بنية وجينوم فيروس الإيدز
جينوم وبروتينات فيروس الإيدز هي محل دراسة مكثفة منذ اكتشاف الفيروس عام 1983.[1][2] اعتُقد خلال البحث عن العامل المسبب لمرض الإيدز أن الفيروس سلالة من فيروس لمفاويات تي البشري[arabic-abajed 1] الذي كان يُعرف في ذلك الوقت بأنه يؤثر على الجهاز المناعي البشري ويسبب بعض سرطانات الدم. لكن قام باحثون في معهد باستور في باريس بعزل فيروس راجع غير معروف مسبقا ومختلف جينيا من المرضى المصابين بالإيدز والذي سُمي لاحقا بفيروس عوز المناعة البشري.[3]
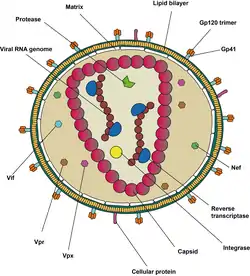
يتكون كل جسيم فيروسي (فيرون) من غلاف فيروسي مرتبط بمطرس يحيط كلاهما بقفيصة والتي تحيط بدورها بنسختي جينوم رنا مفرد السلسلة موجب الاتجاه والعديد من الإنزيمات. جاء اكتشاف الفيروس بعد عامين من الإبلاغ عن أول حالات تفشٍ كبيرة للأمراض المرتبطة بالإيدز.[4][5]
البنية
تم تحديد التسلسل الكامل لجينوم فيروس الإيدز-1 المستخلص من جسيمات الفيرون المعدية بدقة النوكليوتيد المفرد (أي معرفة جميع النوكليوتيدات).[6] يُشفر فيروس الإيدز مجموعة صغيرة من البروتينات الفيروسية، ويقوم بتكوين ارتباطات تعاون ثابتة بين بروتينات الفيروس وبين الفيروس وبروتينات المضيف، وهذا ليغزو الخلايا المضيفة ويستولي على آلياتها الداخلية.[7]
فيروس الإيدز مختلف بنيويا عن الفيروسات الراجعة الأخرى، وقطره حوالي 100 نانومتر. يتكون داخليا من قفيصة على شكل مخروط تتضمن نسختي جينوم رنا مفرد السلسلة موجب الاتجاه، إنزيمات: الناسخ العكسي، الدامج، البروتياز، وبعض البروتينات الصغيرة والبروتينات البنيوية الكبيرة.[8] يُشفر جينوم فيروس عوز المناعة البشري 8 بروتينات فيروسية تلعب أدوارا أساسية أثناء دورة تضاعف الفيرس.[7]

يتكون فيروس الإيدز-1 من نسختي جينوم رنا مفرد السلسلة موجب الاتجاه غير مرتبطتين تساهميا وغير موصولتين، تحيط بهما قفيصة مخروطية مكونة من البروتين الفيروسي P24[arabic-abajed 2] الشائع في قفيصات الفيروسات البطيئة.[9][10] طول سلسلة الرنا هو 9749 نوكليوتيد[11][12] وتملك قبعة 5' وذيل عديد الأدينين 3' والعديد من إطارت القراءة المفتوحة.[13] تُشفَّر البروتينات البنيوية الفيروسية بواسطة إطارات القراءة المفتوحة الطويلة، في حين تشفر الإطارات القصيرة البروتينات المنظمة لدورة حياة الفيروس: الارتباط، اندماج الأغشية، التضاعف، والتجميع.[13]

سلسلة الرنا المفردة مرتبطة بإحكام مع بروتينات P7 الخاصة بالقفيصة المنواة، وبروتين التجميع اللاحق P6، والإنزيمات الضرورية لتطور الفيروس مثل: الناسخ العكسي والدامج. الرنا الناقل لليسين هو مشرع الناسخ العكسي المعتمد على المغنيزيوم.[9] ترتبط القفيصة المنواة مع جينوم الرنا (جزيء لكل سداسي القسمات) وتحمي الرنا من التفكك بواسطة النوكليازات. يوجد كذلك داخل الفيروس بروتينات: Vif[arabic-abajed 3] وVpr[arabic-abajed 4] وNef[arabic-abajed 5] والبروتياز الفيروسي. يحيط مطرس مكون من البروتين الفيروسي P17 بالقفيصة، ويعمل على ضمان سلامة الجسيم الفيروسي. المطرس بدوره محاط بغلاف أصله من الخلية المضيفة، يتشكل الغلاف حين تتبرعم القفيصة من الخلية المضيفة آخذة جزءا من غشاء الخلية معها. يحتوي الغلاف على البروتينات السكرية gp120[arabic-abajed 6] والبروتين السكري gp41 المسؤولان على الارتباط بالخلايا المضيفة والدخول إليها.
