اقتران صبغي
تشابك الصبغيات (بالانجليزية: Synapsis) هي عملية اقتران زوج من الكروموسومات المتماثلة خلال عملية الانقسام المنصف. تسمح العملية بمطابقة الأزواج المتماثلة قبل الفصل، وعملية العبور الجيني بين بعضهم. يحدث التشابك الصبغي خلال الطور الأول من الانقسام المنصف. عندما تتشابك الكروموسومات المتماثلة، ترتبط اطرافهم مع الغلاف النووي.ثم تهاجر هذه الاجسام المركبة، بمساعدة الهيكل الخلوي الخارجي، إلا ان تتطابق الاطراف المتشابهة. بعدها تؤخذ الاجزاء المترابطة في الكروموسوم، وتربط بواسطة مركب (ر ن ا – بروتين)يسمى بالمركب المشبكي الخيطي.[1] الصبغيات الجسدية تمر بعملية التشابك خلال الانقسام المنصف، وتربط معا بواسطة المركب ذاته على طول الكروموسومات. الكروموسومات الجنسية أيضا تمر بعملية التشابك؛ مع ذلك، فإن المركب المشبكي الذي يربط الكروموسومات المتماثلة موجودٌ على الأطراف فقط في الكروموسومات الجنسية.[2]
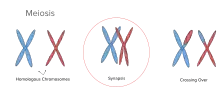
عندما تتداخل الكروماتيدات غير الشقيقة، تنكسر الأقسام الشبيهة ويتم تبادلها في عملية تسمىإعادة الاتحاد الجيني. هذا التبادل ينتج عنه تقاطع، وهي منطقة شكلها كال (X) يرتبط عندها كرموسومان.يلزم وجود تقاطع واحد على الأقل لتثبيت المثان الصبغية على طول صفيحة الطور الثاني عند الانفصال. يتيح أيضاً عبور المادة الجينية حماية ضد آليات «قتل الجينات»؛ عبر إزالة التعرف على «الذات» و «غير الذات» التي تعمل عليها تلك الآلية. من نتائج معاودة التشابك الصبغي هي زيادة التنوع الجيني في الجيل التالي، وتكراره أكثر يسبب بسماح تنقل الجينات بشكل مستقل عبر الأجبال والسماح بتركيز الجينات النافعة وطغيانها على الجينات الضارة.
يتبع التشابك الصبغي نوع من إعادة الترتيب يشار إليه ب (SDSA) Synthesis Depedent Strand Annealing)) والذي يحدث كثيراً. SDSA يتمحور حول تبادل معلومات بين الكروماتيدات المترابطة المتماثلة غير الشقيقة، لكن التبادل يكون غير مادي، SDSA لا يسبب أي عبور جيني. يعمل كلا النوعين من إعادة الترتيب (مع العبور الجيني) و (SDSA) كعمليات إصلاح لأضرار الحمض النووي، تحديداً كسور الشرائط المزدوجة.
إذاً يعتبر الدور الأساسي لعملية التشابك الصبغي هي تحديد المتماثلين بربطهم معاً، وخطوةً جذرية لعملية انقسام منصف ناجحة. تحدث عمليتا إصلاح الحمض النووي وتشكيل التقاطع بعد التشابك ولهم توابع ونتائج على مستوياتٍ عدة، من بقاء الحياة الخلوية على قيد إلى التأثيرات على التطور نفسه.
إخماد الجينات
في الثدييات، آليات المراقبة تقوم بإزالة الخلايا المنصفة التي لا يعمل فيها التشابك. وإحدى تلك الآليات هي الإخماد المنصف التي تتضمن إخماد التناسخ في الكروموسومات غير المتشابكة.[3] في الذكور أو الإناث، أي منطقة غير متشابكة على الكروموسوم تكون معرضة للإخماد المنصف.[4] (ATRAID,BRCA1 and gammaH2AX) يتموضعون على الكروموسومات غير المتشابكة خلال مرحلة تغلظ الأشرطة في الانقسام المنصف في الخلية البيضية لدى البشر وهذا قد يؤدي إلى إخماد الكروموسوم.[5] البروتين المستجيب للأضرار في الحمض النووي (TOPBP1) يعتبر عاملاً مهماً في الإسكات المنصف للكروموسومات الجنسية. هناك ظن أن كسور الشرئط المزدوجة هي مناطق ابتداء إسكات الجينات.
إعادة الترتيب/الدمج
في أنثى ذبابة الفاكهة(دروسوفيلا), يحدث التشابك المنصف للكروموسومات من دون لحاجة إلى وجود إعادة الترتيب.[6] لذا التشابك في الدروسوفيلا يكون مستقلا عن إعادة الترتيب المنصفة، مما يتماشى مع منظور أن التشابك الصبغي هو متطلب سابق لابتداء إعادة الترتيب المنصفة. وتعتبر إعادة الترتيب المنصفة غير مهمة لتشابك الكروموسومات المتماثلة عند الدودة الأسطوانية.[7]
المراجع
- Revenkova E, Jessberger R (2006)، "Shaping meiotic prophase chromosomes: cohesins and synaptonemal complex proteins" (PDF)، Chromosoma، 115 (3): 235–40، doi:10.1007/s00412-006-0060-x، PMID 16518630، مؤرشف من الأصل (PDF) في 25 مارس 2020، اطلع عليه بتاريخ أغسطس 2020.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ الوصول=(مساعدة) - Page J, de la Fuente R, Gómez R, Calvente A, Viera A, Parra M, Santos J, Berríos S, Fernández-Donoso R, Suja J, Rufas J (2006)، "Sex chromosomes, synapsis, and cohesins: a complex affair" (PDF)، Chromosoma، 115 (3): 250–9، doi:10.1007/s00412-006-0059-3، PMID 16544151، مؤرشف من الأصل (PDF) في 25 مارس 2020، اطلع عليه بتاريخ أغسطس 2020.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ الوصول=(مساعدة) - ElInati E, Russell HR, Ojarikre OA, Sangrithi M, Hirota T, de Rooij DG, McKinnon PJ, Turner JM (2017)، "DNA damage response protein TOPBP1 regulates X chromosome silencing in the mammalian germ line"، Proc. Natl. Acad. Sci. U.S.A.، 114 (47): 12536–12541، doi:10.1073/pnas.1712530114، PMC 5703310، PMID 29114052.
- Turner JM (2015)، "Meiotic Silencing in Mammals"، Annu. Rev. Genet.، 49: 395–412، doi:10.1146/annurev-genet-112414-055145، PMID 26631513.
- Garcia-Cruz R, Roig I, Robles P, Scherthan H, Garcia Caldés M (2009)، "ATR, BRCA1 and gammaH2AX localize to unsynapsed chromosomes at the pachytene stage in human oocytes"، Reprod. Biomed. Online، 18 (1): 37–44، doi:10.1016/s1472-6483(10)60422-1، PMID 19146767.
- McKim KS, Green-Marroquin BL, Sekelsky JJ, Chin G, Steinberg C, Khodosh R, Hawley RS (1998)، "Meiotic synapsis in the absence of recombination"، Science، 279 (5352): 876–8، doi:10.1126/science.279.5352.876، PMID 9452390.
- Dernburg AF, McDonald K, Moulder G, Barstead R, Dresser M, Villeneuve AM (1998)، "Meiotic recombination in C. elegans initiates by a conserved mechanism and is dispensable for homologous chromosome synapsis"، Cell، 94 (3): 387–98، doi:10.1016/s0092-8674(00)81481-6، PMID 9708740.
روابط خارجية
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي