وحشيات الأرجل
وحشيات الأرجل أو الثيروبودا (الاسم العلمي: Theropoda)، (من الإغريقية:Thero = θηρίον = وحشي + poda = πούς, ποδός = قدم[1][2])، رتيبة من الديناصورات سحليات الورك ثنائية الحركة ويتفرع منها عدد هائل من فصائل الديناصورات. كانت من آكلات اللحوم بشكل سلالي، رغم من أن عددا من مجموعاتها تطورت لتصبح عاشبات، وآكلة للحوم والنبات معا، وآكلات الأسماك، وآكلات الحشرات. ظهرت وحشيات الأرجل لأول مرة في مرحلة الكارني في أواخر العصر الثلاثي قبل 231.4 مليون سنة،[3] وشملت اللواحم الأرضية الكبيرة الوحيدة منذ الجوراسي المبكر وحتى نهاية العصر الطباشيري على الأقل قبل 66 مليون سنة. خلال العصر الجوراسي، تطورت الطيور من وحشيات الأرجل الصغيرة المتخصصة السيلوروصوريات، ويمثلها اليوم حوالي 10,500 نوع حي وهي الوحيدة من وحشيات الأرجل التي نجت من انقراض العصر الطباشيري-الباليوجيني قبل 65 مليون سنة.
اضغط هنا للاطلاع على كيفية قراءة التصنيف العصر: الثلاثي المتأخر-الحاضر 231.4–0 مليون سنة | |
|---|---|
 | |
| المرتبة التصنيفية | رتيبة |
| التصنيف العلمي | |
| المملكة: | حيوان |
| الشعبة: | حبليات |
| الطائفة: | عظائيات الوجه |
| الطويئفة: | ثنائيات الأقواس |
| الرتبة العليا: | ديناصور |
| الرتبة: | سحليات الورك |
| الرتيبة: | وحشيات الأرجل |
| الاسم العلمي | |
| Theropoda أوثنييل تشارلز مارش ، 1881 | |
| تحت رتيبات | |
| |
الخصائص

احتفظت وحشيات الأرجل على بنيتها الجسمانية بصفة عامة على شكل أسلافها من الديناصورات. وبالمقارنة بينها وبين فصائل ديناصورات أخرى كأشباه الصوروبوديات أو طيريات الورك فليس لديها ألا خصائص قليلة. فقد كانت وحشيات الأرجل تمشي على القدمين الخلفيتين، وكانتا أطول وأكبر من الذراعين في جميع وحشيات الأرجل. وقد كانت الذراعين أو القدمين الأماميتين لا تستخدم في الحركة والمشي. وهذا ما يميز وقفة جسم هذه الديناصورات المائلة نوعا ما إلى الأمام مع انضمام اليدين القصيرتين أمام الصدر.
تتميز وحشيات الأرجل بالتنوع الكبير في أحجامها من بين جميع الديناصورات. فمنها الصغير مثل الميكرورابتور والبارفيكورصور التي يصل طول كل منهما إلى أقل من 1 متر وكان وزن كل منهما بضعة كيلوغرامات فقط. أما وحشيات الأرجل الكبيرة فوصل طولها نحو 15 متر، مثل التيرانوصور والكاركارودونتوصور والجيجانوتوصور والسبينوصور. تعتبر تقدير أحجام تلك الديناصورات افتراضية لأن العديد من ما وجد من هيا كلها لم تكن مكتملة. ويعتبر تقدير أوزان الحيوانات المنقرضة صعبا نوعا ما، وتقدر أوزان وحشيات الأرجل الكبيرة بين 5 و 8 طن.
الجمجمة

تمتلك وحشيات الأرجل جمجمة كبيرة نوعا ما، وبالمقارنة مع الديناصورات الأخرى، فإن تركيبة الجمجمة غير متخصص وتظهر تغييرات طفيفة فيها مقارنة بجمجمة الأركوصورات. وكما هو الحال عند الأركوصورات فإنه بالإضافة إلى الفتحتين لثنائيات الأقواس، توجد فتحة واحدة أخرى أمام محجر العينين. كما تميزت وحشيات الأرجل بوجود عظمة دمعية بارزة أعلى الجمجمة، بالإضافة إلى فتحة الفك العلوي أمام العين.كانت عيون وحشيات الأرجل كبيرة وواسعة وخاصة في السيلوروصوريات، وهي متجه نحو الأمام، كما هو الحال لدى العديد من آكلات اللحوم الأخرى. وقد مكن وضع العينين بالأمام للحيوان بالرؤية الجيدة وتقدير المسافة للفريسة. وكان تجويف الأذن الوسطى ضخم بشكل كبير. مما يجعلنا نستنتج أن لوحشيات الأرجل رؤية وسمع جيدان. ولدى وحشيات الأرجل في أسفل الجمجمة مفصل العظم القذالي والذي يشكل مع فقرة الأطلسية مفصل الرأس العلوي، وهي كبير ومستدير وبالتالي تضمن زيادة ومرونة حركة الرأس مع تركيبة عضلات الرقبة قوية. وتبين جمجمة وحشيات الأرجل وجود حركة فكية معينة لكل عنصر (الحركة القحفية) تجعلها تلتهم فريستها بشكل أسهل، لأن وحشيات الأرجل لا تستطيع أكل طعامها بالمضغ.
من صفات وحشيات الأرجل أيضا وجود بروزات عظمية تشبه القرون على الرأس، وهي تختلف كثيرا عن بعضها البعض في التركيب، فكانت هشة في بعضها كما في الدايلوفوصور أو في الأوفيرابتور. وأحيانا كانت قوية جدا كما في السيراتوصور أو الكارنوتوروس. ويبدو أن تركيبة الجمجمة قد لعبت دورا كبيرا في التفاعل مع أنواع معينة كما كانت تستخدم في الدفاع عن النفس.
الأسنان

أسنان وحشيات الأرجل مفلطحة من الجانبين ومنحنية قليلاً إلى الخلف، والغالبية منها تشبه الشفرة، وحوافها مشرشرة.[4] إن مورفولوجيا الأسنان واضحة بما يكفي لتمييز الفصائل الكبرى عن بعضها البعض،[5] والتي تشير إلى استراتيجيات النظام الغذائي المختلفة. وتختلف شكل الشرشرة على طول الحافة القاطعة بين كل مجموعة، فلدى الأنواع الأصغر عددا أقل ومدببة، بحيث تكون مناسبة للأنشطة الغرائزية، بينما في الأنواع الأكبر مثل التيرانوصوريات فإن الأسنان أكثر انحناء وأقل حدة. ومع ذلك فإن بنية الأسنان بدائية، ويمكن العثور على نفس النمط على سبيل المثال في رصغيات الساق (أسلاف التماسيح القاعدية). وفي الديناصورات القريبة منها كالصوروبودات البدائية يظهر التشرشر في أسنانها أيضا، وبالتالي فإن الشقوق بزاوية 45 درجة إلى أعلى حواف القطع أو تكون في اتجاهات مختلفة.
في يوليو 2015 تبين من أحد الاكتشافات أن "التشققات" في أسنان وحشيات الأرجل كانت في الواقع طيات كانت تساعد في منع تكسر الأسنان من خلال تقوية التسننات الفردية أثناء مهاجمتها لفريستها.[6] وقد ساعدت الطيات على بقاء الأسنان في مكانها لفترة أطول، وخاصة وأن وحشيات الأرجل تطورت إلى أحجام أكبر وكان لديها قوة أكبر في عضتها.[7][8]
يتبين من تركيبة الأسنان أن معظم وحشيات الأرجل كانت آكلات للحوم. وقد كانت تستخدم أسنانها للإمساك بفرائسها وتمزيق لحمها؛ وفي بعض الأنواع التي تمتلك أسنان غير حادة كانت تستخدمها لسحق العظام. أما التريزينوصورات العاشبة فلها أسنان صغيرة ملعقية الشكل.
كان هناك فقدان للأسنان عدة مرات خلال تاريخ تطور وحشيات الأرجل. وهذا ينطبق على الأورنيثوميموصوريات (معظم الأنواع الأصلية التي تمثلها، ومع ذلك فلا يزال لديها أسنان أمامية للفك السفلي)، وكذلك العظائيات السارقة للبيض. وقد دار الجدل حول ما تأكل وحشيات الأرجل العديمة الأسنان (أنظر النظام الغذائي). وعلى عكس الكثير من أسلافها التي عاشت خلال حقبة الحياة الوسطى فإن الطيور الحديثة بلا أسنان أيضا.
الهيكل العظمي والحوض
كما هو الحال مع جميع الديناصورات فقد كانت الأطراف الخلفية في وحشيات الأرجل بوضع عمودي تحت الجسم وكان الجسم متوازن فوق الحوض. وكان العمود الفقري بوضع أفقي تقريبا؛ للسماح بالرؤية للأمام بشكل جيد، وكانت الرقبة منثنية كما في الطيور على شكل حرف "S" -وتمثل النماذج القديمة لوحشيات الأرجل بعمود فقري مائل للأسفل والذيل يسحب على الأرض، لكن هذا الشكل أصبح غير صحيح طبقا للأبحاث الحديثة. وفي السمات المشتركة يوجد لدى وحشيات الأرجل خمس فقرات عجزية أو أكثر. وعادة ما يكون الذيل طويلًا وبعدد كبير من الفقرات الذيلية. في ديناصورات الصلبات الذيل وهي أحد أكثر مجموعتين فرعيتين من وحشيات الأرجل الغنية بالأنواع، فقد كان الذيل لديها متماسكا وأفقيا فوق الأرض.
.jpg.webp)
كانت فقرات الذيل طويلة وذات بروزات من الأمام والخلف (قوس فقري) ومن ناحية أخرى كانت عظام السبعاني (شكل حرف V على الجانب السفلي من الفقرات الذيلية) متماسكة. وفي الطيور الحديثة نمت آخر فقرات ذيلية لتشكل معا فقرة ذنبية داعمة لريش الذيل تسمى (pygostyle) وقد تم العثور على هياكل مماثلة في بعض وحشيات الأرجل الغير طيرية، مثل النومينغيا والبيبياوصور.
يتميز الهيكل العظمي لوحشيات الأرجل بأن عظام الأطراف الطويلة رقيقة الجدران ومجوفة، كما أن الفقرات مجوفة أيضا. وتبدو الفقرات في الماجونغاصور ذات حجيرات هوائية، وهذا يعني أنها مليئة بنتوءات (رُتوج) ذات نظام أكياس هوائية، وتم العثور على اكتشافات مماثلة في ديناصور الميريسيا. وهذه علامة أن وحشيات الأرجل على الأقل (وربما ديناصورات أخرى أيضًا) لديها جهاز تنفسي مشابه لنظام الطيور، مع تدفق دائم أحادي الاتجاه للهواء في ذلك الجزء من الرئتين التي يكون فيها الجزء الرئيسي لتبادل الغازات ("التدفق من خلال الرئة").
الأطراف
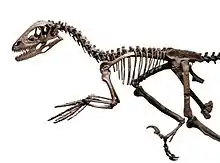
من خصائص منطقة الكتف في وحشيات الأرجل أن عظم الكتف يشبه الحزام، بالإضافة إلى أن لها عظم ترقوة شبيه بالبَمرَنغ -التي كانت تعتبر في السابق ميزة خاصة بالطيور-. وقد كان عظم الترقوة معروف عند الديناصورات القاعدية مثل السويلوفيسيات. بالنسبة إلى تركيبة ورك وحشيات الأرجل فكما هو الحال مع جميع سحليات الورك، فإن عظم العانة بارزة في الأصل للأمام. وخلال مراحل تطور وحشيات الأرجل كان هناك تعاقب، ففي الداينونيكوصورات بارزة في الأسفل وأخيرا في الطيور بارزة في الوراء. وبشكل محير فإن حوض الطيور الحديثة شبيه جدا بشكل حوض طيريات الورك المجموعة الفرعية الثانية من الديناصورات، بالرغم من أن الطيور قد تطورت من سحليات الورك وليس من طيريات الورك، والتشابه يعتبر سطحي فقط.



من السمات المشتركة لوحشيات الأرجل في تركيبة الأطراف الأمامية أن اليدين ضخمتين، حيث يصل طولها إلى 50% من طول الذراع الكامل، ومن ناحية أخرى، فإن الأصبعين الرابع والخامس قصيرين لدرجة الانعدام. تتكون اليد عادةً من أصابع الأول والثاني والثالث (يتم عدها من الإبهام)، حيث أن الأصبع الأول متحرك جزئيًا، لمقابلة الأصبع الآخر بشكل جزئي لمسك الأشياء. وفي التيرانوصوريات الإصبع الثالث قصير وفي ألفاريزصوريات مثل المونونيكس فإن ("المخلب") ليس إلا إصبع متطور. أما الأصابع المتبقية فهي طويلة نوعا ما، ولديها مجال كبير في الحركة بفضل الانحناء في الجزء العلوي من الطرف الأمامي لعظام المشط. يتميز العديد من وحشيات الأرجل، وغالبا في الصغيرة منها بأن لديها أذرع طويلة مع أيدي كبيرة وأصابع نحيلة. وقد تمكنت هذه الحيوانات بالإمساك باليد واصطياد الفريسة. (في الطيور، تطورت الذراعين بشكل خاص للتكيف مع الطيران، لذلك لديها مشط رسغي.)
في المقابل يظهر في بعض وحشيات الأرجل الكبيرة مثل الأبليصوريات والتيرانوصوريات قصرا في الأطراف الأمامية، وبالتالي لا تستطيع الوصول إلى أفواهها وبالتالي لم تكن مهمة في الصيد. لا زال هناك تسائل حول وظيفة هذه الأذرع القصيرة. وفقًا لإحدى النظريات، فإن هذه الميزة مرتبطة من ناحية الميكانيكا الحيوية: للحفاظ على التوازن بسبب ضخامة الرأس وثقله، قلت احجم الذراعين. واللافت أن هذه الأذرع القصيرة كانت قوية جدًا وعضلية. وربما تخدمها لتتمكن من الوقوف مرة أخرى خلال وضعية النوم المفترضة (الاستلقاء على البطن). وتقول نظريات أخرى أن وظيفتها هو تثبيت الشريك أثناء التزاوج.
في وحشيات الأرجل دائما تكون الأطراف الخلفية أطول من الأطراف الأمامية. وعادة يكون الجزء السفلي من الساق أطول من الفخذ (خاصة في الترودونتيدات والأورنيثوميموصوريات)، مما يوحي إلى أن هذه الحيوانات قد تصل إلى سرعات عالية. عند اصطدام اقدمها بالأرض تصبح ضيقة ويتمدد مشط القدم - وهذا ما تتميز بها طيريات الرقبة (الديناصورات والتيروصورات) التي تتميز بأن كاحلها في منتصف الرصغ. ويكون أصبع القدم الثاني والثالث والرابع متجه للأمام بشكل متماثل وتتميز بعدد من السلاميات (صيغتها 3 و 4 و 5). وهذه الأصابع هي أصابع للمشي لأن الأصبع الداخلي والأول فقط هي القصيرة ولا تلمس الأرض، وأما الخارجية والخامس فيكون العظم فيها صغير جدا. لذلك كانت المهام في أقدام وحشيات الأرجل النموذجية في الثلاث أصابع. يشير إصبع القدم الأول في الطيور الحديثة عادةً إلى الخلف (متباينة الأصابع)، ومع ذلك، فقد تطور ترتيب أصابع القدم الأخرى في هذه المجموعة.
تنتهي أصابع القدم بمخالب حادة، والتي تناقص حجمها لتصبح مدببة تكون منحنية. وقد تطورت بشكل خاص في مجموعة الداينونيكوصورات. يتكون إصبع القدم الثاني من مخلب كبير وحاد بشكل خاص وله مفصل خاص كذلك، وهذا يجعل من الممكن رفع إصبع القدم عند الحركة، ولتجنب التلف، وزيادة من الرشاقة عند الصيد. كما أن طيريات لأرجل ذات القدمين بالثلاثة أصابع من مجموعة ديناصورات طيريات الورك تفتقر إلى المخالب الطويلة، وهو أمر مهم لآثار الأقدام الأحفورية.
يعتقد أن هناك اختلاف في وظيفة الأطراف الأمامية لمختلف فصائل ن وحشيات الأرجل. وربما كان السبينوصور يستخدم أطرافه الأمامية في الإمساك بالأسماك. ويعتقد أن بعض وحشيات الأرجل الصغيرة مثل السيلوروصوريات والدرومايوصوريات كانت تستخدم أطرافها الأمامية للتسلق على الأشجار.