لكونهما البروتينان الوحيدان على سطح الفيروس، بروتينات الغلاف السكرية (gp120 وgp41) هي هدف رئيسي لجهود تصنيع لقاح لفيروس الإيدز.[14] أزيد من نصف كتلة حسكة الغلاف ثلاثية القسمات هي غليكانات مرتبطة بالنيتروجين، وكثافتها عالية لأن الغليكانات تقي البروتينات الفيروسية الموجودة أسفلها من التحييد بواسطة الأجسام المضادة. حسكات فيروس الإيدز أحد أكثر الجزيئات المغلكزة المعروفة والكثافة مرتفعة بشكل كافٍ لمنع عملية النضج الطبيعي للغليكانات أثناء التخليق الحيوي في الشبكة الإندوبلازمية وجهاز غولجي.[15][16] ونتيجة ذلك تبقى معظم الغليكانات غير ناضجة «عالية المانوز» لا يتم إفرازها ولا تتواجد عادة في البروتينات السكرية البشرية على سطح الخلية.[17] تعني هذه المعالجة غير العادية والكثافة العالية أن جميع الأجسام المضادة المحيِّدة والتي تم تحديدها لحد الآن (من مجموعة فرعية من المرضى المصابين منذ شهور إلى أعوام) ترتبط بغليكانات الغلاف هذه أو تتأقلم للتكيف معها.[18]
تم تحديد البنية الجزيئية للحسكة الفيروسية بواسطة علم البلورات السيني[19] والمجهر الإلكتروني فائق البرودة.[20] هذه التقدمات في علم الأحياء البنيوي كانت ممكنة بفضل تطوير هيئات مؤشبة مستقرة من الحسكة الفيروسية عبر إدراج رابطة ثنائي كبريتيد بين الوحدات الفرعية وتطفير باستبدال إيزولوسين مكان برولين في البروتين gp41.[21] لا تعيد الحسكة ثلاثية القسمات التي تسمى SOSIP إنتاج الخصائص المستضدية الخاصة بالحسكة الطبيعية وحسب، بل تُظهر كذلك نفس درجة عدم نضوج الغليكانات كما هو الحال في لدى الفيروس الطبيعي.[22] الحسكات الفيروسية المؤشبة ثلاثية القسمات هي مرشحات لقاح واعد لأنها تُظهر حاتمات غير مسبِّبة للتحييد أقل من البروتين السكري المأشوب أحادي القسمات gp120، الذي يعمل على كبت استهداف الجهاز المناعي للحاتمات.[23]
تنظيم الجينوم

لفيروس الإيدز عدة جينات رئيسية تشفر بروتينات بنيوية متواجدة لدى جميع الفيروسات الراجعة، وكذلك العديد من الجينات غير البنيوية (الملحقة) الخاصة الفريدة لديه.[24] يحتوي جينوم فيروس الإيدز على تسع جينات تشفِّر 15 بروتينا فيروسيا.[25] تُخلق هذه البروتينات كعديدات بروتين ثم تُعالج لتنتج بروتينات أو محتوى الفيرون الداخلي وهي: Gag، الإنزيمات الفيروسية Pol، والبروتينات السكرية الخاصة بالفيرون env .[26] زيادة على ذلك، يشفر فيروس الإيدز بروتينات لها بعض الوظائف التنظيمية والوظائف المساعدة كذلك.[26] يملك فيروس الإيدز-1 عنصرين تنظيميين مهمين هما: Tat[arabic-abajed 7] وRev[arabic-abajed 8] وبعض البروتينات الملحقة المهمة مثل: Nef وVpr وVif وVpu[arabic-abajed 9] التي هي غير أساسية للتضاعف في بعض الأنسجة.[26] يوفر جين Gag البنية التحتية الهيكلية للفيروس، ويوفر pol الآلية الأساسية التي يتكاثر ويتضاعف بها، بينما تساعد الجينات الأخرى الفيروس على الدخول إلى الخلية المستضيفة وتحسن من تكاثره. رغم أن هذه الجينات قد تتغير بالتطفر، إلا أن جميعها باستثناء tev تتواجد لدى جميع سلالات فيروس الإيدز، انظر التباين الجيني لفيروس الإيدز.
