تقزم جزيري
تقزّم جَزِيْرِيَّ أو تقزّم معزول وهو شكل من أشكال التقزّم النمطي،[1] وعملية تطوّر للحيوانات الكبيرة وحالتها،[3] يقتصر نطاق مسكنها على بيئة صغيرة، في المقام الأول بيئة الجُزُر. تختلف هذه العملية الطبيعية عن الإيجاد المُتعمّد لسُلالات قزميّة والتي تسمى التقزّم. حدثت هذه العملية عدة مرات خلال التاريخ التطوري، مع أمثلة منها الديناصورات، كالأنواع: Europasaurus و Magyarosaurus dacus ، والحيوانات الحديثة كالفيلة وأقاربها. هذه العملية، وغيرها من القطع الأثرية «لعلم الوراثة الجزرية»، ليست فقط في الجزر، لكن في حالات أخرى يكون فيها النظام البيئي معزولًا عن الموارد الخارجية والتكاثر. كما يمكن أن يشمل ذلك الكهوف والواحات الصحراوية والوديان والجبال المعزولة («جزر السماء»). يُعد التقزّم الجزيرِي أحد جوانب "تأثير الجزيرة" الأكثر عمومية أو "قاعدة فوستر" ، والتي تفترض أنه عندما تستعمر حيوانات البر الرئيسي للجُزُر، تميل الأنواع الصغيرة إلى تطوير أجسام أكبر (عملقة جزيرية)، وتميل الأنواع الكبيرة إلى تطوير أجسام أصغر. هذا في حد ذاته أحد جوانب مُتلازمة الجزيرة، الذي تصف الاختلافات في التشكل والبيئة وعلم وظائف الأعضاء والسلوك للأنواع المعزولة مُقارنة بنظيراتها القارية.

الأسباب المُحتملة
_(8609857748).jpg.webp)
هناك العديد من التفسيرات المُقترحة للآلية التي تنتج مثل هذا التقزّم.[4] [5]
الأول هو عملية انتقائية؛ حيث تعيش الحيوانات الصغيرة المُحاصرة في الجزيرة فقط، حيث ينخفض الطعام بشكل دوريّ إلى مستوى محدود. ومن المُرجّح أن تحتاج الحيوانات الأصغر حجمًا إلى موارد أقل وأراضِِ أصغر، فبالتالي يسمح انخفاض عدد السكان بتجديد مصادر الغذاء بما يفي لبقاء الناجين. والحجم الأصغر مُفيد أيضًا من وجهة نظر الإنجاب، لأنه يستلزم فترات حمل وأوقات جيل أقصر.[4]
في المناطق الاستوائية، يجعل الحجم الصغير تنظيمها الحراري أسهل. [4]
من بين الحيوانات العاشبة، يمنح الحجم الكبير مزايا في التعامل مع كل من المنافسين والحيوانات المفترسة، لذا فإن تقليل أو عدم وجود أي منهما سيُسهّل التقزّم؛ كما يبدو أن المنافسة هي العامل الأكثر أهمية.[5]
في الحيوانات آكلة اللحوم، فأنهُ يُعتقد بأن العامل الرئيسي هو حجم الفرائس وتوافرها، والمُنافسة تكون أقل أهمية.[5] في ثعابين النمر، يحدث تقزّم جزيرِي في الجُزُر التي تقطنها، حيث يُقتصر على الفريسة المتاحة أن تكون أقل حجماً، وهي أصغر مما تأخذه ثعابين البر الرئيسي. نظرًا لأن تفضيل حجم الفريسة لدى الثعابين يتناسب مع حجم أجسامهم، فقد تكون الأفاعي الصغيرة أكثر تكيفًا لأخذ الفريسة الصغيرة.[6]
التقزّم والعملقة
العملية العكسية؛ تتكاثر فيها الحيوانات الصغيرة في الجُزُر المعزولة التي تفتقر إلى حيوانات مُفترسة ذات كتل كبيرة في اليابسة، وقد يؤدي ذلك لزيادةٌ أكبر بكثير من حجمها المُعتاد، وهي ما تُسمى بالعملقة الجزيرية. وخير مثال على ذلك هو طائر الدودو، حيث كان أسلافه من الحمام ذي الحجم الطبيعي. هناك أيضًا العديد من أنواع الفئران العملاقة، التي لا تزال أحدها موجودًا، والتي تعايشت مع كل من إنسان فلوريس والستيغودون القزمي في فلوريس.