كانت حرية حركة الأطراف الأمامية لوحشيات الأرجل محدودة للغاية،[9] ويرجح أن حركة الذراعين كانت مشابهة لحركة جناح الطائر. في الكارنوصوريات مثل الأكروكونثوصور تكون اليد واسعة الحركة وكذلك الأصابع قابلة للحركة. وكان ذلك أيضا هو صفة وحشيات الأرجل القاعدية مثل الهيريراصوريات والدايلوفوصوريات. وتبدي السيلوروصوريات وظيفة إمكانية ومرونة لها عند الكتف مما يجعل حركة الذراع إلى أعلى ممكنة، كما أن حركتها أكبر في الطيور عند الطيران. ويبدو أن الذراعين تفقد معظم حرية حركتها في السيلوروصوريات مثل الأورنيثوميموصوريات وبصفة خاصة بالنسبة إلى الدرومايوصوريات، بينما تزداد حرية حركة الأصابع. كما تبدي الدرومايوصوريات والمانيرابتورات حركة حرة عالية لمفصل اليد غير موجود في وحشيات الأرجل الأخرى، أذ أن شكل المفصل في الدرومايوصوريات نصف دائري مما يتيح لليد للحركة أيضا إلى الخلف بطريقة حركة جناح الطيور في وقتنا الحاضر.[9]
الجلد والريش والتنظيم الحراري

ربما كان التغير الجذري في النظريات حول ظهور وحشيات الأرجل كان يتعلق بالجلد أو الحراشف. عندما كانت وحشيات الأرجل التي من حقبة الحياة الوسطى متنوعة للغاية من حيث نسيج الجلد وتغطيته. كانت معظم سلالات وحشيات الأرجل يظهر عليها الريش أو شبه الريش. (انظر ديناصور ذو ريش). ومع ذلك، قد يكون الريش محصوراً في الصغار، والأنواع الأصغر، أو أجزاء محدودة من الحيوانات التي خارج رتيبة السيلوروصوريات. وكانت وحشيات الأرجل الأكبر حجما مغطاة بجلد ذات حراشف صغيرة مدببة. وفي بعض الأنواع، كانت تتخللها حراشف أكبر وعظمية، أو جلد عظمي، يشتهر سيراتوصور الكارنوتوروس بهذا النوع من الجلد، الذي حفظ بانطباعات جلدية واسعة.[10]
كانت سلالات السيلوروصوريات هي الأبعد عن الطيور حيث أن لها ريشا قصيرا نوعا ما ويتكون من شعيرات متفرعة بسيطة.[11] تظهر الشعيرات البسيطة أيضًا في االتريزينوصوريات، التي تمتلك أيضًا ريش صلب كبير. أكثر وحشيات الأرجل المغطاة بالريش بالكامل، مثل الدرومايوصوريات، وكانت الحراشف تغطي سيقانها فقط. وقد يكون لبعض الأنواع ريش مختلط في أماكن أخرى من الجسم أيضًا. تمتلك السانسوريأوبتريكس حراشف بالقرب من الجانب السفلي من الذيل،[12] وتمتلك الجورافيناتور [الإنجليزية] حراشف تتخللها شعيرات بسيطة.[13] ومن ناحية أخرى، كانت بعض وحشيات الأرجل مغطاة بالكامل بالريش، مثل ترودونتيد الأنكيورنيس، التي كان الريش يغطي حتى القدمين والأصابع.[14]
في عام 1996 تم اكتشاف علامات الريش لأول مرة في ديناصور السينوساروبيتركس أحد أنواع وحشيات الأرجل الغير طيرية. وفي السنوات التالية ثم اكتشاف عدة ديناصورات ذات ريش، وقد وصل عدد الأنواع التي يغطيها الريشي نحو 15 نوعا. وتصنف أنواع الريش إلى عدة أنواع ما بين ريش بسيط شبه شعري إلى الريش الذي المعروف لدى أجنحة الطيور المعاصرة. (نادرًا ما يتحجر الريش بشكله الناعم، فقد اقتصرت هذه الاكتشافات حتى الآن على مواقع قليلة).
لا يظهر على معظم وحشيات الأرجل ذات الريش القدرة على الطيران بسب البنية الجسمانية، ويعتقد أن وظيفة الريش هو المحافظة عل درجة حرارة الجسم. وربما كانت هناك وظائف أخرى للريش مثل التمويه والاختفاء أو إرسال إشارة معينة للاتصال بالقرائن من نفس النوع.

طبقا لنظريات العالمين "ريشارد بروم" و"آلان بروش" أن الريش تطور على 5 مراحل.[15] (أنظر نظرية تطور الريش):
- المرحلة الأولى تتكون قصبات أسطوانية رفيعة مجوفة.
- المرحلة الثانية تتكون خصلات شعر مقنزعة تشبه ريش الطيور الموجودة اليوم.
- المرحلة الثالثة تطور نوعين من الريش: الأولى وهي الريشة كما نعرفها أيامنا هذه، ذات قصبة ثنائية الفلقة ولكن دون تشابك الشعيرات الريشية في الفلقتين. الثانية تتكون من قصبة عليها عدة خصل من الشعر، ونجدها في الطيور المعاصرة،
- المرحلة الرابعة تطور الريش إلى ما يشبه الريش المعروفة الآن، وتكون متناظرة.
- المرحلة الخامسة تطورت الريش إلى الريش المعروفة الآن، ذات الفلقتين غير متناظرتين وشعيرات كل كلنهما متشابكة.