يستخدم فيروس الإيدز نظام توصيل رنا تفاضليا متطورا لكي يحصل على تسع نواتج جينية مختلفة من جينوم يقل عن 10 آلاف كليو قاعدة.[27] لفيروس الإيدز نسخة جينومية غير موصولة طولها 9.2 كيلو قاعدة تُشفر طلائع gag وpol، نسخة موصولة مرة واحدة طولها 4.5 كيلو قاعدة تُشفر بروتينات env وVif وVpr وVpu، ونسخة رنا رسول موصولة عدة مرات طولها 2 كيلو قاعدة تشفر بروتينات: Tat وRev و Nef.[27]
بروتينات فيروسية بنيوية
- gag:[arabic-abajed 10] (مستضد مختص بمجموعة) يُشفر طليعة عديد البروتين gag الذي يُعالج بواسطة بروتياز فيروسي أثناء النمو لينتج عنه: MA[arabic-abajed 11] (بروتين المطرس، P17)، CA[arabic-abajed 12] (بروتين القفيصة، P24)، SP1[arabic-abajed 13] (الببتيد الفاصل 1، p2)، NC[arabic-abajed 14] (بروتين القفيصة المنواة، p7)، SP2 (الببتيد الفاصل 2، p1) وبروتين P6.[28]
- بي أوة ال:[arabic-abajed 15] ويُشفر الإنزيمات الفيروسية: الناسخ العكسي، الرناز H، الدامج وبروتياز فيروس الإيدز.[26] بروتياز فيروس الإيدز ضروري لقص عديد البروتين Gag لإنتاج البروتينات البنيوية، الناسخ العكسي ضروري لنسخ الدنا من سلسلة الرنا القالب، والدامج ضروري لدمج سلسلة الدنا المزدوجة الفيروسية في جينوم المضيف.[24]
- إنف:[arabic-abajed 16] يشفر عديد البروتين gp160 الذي يُقص بواسطة بروتياز الخلية المضيفة فيورين داخل الشبكة الإندوبلازمية. يُنتِج تعديل ما بعد الترجمة بروتين سكري سطحي gp120 أو SU[arabic-abajed 17] الذي يرتبط بمستقبلات كتلة التمايز 4 المتواجدة في اللمفاويات، وبروتين عبر غشائي gp41 أو TM[arabic-abajed 18] والذي ينغرس في الغلاف الفيروسي ليمكن الفيروس من الارتباط بالخلايا المستهدفة ودمج الغشائين معا.[24][28]
عناصر تنظيمية ضرورية
- tat: (منشط النسخ-مفروق) يلعب دورا مهما في تنظيم النسخ العكسي لجينوم الرنا الفيروسي، ويحرص على كفاءة تخليق جزيئات الرنا الرسول الفيروسية وتحرير الفيرونات الجديدة من الخلايا المصابة.[26] يُعبر عن Tat كبروتين يتكون من 72 حمض أميني بإكسون واحد وكذلك كبروتين يتكون من 101 بإكسونين (86–101)، ويلعب دورا مهما في الإصابة الأولية بالفيروس. وزن Tat الجزيئي هو 14-15 كيلو دالتون ويرتبط ببنية حلقة جذعية ثانوية رنوية ناتئة بالقرب من منطقة التكرار الطرفي الطويل 5' مشكلا عنصر الاستجابة للتنشيط-مفروق (TAR).[9][26]
- rev: (منظم التعبير عن بروتينات الفيرون)، يرتبط Rev بالجينوم الفيروسي عبر نمط ارتباط بالرنا غني بالأرجنين يعمل كذلك كإشارة توضيع نووي (NLS) ضروري لنقل Rev إلى النواة من العصارة الخلوية أثناء التضاعف الفيروسي.[26] يتعرف Rev على بنية الحلقة الجذعية الخاصة بالرنا الرسول لـenv المتواجدة في الإنترون الفاصل بين الإكسون المشفِّر لـTat وRev المعروف بعنصر الاستجابة لـRev [الإنجليزية] .[9][26] Rev مهم لتخليق البروتينات الفيروسية الأساسية ولذلك هو أساسي للتضاعف الفيروسي.