يمكن أن تحدُث عملية تقزّم جزيرِية بسرعة نسبية وفقًا لمعايير تطورية. هذا على النقيض من الزيادات في الحد الأقصى لحجم الجسم، والتي تكون أكثر تدريجيًا. بالوضع الطبيعي مع طول الجيل، وجد أن الحد الأقصى لمُعدل انخفاض كتلة الجسم لدى الثدييات أثناء التقزم الجزيرِي يزيد 30 مرة عن الحد الأقصى لمعدل زيادة كتلة الجسم لكي تتغير بعشرة أضعاف.[7] يُعتقد أن هذا التفاوت يعكس حقيقة أن تعدد الأشكال يوفر طريقًا سهلًا نسبيًا لتطوير حجم الجسم البالغ الأصغر؛ من ناحية أخرى، من المحتمل أن يتوقف تطور الحجم الأقصى للجسم الأكبر عن طريق ظهور سلسلة من القيود التي يجب التغلب عليها من خلال الابتكارات التطورية قبل أن تستمر العملية. [7]
العوامل المؤثرة في حجم التقزّم
بالنسبة لكل من الحيوانات العاشبة والحيوانات آكلة اللحوم، يبدو أن حجم الجزيرة ودرجة انعزالها وحجم الأنواع القارية الموروثة عن الأجداد ليست ذات أهمية مباشرة كبيرة لدرجة التقزم.[5] ومع ذلك، عند النظر فقط في كتل الجسم للحيوانات العاشبة والحيوانات آكلة اللحوم الحديثة، بما في ذلك البيانات كل من كتل اليابسة القارية والجزرية، وجد أن كتل الجسم لأكبر الأنواع في كتلة اليابسة تتناسب مع حجم كتلة الأرض، مع منحدرات تبلغ حوالي 0.5 لوغاريتمات (كتلة الجسم / كجم) لكل لوغاريتم (مساحة الأرض / كم 2).[8] كانت هناك خطوط انحدار مُنفصلة للحيوانات المفترسة العلوية الماصة للحرارة، والحيوانات المُفترسة العلوية خارجية الحرارة، والحيوانات العاشبة العلوية الماصة للحرارة و (على أساس بيانات محدودة) العواشب العلوية الخارجة للحرارة، مثل أن تناول الطعام كان أعلى من 7 إلى 24 ضعفًا للحيوانات العاشبة مقارنة بالحيوانات المفترسة العليا، وتقريبًا نفس الشيء بالنسبة للامتصاص الحراري وخارجية الحرارة من نفس المستوى الغذائي (وهذا يؤدي إلى أن تكون الحرارة الخارجية أثقل من 5 إلى 16 مرة من ماصات الحرارة المقابلة). [8]
أمثلة
ديناصورات غير طائرة
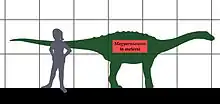
نشأ الاعتراف بأن التقزّم الجزيرِي يمكن أن ينطبق على الديناصورات من خلال عمل فيرينك نوبكسا، وهو أرستقراطي مجري المولد ومغامر وباحث وعالم حفريات. درس نوبكسا ديناصورات ترانسيلفانيا بشكل مكثف، ولاحظ أنها كانت أصغر من أبناء عمومتها في أي مكان آخر من العالم. على سبيل المثال، اكتشف الصَرَاعِيد التي يبلغ طولها ستة أمتار، وهي مجموعة من الديناصورات التي نمت في أماكن أخرى إلى 30 مترًا أو أكثر. استنتج نوبكسا أن المنطقة التي تم العثور فيها على بقايا ديناصورات كانت جزيرية، في جزيرة هاتيج (الآن حوض هاتيج أو هاتزغ في رومانيا) خلال حقبة الحياة الوسطى.[9] [10] تم قبول اقتراح نوبكسا لتقزم الديناصورات في جزيرة هاتيج على نطاق واسع اليوم بعد أن أكدت المزيد من الأبحاث أن البقايا التي تم العثور عليها ليست من الأحداث.[11]
بالإضافة إلى ذلك، تم وصف جنس بالور في البداية على أنه درومايوصوريات بحجم فيلوسيرابتور (ونتيجة لذلك هو مثال مشكوك فيه على التقزم الجزيري)، ولكن منذ ذلك الحين أعيد تصنيفه على أنه طائر ساق لا يطير بشكل ثانوي، أقرب إلى الطيور الحديثة من طائر جيهول (وبالتالي هو مثال واقع من العملقة الجزيرية).