تشير البحوث الإضافية إلى التطابق المفترض بين تطور الريش مع التطور العام لوحشيات الأرجل. فمثلا كانت السيلوروصوريات مغطاة بالريش، وهذا ينطبق على ديناصورات السينوساروبيتركس (التي تصنف على أنها أنيقات الفك وبالتالي على أنها سيلوروصوريات قاعدية) وكان لها ريش من النوع 1 أو 2. وحتى مع الديلونغ أحد التيرانوصورويدات الأولية تم العثور على علامات تشير إلى هذا الريش البسيط. وفي تريزينوصور البيبياوصور وجد أن لديه ريش من النوع 2 أو 3، بينما للأوفيرابتوروصوريا الكوديبتركس كان الريش لديها من النوع 4. وفي الداينونيكوصورات ويمثلها الميكرورابتور فيبدو أن كان له ريش غير متناظر الفلقتين من النوع 5، وقد تم اكتشاف هذا هو النوع من الريش على الطائر القديم الأركيوبتركس.
السؤال الذي حير العلماء هو هل الديناصورات من ذوات الدم الحار أم البارد (انظر ذوات الدم الحار للديناصور). يعتقد العلماء منذ اكتشاف الديناصورات ذات الريش إنه كانت للسيلوروصوريات ذات الريش خاصية تنظيم درجة حرارة الداخلية وأن بعض وحشيات الأرجل ذات الريش المتطور لديها تنظيم فعال لدرجة حرارة جسمها كما في الطيور في العصر الحاضر. كما يعتقد بعض الباحثون أن أنواعا أخرى من وحشيات الأرجل لها خاصية تنظيم درجة الحرارة بطرق أخرى مختلفة، على سبيل المثال لدى السبينوصور أشواك في الظهر شائكة وممتدة، ولدى الأكروكونثوصور فقرات شراعية الشكل، قد تكون لهذا الغرض. ومع ذلك فإن العلماء لم يتفقوا بعد على ذلك.
يعتقد بعض العلماء أن السيلوروصوريات الكبيرة لم يكن لها ريش أو شعر كما لدى التيرانوصور.[16] غالبا في الحيوانات الكبيرة، تكون نسبة مساحة السطح بالحجم أصغر مما هي عليه في الحيوانات الأصغر. وبما أن الحرارة تتسرب عبر سطح الجلد، فإنه من المرجح أن ترتفع درجة حرارة الحيوانات الكبيرة غذا كان عليها ريش أو شعر، لأن يعيق انبعاث الحرارة. (لهذا الأسباب، فإن أكبر الثدييات، كالفيل ووحيد القرن، لا يوجد لديها شعر). وبملاحظة التخطيط النمطي لريش السيلوروصوريات، باستثناء التيرانوصوريات، لا يمكن أن يوجد بأشكال عملاقة.
لا يعرف بالضبط كيف كانت تغطية الجلد لدى وحشيات الأرجل الغير سيلوروصوريات. وتوجد مؤشرات في الكارنوتوروس من فصيلة الأبليصوريات تبين أن جلده كان مغطى بالحراشف.
الحجم

منذ عقود عديدة كان التيرانوصور أكبر وحشيات الأرجل المعروف وأكثرها شهرة لعامة الناس. ومع ذلك، فقد تم وصف عدد من الديناصورات العملاقة الأخرى الآكلة للحوم منذ اكتشافها، منها السبينوصور، والكاركارودونتوصور، والجيجانوتوصور.[17] تدعم عينات السبينوصور الأصلية (بالإضافة إلى الحفريات الأحدث الموصوفة في عام 2006) فكرة أن السبينوصور أطول من تيرانوصور، وتبين أن السبينوصور ربما كان أطول بثلاثة أمتار من التيرانوصور بالرغم من أن التيرانوصور لا يزال أعظم من السبينوصور.[18] وتبين عينات التيرانوصور بأنه أكثر وحشيات الأرجل وحشية وقوة معروف للعلماء. لا يوجد تفسير واضح حتى الآن لسبب نمو هذه الحيوانات بشكل أكبر بكثير من الحيوانات المفترسة التي جاءت قبلها وبعدها.
أكبر وحشيات الأرجل الموجودة حاليا هي النعامة الشائع، حيث يصل ارتفاعها إلى 2.74 م وتزن ما بين 60 و 145 كجم. وأصغر وحشيات الأرجل الغير طيرية المعروفة من العينات البالغة هي ترودونتيد الأنكيورنيس، يصل طوله إلى 34 سم ويزن 110 جرام.[14] وعندما يتم ادراج الطيور الحديثة، فإن طائر النحلة الطنان يعتبر أصغر حجما حيث يصل طوله إلى 5.5 سم ووزنه 1.9 جم.[19] تقول بعض النظريات الحديثة أن حجم أحسام وحشيات الأرجل قد تقلص بشكل مستمر على مدى 50 مليون سنة، من 163 كجم إلى 0.8 كجم، حتى تطورت في النهاية إلى طيور حديثة. وهذا دليل على أن وحشيات الأرجل هي الديناصورات الوحيدة التي كانت تصغر باستمرار، وأن هياكلها العظمية قد تغيرت أسرع أربع مرات من الأنواع الأخرى من الديناصورات.[20][21]
البيولوجيا القديمة
الانتشار والموائل
كانت وحشيات الأرجل منتشرة في جميع أنحاء العالم، وقد وجدت جميع أحافيرها المعروفة من جميع القارات -حتى أن الكرايولوفوصور أيضًا وجد في القارة القطبية الجنوبية. أرتبط الانتشار العالمي المبكر خلال تطور وحشيات الأرجل كانت جميع القارات لا تزال متصلة في قارة بانجيا العظمى. ومنذ تفكك هذه القارة، تطورت إلى وحشيات أرجل مختلفة على أراضي فردية. عرفت الأبليصوريات بشكل حصري من القارة الجنوبية السابقة من غندوانا، ومجموعات أخرى مثل التيرانوصورويدات، عرفت فقط من شمال القارة لوراسيا. بسبب ندرة اكتشافات الحفريات بشكل عام في نصف الكرة الجنوبي، فلا يمكن إعطاء معلومات دقيقة عن مناطق توزيع العديد من أصناف وحشيات الأرجل.