المصطلحات والاختصارات الأجنبية
- HTLV اختصار لـHuman T-cell leukemia virus.
- حرف الـP اختصار لكلمة Protein (بروتين).
- Vif اختصار لـViral infectivity factor، عامل الإعداء الفيروسي.
- Vpr اختصار لـViral Protein R، البروتين الفيروسي ر.
- Nef اختصار لـNegative Regulatory Factor، عامل التنظيم السلبي.
- حرفي gp اختصار لكلمة glycoprotein (بروتين سكري).
- Tat اختصار لـ Trans-Activator of Transcription، منشط النسخ-مفروق.
- Rev اختصار لـregulator of expression of virion proteins، منظم التعبير عن بروتينات الفيرون.
- Vpu اختصار لـViral Protein U، البروتين الفيروسي يو.
- Gag يُقصد به group-specific antigen، مستضد مختص بمجموعة.
- MA اختصار لـmatrix، مطرس.
- CA اختصار لـcapsid، قفيصة.
- SP1 اختصار لـspacer peptide 1، الببتيد الفاصل 1.
- NC اختصار لـnucleocapsid، قفيصة منواة.
- Pol اختصار لـpolymerase، بوليميراز.
- env اختصار لـenvelope، غلاف.
- SU اختصار لـsurface، سطح.
- TM اختصار لـtransmembrane، عابر للغشاء.
المراجع
- Barré-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vézinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L (مايو 1983)، "Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS)"، Science، 220 (4599): 868–71، Bibcode:1983Sci...220..868B، doi:10.1126/science.6189183، PMID 6189183، مؤرشف من الأصل في 25 يوليو 2020.
- Gallo RC, Sarin PS, Gelmann EP, Robert-Guroff M, Richardson E, Kalyanaraman VS, Mann D, Sidhu GD, Stahl RE, Zolla-Pazner S, Leibowitch J, Popovic M (مايو 1983)، "Isolation of human T-cell leukemia virus in acquired immune deficiency syndrome (AIDS)"، Science، 220 (4599): 865–7، Bibcode:1983Sci...220..865G، doi:10.1126/science.6601823، PMID 6601823.
- Churi, C.؛ Ross, M. W. (2015)، "Hiv/Aids"، في Whelehan, P.؛ Bolin, A. (المحررون)، The international encyclopedia of human sexuality، Wiley، ISBN 9781405190060، OCLC 949701914.
- Centers for Disease Control (يونيو 1981)، "Pneumocystis pneumonia--Los Angeles"، MMWR. Morbidity and Mortality Weekly Report، 30 (21): 250–2، PMID 6265753.
- Centers for Disease Control (CDC) (يوليو 1981)، "Kaposi's sarcoma and Pneumocystis pneumonia among homosexual men--New York City and California" (PDF)، MMWR. Morbidity and Mortality Weekly Report، 30 (25): 305–8، PMID 6789108، مؤرشف من الأصل في أكتوبر 22, 2012، اطلع عليه بتاريخ سبتمبر 15, 2017.
{{استشهاد بدورية محكمة}}: صيانة CS1: رابط غير صالح (link) - Watts JM, Dang KK, Gorelick RJ, Leonard CW, Bess JW, Swanstrom R, Burch CL, Weeks KM (أغسطس 2009)، "Architecture and secondary structure of an entire HIV-1 RNA genome"، Nature، 460 (7256): 711–6، Bibcode:2009Natur.460..711W، doi:10.1038/nature08237، PMC 2724670، PMID 19661910.