طيور
| مثال محتمل | الاسم الثنائي / العلمي | النطاق الأصلي | حالة | قريب قاري |
|---|---|---|---|---|
.jpg.webp) |
Apteribis glenos | مولوكاي | مُنقرض (الرباعي المُتأخر) |  أبو منجل أمريكي |
| Apteribis brevis | ماوي (جزيرة) | |||
_-_Mimidae_-_bird_skin_specimen.jpeg.webp) |
Toxostoma gluttatum | كوزوميل | مُهدد الأنقراض الأقصى |  |
حيوانات
| مثال محتمل | الاسم الثنائي / العلمي | النطاق الأصلي | حالة | قريب قاري |
|---|---|---|---|---|
 كولبس أحمر زنجباري |
Piliocolobus kirkii | أنغوجا | مُهدد بالانقراض |  كولبس أحمر أوزونغواني |
 ذئب ياباني |
Canis lupus hodophilax | اليابان (باستثناء هوكايدو) | مُنقرض (1905 م) |  ذئب رمادي |
 نمر زنجباري |
Panthera pardus pardus | أنغوجا | مهدد بالانقراض الأقصى أو مُنقرض |  نمر إفريقي |
 راكون كوزوميل |
Procyon pygmaeus | كوزوميل | مهدد بالانقراض الأقصى |  راكون شائع |
 أيل فلوريدا كيز |
Odocoileus virginianus clavium | فلوريدا كيز | مُهدد بالانقراض |  أيل أبيض الذيل |
 رنة سفالبارد |
Rangifer tarandus platyrhynchus | سفالبارد | مهدد انقراض الأدنى |  رنة |
 أيل فلبيني |
Rusa marianna | الفلبين | مهدد انقراض الأدنى | _(7109798353).jpg.webp) أيل الصمبر |


أنظر أيضا
ملحوظات
مراجع
- Prothero, D. R.؛ Sereno (Winter 1982)، "Allometry and Paleoecology of Medial Miocene Dwarf Rhinoceroses from the Texas Gulf Coastal Plain"، Paleobiology، 8 (1): 16–30، doi:10.1017/S0094837300004322، JSTOR 2400564.
- Perelman؛ وآخرون (2011)، "A Molecular Phylogeny of Living Primates"، PLOS Genetics، 7 (3): 1–17، doi:10.1371/journal.pgen.1001342، PMC 3060065، PMID 21436896.
- An example of noninsular phyletic dwarfism is the evolution of the dwarfed بهيات الشعر among New World monkeys, culminating in the appearance of the smallest example, قشة قزم غربي.[2]
- Van Den Bergh, G. D.؛ Rokhus Due Awe؛ Morwood, M. J.؛ Sutikna, T.؛ Jatmiko؛ Wahyu Saptomo, E. (مايو 2008)، "The youngest Stegodon remains in Southeast Asia from the Late Pleistocene archaeological site Liang Bua, Flores, Indonesia"، Quaternary International، 182 (1): 16–48، Bibcode:2008QuInt.182...16V، doi:10.1016/j.quaint.2007.02.001.
- Raia, P.؛ Meiri, S. (أغسطس 2006)، "The island rule in large mammals: paleontology meets ecology"، Evolution، 60 (8): 1731–1742، doi:10.1111/j.0014-3820.2006.tb00516.x، PMID 17017072.