هناك أيضًا مناطق كبيرة فيها ترسبات لوحشيات الأرجل. بجانب مجاري الأنهار، وفي السهول الفيضية وبقرب البحيرات، واكتشفت بعض الأماكن الجافة القريبة من الصحاري. ولا يعرف ما تفضله وحشيات الأرجل من المواطن.
السلوك الاجتماعي
التكهنات حول السلوك الاجتماعي لوحشيات الأرجل صعب كما هو الحال مع جميع الحيوانات التي عرفت من الاكتشافات الأحفورية فقط، حتى أن اكتشاف بقايا حيوانات عديدة في مكان واحد لا يشير إلى حياة المجموعة، ولكن يمكن أن يكون بسبب تقنية الترسيب، ومعظم الاكتشافات تأتي من حيوانات فردية. ومع ذلك، فهناك نوعان تم اكتشاف جماعات منها بالمئات: السيلوفايسس في غوست رانش (نيومكسيكو) والألوصور في محجر كليفلاند لويد (يوتا). وقد أثارت هذه الاكتشافات الجماعية الجدل حول تفسيرها، يعتقد أنها تجمعات للصيد وأيضا أحدد الاحتمالات أنها كانت بالقرب من مصادر المياه، بحيث أن الحيوانات تجمعت ونفقت في نهاية المطاف من العطش؛ ويعتقد أنه كان فخا جذب الحيوانات ولم يعد بإمكانها تحرير نفسها منه. وقد وجد أيضا عدة ديناصورات من الدينونيكوس، بين الأخرى ويعتقد أن له صلة بالفريسة المحتملة للتينونتوصور، والذي يعتبر مؤشرا على الصيد الجماعي.[22]

ربما لعبت النتوءات والبروزات في أعلى الجمجمة دورا في التفاعل مع القرين. في حين أن القمم الهشة للعديد من الحيوانات قد لا يكون لها أي غرض آخر، سوى الاستعراض، والتي لها قرون قوية كانت تستخدم للقتال المباشر. لا تزال التكهنات فيما يتعلق بالتزاوج، أو التسلسل الهرمي للمجموعة.
تظهر على بعض الثيروبودات علامات ازدواج الشكل الجنسي. على سبيل المثال، هناك شكلان من بعض السيلوفايسيات، أحدها برأس ورقبة طويلة وأطراف قوية، وأخرى برأس قصير وأطراف أنحف. قد يكون هناك أختلافا جسديا بين الذكور والإناث. وقد تكون هناك أيضا اختلافات مماثلة بين الجنسين في التيرانوصور، يتم تحديد الشكل الأكثر قوة بأنها أنثى بسبب الحوض الواسع.[23]
الوقوف والمشي
.jpg.webp)

من المحتمل أن وحشيات الأرجل تبنت طريقة الوقوف أو الانتصاب لسلالات متنوعة جدا بين مختلف الحيوانات عبر الزمن.[24] تعتبر جميع وحشيات الأرجل المعروفة ثنائيات الحركة، مع قوائم أمامية قصيرة مخصصة لمهام متنوعة (انظر أدناه). في الطيور الحديثة، عادة ما يكون وضع جسمها منصبا إلى حد ما، بحيث يكون الجزء العلوي من الساق (عظم الفخذ) موازيا للعمود الفقري وبقوة حركة أمامية ناتجة من الركبة. ولا يزال العلماء غير متأكدين إلى أي مدى يمتد هذا النوع من الوقوف والحركة في شجرة فصائل وحشيات الأرجل.[24]
تم التعرف على وحشيات الأرجل الغير طيرية لأول مرة بأنها ثنائيات الحركة خلال القرن 19، قبل أن يتم قبول علاقتها بالطيور بشكل أكبر. خلال هذه الفترة، كان يُعتقد أن وحشيات الأرجل مثل الكارنوصوريات والتيرانوصوريات بأنها تسير بوضعية شبه منتصبة بتعامد الفخذ واستقامة العمود الفقري باستخدام عضلات ذيولها الطويلة كدعم إضافي في وضع ثلاثي الأرجل كما يفعل الكنغر.[24] مع بداية العقد 1970، بدأت الدراسات البيولوجية الميكانيكية لوحشيات الأرجل العملاقة المنقرضة تشكك في هذا التفسير. تشير دراسات لمفاصل عظام الأطراف وانعدام اثر مسار جر الذيل عند المشي إلى أن وحشيات الأرجل طويلة الذيل العملاقة كان من الممكن أن تتخذ وضعية أفقية أكثر بحيث يكون الذيل موازيا للأرض.[24][25] ومع ذلك، لا يزال وضع الساقين في هذه الأنواع من الديناصورات أثناء المشي مثيرا للجدل. بعض الدراسات تدعم تقليديا عظمة الفخذ ذات الاتجاه الرأسي، على الأقل في وحشيات الأرجل ذات الذيول الطويلة والكبيرة،[25] ويقترح البعض الآخر بأن الركبة كانت تنثني بقوة في جميع وحشيات الأرجل أثناء المشي، حتى العمالقة منها كتيرانوصوريات.[26][27] يحتمل أن تكون هناك أوضاع متنوعة لانتصاب الجسم والمشي في العديد من مجموعات وحشيات الأرجل المنقرضة.[24][28]
وكما هو معروف، أن الثيروبودات تتحرك فقط على الأطراف الخلفية، فإن أصابع القدم فقط هي التي تلامس الأرض، لذلك فهي حيوانات إصبعية. يتبين من أحافير آثار الأقدام أن عرض الخطوة ضيقا جدا، وفي كثير من الأحيان تضع قدم واحدة أمام الأخرى، ويمكن تحريك الأرجل في مستوى واحد فقط (للأمام - للخلف). وبسبب تركيبة المفاصل لا تتمكن الثيروبودات من ثني الأطراف للخارج كالثدييات.
