0282. PMID 14558687. S2CID 20430256.
- 1 2 Couch, B. C.; Fudal, I; Lebrun, M. H.; Tharreau, D; Valent, B; Van Kim, P; Nottéghem, J. L.; Kohn, L. M. (2005). "Origins of host-specific populations of the blast pathogen Magnaporthe oryzae in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice". Genetics. 170 (2): 613–30. doi:10.1534/genetics.105.041780. PMC 1450392. PMID 15802503.
- 1 2 3 4 5 6 7 8 S.C. Scardaci; et al. (2003). "Rice Blast: A New Disease in California". University of California-Davis: Agronomy Fact Sheet Series 1997-2. Archived from the original on 2006-09-11. Retrieved 2014-02-25.
- ↑ Rice Blast Archived 2015-09-24 at the Wayback Machine at University of California Integrated Pest Management
- ↑ Rice Blast Archived 2018-03-23 at the Wayback Machine at the Online Information Service for Non-Chemical Pest Management in the Tropics
- ↑ Rice Blast Archived 2010-10-20 at the Wayback Machine at Factsheets on Chemical and Biological Warfare Agents
- ↑ Sakulkoo, Wasin; Osés-Ruiz, Miriam; Oliveira Garcia, Ely; Soanes, Darren; Littlejohn, George; Hacker, Christian; Correia, Ana; Valent, Barbara; Talbot, Nicholas (23 Mar 2018). "A single fungal MAP kinase controls plant cell-to-cell invasion by the rice blast fungus". Science. 359 (6382): 1399–1403. Bibcode:2018Sci...359.1399S. doi:10.1126/science.aaq0892. PMID 29567712.
- ↑ Agrios, George N. (2005). Plant Pathology. Amsterdam: Elsevier Academic Press.
- ↑ Wilson, Richard; Talbot, Nicholas (1 Mar 2009). "Under pressure: investigating the biology of plant infection by Magnaporthe oryzae". Nature Reviews Microbiology. 7 (3): 185–189. doi:10.1038/nrmicro2032. PMID 19219052. S2CID 42684382.
- ↑ Diagnostic Methods for Rice Blast at PaDIL Plant Biosecurity Toolbox
- 1 2 Kuyek, Devlin (2000). "Implications of corporate strategies on rice research in asia". Grain. Archived from the original on 2010-07-04. Retrieved 2010-10-20.
- ↑ "Researchers in Zambia confirm: Wheat blast has made the intercontinental jump to Africa". 24 September 2020. Archived from the original on 6 October 2021. Retrieved 13 May 2022.
- ↑ Tembo, Batiseba; Mulenga, Rabson M.; Sichilima, Suwilanji; m'Siska, Kenneth K.; Mwale, Moses; Chikoti, Patrick C.; Singh, Pawan K.; He, Xinyao; Pedley, Kerry F.; Peterson, Gary L.; Singh, Ravi P.; Braun, Hans J. (2020). "Detection and characterization of fungus (Magnaporthe oryzae pathotype Triticum) causing wheat blast disease on rain-fed grown wheat (Triticum aestivum L.) in Zambia". PLoS ONE. Public Library of Science. 15 (9): e0238724. Bibcode:2020PLoSO..1538724T. doi:10.1371/journal.pone.0238724. PMC 7505438. PMID 32956369. S2CID 221843315.
- 1 2 3 4 Islam, M. Tofazzal; Croll, Daniel; Gladieux, Pierre; Soanes, Darren M.; Persoons, Antoine; Bhattacharjee, Pallab; Hossain, Md. Shaid; Gupta, Dipali Rani; Rahman, Md. Mahbubur; Mahboob, M. Golam; Cook, Nicola; Salam, Moin U.; Surovy, Musrat Zahan; Sancho, Vanessa Bueno; Maciel, João Leodato Nunes; NhaniJúnior, Antonio; Castroagudín, Vanina Lilián; Reges, Juliana T. de Assis; Ceresini, Paulo Cezar; Ravel, Sebastien; Kellner, Ronny; Fournier, Elisabeth; Tharreau, Didier; Lebrun, Marc-Henri; McDonald, Bruce A.; Stitt, Timothy; Swan, Daniel; Talbot, Nicholas J.; Saunders, Diane G. O.; Win, Joe; Kamoun, Sophien (2016-10-03). "Emergence of wheat blast in Bangladesh was caused by a South American lineage of Magnaporthe oryzae". BMC Biology. Springer Science and Business Media LLC. 14 (1): 84. doi:10.1186/s12915-016-0309-7. ISSN 1741-7007. PMC 5047043. PMID 27716181.