- Li G, De Clercq E (سبتمبر 2016)، "HIV Genome-Wide Protein Associations: a Review of 30 Years of Research"، Microbiology and Molecular Biology Reviews، 80 (3): 679–731، doi:10.1128/MMBR.00065-15، PMC 4981665، PMID 27357278.
- Singleton, P.؛ Sainsbury, D., المحررون (2006)، "Hiv"، Dictionary of microbiology & molecular biology (ط. 3rd)، Hoboken, NJ: Wiley، ISBN 9780470035450، OCLC 71223221.
- Montagnier, Luc (1999)، "Human Immunodeficiency Viruses (Retroviridae)"، Encyclopedia of Virology (ط. 2nd)، ص. 763–774.
- Lu K, Heng X, Summers MF (يوليو 2011)، "Structural determinants and mechanism of HIV-1 genome packaging"، Journal of Molecular Biology، 410 (4): 609–33، doi:10.1016/j.jmb.2011.04.029، PMC 3139105، PMID 21762803.
- Wain-Hobson S, Sonigo P, Danos O, Cole S, Alizon M (يناير 1985)، "Nucleotide sequence of the AIDS virus, LAV"، Cell، 40 (1): 9–17، doi:10.1016/0092-8674(85)90303-4، PMID 2981635.
- Ratner L, Haseltine W, Patarca R, Livak KJ, Starcich B, Josephs SF, Doran ER, Rafalski JA, Whitehorn EA, Baumeister K (1985)، "Complete nucleotide sequence of the AIDS virus, HTLV-III"، Nature، 313 (6000): 277–84، Bibcode:1985Natur.313..277R، doi:10.1038/313277a0، PMID 2578615.
- Castelli JC, Levy A (2002)، "HIV (Human Immunodeficiency Virus)"، Encyclopedia of Cancer (ط. 2nd)، ج. 2، ص. 407–415.
- National Institute of Health (17 يونيو 1998)، "Crystal structure of key HIV protein reveals new prevention, treatment targets" (Press release)، مؤرشف من الأصل في 19 فبراير 2006، اطلع عليه بتاريخ 14 سبتمبر 2006.
- Behrens AJ, Vasiljevic S, Pritchard LK, Harvey DJ, Andev RS, Krumm SA, Struwe WB, Cupo A, Kumar A, Zitzmann N, Seabright GE, Kramer HB, Spencer DI, Royle L, Lee JH, Klasse PJ, Burton DR, Wilson IA, Ward AB, Sanders RW, Moore JP, Doores KJ, Crispin M (مارس 2016)، "Composition and Antigenic Effects of Individual Glycan Sites of a Trimeric HIV-1 Envelope Glycoprotein"، Cell Reports (باللغة الإنجليزية)، 14 (11): 2695–706، doi:10.1016/j.celrep.2016.02.058، PMC 4805854، PMID 26972002.
- Pritchard LK, Spencer DI, Royle L, Bonomelli C, Seabright GE, Behrens AJ, Kulp DW, Menis S, Krumm SA, Dunlop DC, Crispin DJ, Bowden TA, Scanlan CN, Ward AB, Schief WR, Doores KJ, Crispin M (يونيو 2015)، "Glycan clustering stabilizes the mannose patch of HIV-1 and preserves vulnerability to broadly neutralizing antibodies"، Nature Communications، 6: 7479، Bibcode:2015NatCo...6.7479P، doi:10.1038/ncomms8479، PMC 4500839، PMID 26105115.
- Pritchard LK, Harvey DJ, Bonomelli C, Crispin M, Doores KJ (سبتمبر 2015)، "Cell- and Protein-Directed Glycosylation of Native Cleaved HIV-1 Envelope"، Journal of Virology، 89 (17): 8932–44، doi:10.1128/JVI.01190-15، PMC 4524065، PMID 26085151.
- Crispin M, Doores KJ (أبريل 2015)، "Targeting host-derived glycans on enveloped viruses for antibody-based vaccine design"، Current Opinion in Virology، Viral pathogenesis • Preventive and therapeutic vaccines، 11: 63–9، doi:10.1016/j.coviro.2015.02.002، PMC 4827424، PMID 25747313.