- Keogh, J. S.؛ Scott, I. A. W.؛ Hayes, C. (يناير 2005)، "Rapid and repeated origin of insular gigantism and dwarfism in Australian tiger snakes"، Evolution، 59 (1): 226–233، doi:10.1111/j.0014-3820.2005.tb00909.x، PMID 15792242.
- Evans, A. R.؛ وآخرون (30 يناير 2012)، "The maximum rate of mammal evolution"، PNAS، 109 (11): 4187–4190، Bibcode:2012PNAS..109.4187E، doi:10.1073/pnas.1120774109، PMID 22308461.
- Burness, G. P.؛ Diamond؛ Flannery (04 ديسمبر 2001)، "Dinosaurs, dragons, and dwarfs: The evolution of maximal body size"، Proceedings of the National Academy of Sciences، 98 (25): 14518–14523، Bibcode:2001PNAS...9814518B، doi:10.1073/pnas.251548698، ISSN 0027-8424، JSTOR 3057309، PMID 11724953.
- "Dwarf dinosaur island really did exist, scientists claim"، مجموعة تلغراف ميديا، 22 فبراير 2010، مؤرشف من الأصل في 25 فبراير 2010، اطلع عليه بتاريخ 26 فبراير 2010.
- Benton, M. J.؛ Csiki؛ Grigorescu؛ Redelstorff؛ Sander؛ Stein؛ Weishampel (28 يناير 2010)، "Dinosaurs and the island rule: The dwarfed dinosaurs from Haţeg Island" (PDF)، Palaeogeography, Palaeoclimatology, Palaeoecology، 293 (3–4): 438–454، Bibcode:2010PPP...293..438B، doi:10.1016/j.palaeo.2010.01.026، مؤرشف من الأصل (PDF) في 10 يوليو 2011، اطلع عليه بتاريخ 30 يوليو 2017.
- Dyke, G. (20 سبتمبر 2011)، "The Dinosaur Baron of Transylvania"، Scientific American، 305 (4): 80–83، Bibcode:2011SciAm.305c..80D، doi:10.1038/scientificamerican1011-80، PMID 22106812.
- Cuarón؛ Martínez-Morales؛ McFadden؛ Valenzuela؛ Gompper (2004)، "The status of dwarf carnivores on Cozumel Island, Mexico"، Biodiversity and Conservation، 13 (2): 317–331، CiteSeerX 10.1.1.511.2040، doi:10.1023/b:bioc.0000006501.80472.cc، S2CID 25730672.
- Wilder؛ Felger (30 سبتمبر 2010)، "Dwarf Giants, Guano, and Isolation: Vegetation and Floristic Diversity of San Pedro Mártir Island, Gulf of California, Mexico" (PDF)، Proceedings of the San Diego Society of Natural History، 42: 1–24, see pp. 9–13، مؤرشف من الأصل (PDF) في 23 نوفمبر 2021، اطلع عليه بتاريخ 05 يناير 2020،
(p. 12) The dwarfing of the San Pedro Mártir plants seems to be due to a selection for shorter individuals to survive fierce tropical storms, possible root competition in such a dense forest, and the undefined effect of high levels of nitrogen and phosphorus from the abundant guano that might stunt growth. Genetic studies have not been undertaken...
- Burns, K.C. (مايو 2019)، Evolution in Isolation: The Search for an Island Syndrome in Plants، مطبعة جامعة كامبريدج، ص. 174–177، doi:10.1017/9781108379953، ISBN 978-1108379953، OCLC 1108160200، S2CID 186536407، مؤرشف من الأصل في 29 يونيو 2022،
(pp. 174-175) ... the extent to which its dwarfed stature is genetically determined, and an explanation for why insular dwarfism might be selectively advantageous, awaits additional study.
روابط خارجية
 بوابة جزر
بوابة جزر بوابة علم الأحياء
بوابة علم الأحياء بوابة علم البيئة
بوابة علم البيئة بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري بوابة علم الحيوان
بوابة علم الحيوان