هناك محاولات مختلفة بواسطة روبرت ماكنيل ألكسندر[29] وريتشارد ثولبورن[30] لحساب سرعة الديناصورات بناءً على هيكلها العظمي وأحافيرها. ووفقا لهذه الحسابات، قد تصل سرعة الثيروبودات الأصغر إلى 40 كم/ساعة، وتصل سرعة الأورنيثوميموصوريا إلى 60 كم/ساعة. ويُفترض أن تكون سرعات الثيروبودات الأكبر منخفضة. ولا زالت هذه الحسابات مثيرة للجدل.
من المحتمل في بعض الداينونيكوصورات مثل الميكرورابتور أنها تتسلق الأشجار.
حجم الدماغ
يعتبر دماغ وحشيات الأرجل هو الأكبر نسبيا من بين الديناصورات. ولحساب حجم الدماغ طور العالم جيمس هبسون عملية النسبة الدماغية (Encephalization quotient).[31] وتعمل هذه الطريقة بحساب حجم تجويف القحف وعامل قياس التنامي حيث أن الكائنات الأكبر لها أدمغة أصغر من الكائنات الأصغر. ووضعت التماسيح كقيمة مقارنة، حيث أن قيمة النسبة الدماغية =1. وقيمة النسبة الدماغية لجميع وحشيات الأرجل >1 (للمقارنة: من بين بعض الديناصورات فقط وحشيات الأرجل التي النسبة الدماغية لها > 1، وجميع الديناصورات الأخرى أقل من ذلك. وتعتبر النسبة الدماغية لأشباه الصوروبوديات هي الأقل ثم يليها الأنكيلوصوريات والستيغوصوريات). وداخل رتيبة وحشيات الأرجل تمتلك الدرومايوصوريات أعلى معدل للنسبة الدماغية حيث تعادل 5.8 وهو الأقرب للنعام.[32]
النظام الغذائي

تعتبر وحشيات الأرجل المجموعة الوحيدة من الديناصورات التي تتغذى في الغالب على اللحوم (باستثناء محتمل لبعض الأنواع البدائية، انظر أدناه). نظرًا للتنوع الكبير في أنواع وأحجام أجسامها فقد طورت استراتيجيات غذائية مختلفة. تسمح الاكتشافات الأحفورية المعزولة كعلامات العض أو الكوبروليت (البراز المتحجر) باستخلاص استنتاجات حول النظام الغذائي، ومع ذلك، ففي كثير من الحالات تظل طريقة صيد الفرائس وأنواع الحيوانات المفترسة محل الشكوك.[33] حدد العالمين "ديفيد فاستوفسكي" و"جوشوا سميث" خمسة أنماط شكلية من وحشيات الأرجل الآكلة للحوم، وهذا ليس تصنيفا علائقيا، لهذا السبب لم يتم إعطاء أسماء للأنواع.
- النمط الشكلي الأول: يشمل الأنواع الكبيرة جدا ويصل طولها فوق 10 أمتار، وتمتلك هذه الأنواع جمجمة كبيرة، وأطراف أمامية قصيرة وعدد الأصابع لديها قليل، ومن هذه الأنواع التيرانوصور، والتربوصور، والكاركارودونتوصور، والجيجانوتوصور. تشمل هذه الأنواع أكبر الحيوانات الآكلة للحوم التي عاشت على الأرض على حتى الآن، ولا توجد مقارنات حديثة لهذه الحيوانات. ونظامها الغذائي مثير للجدل، ولا يزال النقاش يدور حول بعض الأنواع كالتيرانوصوريات هل كان قمامة أو أنها تصطاد فرائسها بنفسها، ومن الأدلة بأنها قمامة، الأسنان الواسعة المناسبة لسحق العظام، وكذلك تركيبة الأطراف. على عكس معظم وحشيات الأرجل الأخرى، فإن الفخذ والساق بنفس الطول تقريبا، مما يشير إلى أن حركتها بطيئة.
- النمط الشكلي الثاني: يشمل الأنواع المتوسطة الحجم، حوالي 3 إلى 8 أمتار. لديها جمجمة كبيرة إلى حد ما أيضا، ومقارنة بالنمط الأول فإن لديها الأطراف الخلفية أنحف والساقين أطول بكثير من الفخذين. وغالبا ما تكون الأيدي لديها متطورة. ومن هذه الأنواع الألوصور، والألبرتوصور، والدايلوفوصور، والسيراتوصور، والكارنوتوروس. ومن شبه المؤكد أن هذه الأنواع كانت صيادات نشطة، ومن المحتمل أنها كانت تهاجم الفرائس الكبيرة مثل سحليات الأرجل.