- Julien JP, Cupo A, Sok D, Stanfield RL, Lyumkis D, Deller MC, Klasse PJ, Burton DR, Sanders RW, Moore JP, Ward AB, Wilson IA (ديسمبر 2013)، "Crystal structure of a soluble cleaved HIV-1 envelope trimer"، Science، 342 (6165): 1477–83، Bibcode:2013Sci...342.1477J، doi:10.1126/science.1245625، PMC 3886632، PMID 24179159.
- Lyumkis D, Julien JP, de Val N, Cupo A, Potter CS, Klasse PJ, Burton DR, Sanders RW, Moore JP, Carragher B, Wilson IA, Ward AB (ديسمبر 2013)، "Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer"، Science، 342 (6165): 1484–90، Bibcode:2013Sci...342.1484L، doi:10.1126/science.1245627، PMC 3954647، PMID 24179160.
- Sanders RW, Derking R, Cupo A, Julien JP, Yasmeen A, de Val N, Kim HJ, Blattner C, de la Peña AT, Korzun J, Golabek M, de Los Reyes K, Ketas TJ, van Gils MJ, King CR, Wilson IA, Ward AB, Klasse PJ, Moore JP (سبتمبر 2013)، "A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies"، PLOS Pathogens، 9 (9): e1003618، doi:10.1371/journal.ppat.1003618، PMC 3777863، PMID 24068931.
- Pritchard LK, Vasiljevic S, Ozorowski G, Seabright GE, Cupo A, Ringe R, Kim HJ, Sanders RW, Doores KJ, Burton DR, Wilson IA, Ward AB, Moore JP, Crispin M (يونيو 2015)، "Structural Constraints Determine the Glycosylation of HIV-1 Envelope Trimers"، Cell Reports، 11 (10): 1604–13، doi:10.1016/j.celrep.2015.05.017، PMC 4555872، PMID 26051934.
- de Taeye SW, Ozorowski G, Torrents de la Peña A, Guttman M, Julien JP, van den Kerkhof TL, Burger JA, Pritchard LK, Pugach P, Yasmeen A, Crampton J, Hu J, Bontjer I, Torres JL, Arendt H, DeStefano J, Koff WC, Schuitemaker H, Eggink D, Berkhout B, Dean H, LaBranche C, Crotty S, Crispin M, Montefiori DC, Klasse PJ, Lee KK, Moore JP, Wilson IA, Ward AB, Sanders RW (ديسمبر 2015)، "Immunogenicity of Stabilized HIV-1 Envelope Trimers with Reduced Exposure of Non-neutralizing Epitopes"، Cell، 163 (7): 1702–15، doi:10.1016/j.cell.2015.11.056، PMC 4732737، PMID 26687358.
- Mushahwar, Isa K. (2007)، "Human Immunodeficiency Viruses: Molecular Virology, pathogenesis, diagnosis and treatment"، Perspectives in Medical Virology، 13: 75–87، doi:10.1016/S0168-7069(06)13005-0، ISBN 9780444520739.
- Li G, Piampongsant S, Faria NR, Voet A, Pineda-Peña AC, Khouri R, Lemey P, Vandamme AM, Theys K (فبراير 2015)، "An integrated map of HIV genome-wide variation from a population perspective"، Retrovirology (باللغة الإنجليزية)، 12 (1): 18، doi:10.1186/s12977-015-0148-6، PMC 4358901، PMID 25808207.
- Votteler J, Schubert U (2008)، "Human Immunodeficiency Viruses: Molecular Biology"، Encyclopedia of Virology (ط. 3rd)، ص. 517–525.
- Feinberg Mark B, Greene Warner C (1992)، "Molecular Insights into human immunodeficiency virus type1 pathogenesis"، Current Opinion in Immunology، 4 (4): 466–474، doi:10.1016/s0952-7915(06)80041-5، PMID 1356348.
- King Steven R (1994)، "HIV: Virology and Mechanisms of disease"، Annals of Emergency Medicine، 24 (3): 443–449، doi:10.1016/s0196-0644(94)70181-4، PMID 7915889.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الفيروسات
بوابة علم الفيروسات