- 1 2 3 4 5 "New infographic highlights an early warning system for wheat blast in Bangladesh". CGIAR WHEAT. 2020-07-15. Archived from the original on 2020-12-01. Retrieved 2020-12-26.
- ↑ Kurahasi, Yoshio (1997). "Biological Activity of Carpropamid (KTU 3616): A new fungicide for rice blast disease". Journal of Pesticide Science. Archived from the original on 2022-05-16. Retrieved 2014-02-25.
- ↑ Fourmentin, Sophie; Crini, Grégorio; Lichtfouse, Eric, eds. (2018). Environmental Chemistry for a Sustainable World. Cham: Springer International Publishing. doi:10.1007/978-3-319-76162-6. ISBN 978-3-319-76161-9. ISSN 2213-7114.
- ↑ Rice Blast Archived 2010-07-31 at the Wayback Machine at Cereal Knowledge Bank
- ↑ Bhadauria, Vijai; Banniza, Sabine; Vandenberg, Albert; Selvaraj, Gopalan; Wei, Yangdou (2012-04-27). "Peroxisomal Alanine: Glyoxylate Aminotransferase AGT1 Is Indispensable for Appressorium Function of the Rice Blast Pathogen, Magnaporthe oryzae". PLOS ONE. 7 (4): e36266. doi:10.1371/journal.pone.0036266. ISSN 1932-6203. PMC 3338719. PMID 22558413. Archived from the original on 2020-08-17. Retrieved 2022-05-16.
- ↑ Bhadauria, Vijai; Banniza, Sabine; Vandenberg, Albert; Selvaraj, Gopalan; Wei, Yangdou (2012-09-01). "Alanine". Plant Signaling & Behavior. 7 (9): 1206–1208. doi:10.4161/psb.21368. PMC 3489663. PMID 22899049.
- Talbot, Nicholas J. (2003). "On the Trail of a Cereal Killer: Exploring the Biology of Magnaporthe grisea". Annual Review of Microbiology. Annual Reviews. 57 (1): 177–202. doi:10.1146/annurev.micro.57.030502.090957. ISSN 0066-4227. PMID 14527276.
- ↑ p. 184, "
APPRESSORIUM DEVELOPMENT AND FUNCTION
Once formed, the M. grisea appressorium is a dome-shaped cell with a highly differentiated cell wall structure (7, 100). The cell wall is rich in chitin and contains a layer of melanin on the inner side of the wall." - 1 2 p. 184, "Three mutants of M. grisea, albino, buff, and rosy (corresponding to the ALB1, BUF1, and RSY1 loci, respectively), have been studied extensively and are nonpathogenic. This is due to an inability to cross the plant cuticle because of the lack of melanin deposition in the appressorium."
Further reading
- California EPA. Rice Crop Infestation in Three Counties Leads To Emergency Burn Agreement Archived 2018-12-15 at the Wayback Machine, February 11, 1998
- CIMMYT. What is wheat blast? Archived 2022-03-09 at the Wayback Machine, 2019.
- Kadlec, RP. Biological Weapons for Waging Economic Warfare, Air & Space Power Chronicles
- NSF. Microbial Genome Helps Blast Devastating Rice Disease Archived 2018-12-15 at the Wayback Machine, April 21, 2005
- United States Congress. Testimony of Dr. Kenneth Alibek, 1999
- "What is wheat blast?". CIMMYT. 2019-12-11. Archived from the original on 2021-01-22. Retrieved 2020-12-21.
- "What is wheat blast? How can I manage it?" (PDF). CIMMYT. Archived (PDF) from the original on 2022-01-19. Retrieved 2022-05-13.
- 加藤 Katō, 肇 Hajime; 山口 Yamaguchi, 富夫 Tomio; 西原 Nishihara, 夏樹 Natsuki (1976). "The perfect state of Pyricularia oryzae Cav. in culture". Japanese Journal of Phytopathology. The Phytopathological Society of Japan. 42 (4): 507–510. doi:10.3186/jjphytopath.42.507. ISSN 1882-0484.
- "Blast (node and neck)". Rice Knowledge Bank. IRRI (International Rice Research Institute). 2017-08-15. Archived from the original on 2020-07-15. Retrieved 2021-03-04.
External links
- GROMO - Genomic Resources of Magnaporthe oryzae
- Magnaporthe grisea Genome Archived 2006-09-19 at the Wayback Machine
- The official Website of the International Rice Blast Genome Consortium
- Index Fungorum Archived 2008-05-11 at the Wayback Machine
- Magnaporthe grisea at MetaPathogen: stages, tissues, mating types, strains, references