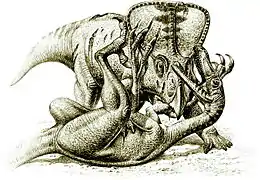

- النمط الشكلي الثالث: يشمل الأنواع الصغيرة الحجم نوعًا ما (من 2 إلى 6 أمتار)، لديها جمجمة كبيرة وعيونها متجهة إلى الأمام، ولها أذرع طويلة وأيدي كبيرة بثلاثة أصابع للإمساك، وأجسامها نحيلة ولديها ذيل صلب بشكل ملحوظ، ويتميز إصبع القدم الثاني لديها بمخلب حاد متحرك. ومن هذه الأنواع الدينونيكوس، والترودون، والفيلوسيرابتور واليوتارابتور. يقول عنها العالم جون أستروم بأنها "آلات قتال أساسية".[34] ومن المحتمل أن تكون هذه الحيوانات سريعة الجري ورشيقة، وصيادات ماهرة، وقد تكون في مجموعات لملاحقة فرائسها، ويشار إليها بأنها "الديناصورات المحاربة". عُثر على عينة من ديناصور الفيلوسيرابتور مونجولينسيس وهي في معركة مع البروتوسيراتوبس أندروسي (أحد أنواع طيريات الورك وكلاهما نفقا ربما بسبب عاصفة رملية).
- النمط الشكلي الرابع: يشمل أيضا الأنواع الصغيرة الحجم نوعا ما (من 2 إلى 6 أمتار). وهي مثل النمط الشكلي الرابع أجسامها نحيلة وأذرعها طويلة بأيدي كبيرة، لكنها تختلف في الجمجمة الأصغر وأطرافها الخلفية القصيرة نوعا ما. ومن هذه الأنواع السيلوفايسس، والكومبسوغناثوس، والميغابنوصور. وهي من الصيادات النشطة أيضا، ومن فرائسها الحيوانات الصغيرة كالسحالي، ووحشيات الأقواس (وكذلك الثدييات) أو تتغذى على الحشرات. وأظهرت بعض العينات دليلاً مباشرا على أن ليديها سلوك الافتراس، فقد تم العثور على أحفورة لديناصور الكومبسوغناثوس لونغيبس وقد كانت في معدته سحلية، وكذلك وجدت أحافير لحيوانات صغيرة في معدة السيلوفايسس.
- النمط الشكلي الخامس: يشمل أيضا الأنواع الصغيرة الحجم (من 2 إلى 6 أمتار). لكنها تتميز بأنها لا تمتلك أسنان، وذات جمجمة صغيرة، وجسدها نحيل والأذرع طويلة ذات أيدي كبيرة تحمل ثلاثة أصابع. ومن هذه الأنواع سارقات البيض. وتبين بنية أجسامها بأنها صيادة ماهرة (تشبه النمط الشكلي الثالث، باستثناء أنها لا تمتلك أسنان ولا مخلب في إصبع القدم)، ولكن لم يتضح ما تتغذى عليه هذه الحيوانات. وتبعا للفرضية الأساسية، فإنها تتغذى على البيض رغم أن منقارها غير مناسب، وتقول نظرية أخرى أن المنقار مناسب لتكسير المحار، لكن هذا مثير للجدل، لأنه لم يعثر على أي دليل على وجود الماء في عدد من المواقع الأحفورية. في نهاية المطاف لا يزال النظام الغذائي لهذه الحيوانات غير واضح، والتخمينات قول بأنها السحالي والثدييات الصغيرة الأخرى.
تمثل السبينوصوريات حالة خاصة ضمن وحشيات الأرجل الآكلة للحوم، حيث أن أسنانها مخروطية الشكل وأسنانها الأمامية كبيرة. ويعتقد أنه سبب تشابهها بالتماسيح هو لأن بعض هذه الحيوانات تأكل الأسماك الكبيرة. وفي الواقع فقد تم العثور على قشور للسمك وبقايا الإغواندون الصغير في تجويف بطن الباريونيكس، الذي يعتقد أنه له نظام غذائي مختلط من الأسماك والحيوانات البرية.[35][36]
بالرغم من أن الغالبية العظمى من وحشيات الأرجل آكلة للحوم، إلا أن مجموعتين منها قد تطورت عكس البقية وأصبحت حيوانات عاشبة:

- المجموعة الأولى هي الديناصورات المحاكية للطيور التي ليس لديها أسنان باستثناء عدد قليل من الأنواع القاعدية التي ينتهي فمها بمنقار مشابه لمنقار الطيور. وقد كانت أجسامها نحيلة ومناسبة للسرعات العالية، وأطرافها الأمامية ضعيفة، قد تكون مناسبة لسحب الفروع. وكان لديها حصيات المعدة كما في الطيور. وتقول الفرضيات حول النظام الغذائي لهذه الحيوانات بأنها إما عواشب أو آكلة اللحوم، على غرار النعام في العصر الحديث. واكتشف بعض الباحثين أيضًا دليلا على وجود بنية في المنقار على شكل المشط مما يدل على علامة لترشيح تناول الطعام. بينت جميع الاكتشافات أن غالبية الديناصورات المحاكية للطيور كانت بالقرب من موائل المياه العذبة، وما زالت هذه الاكتشافات مثيرة للجدل.
- المجموعة الثانية هي التريزينوصوريات، وهي صغيرة البنية، لديها أسنان ملوقية الشكل،[37] ولها أيدي كبيرة بشكل لافت ولها مخالب طويلة، وحوضها ذيلي مما يجعل الجذع لديها واسعا، وبالتالي يخلق مساحة أكبر للجهاز الهضمي الطويل. تشابه طريقة حياتها المفترضة أحيانا بكسلان الأرض العملاق، حيث يفترض أنها تسحب الأغصان بمخالبها الطويلة للوصول إلى الطعام.
تظهر سلالات عديدة أخرى من مانيرابتورات المبكر تكيفها للنظام الغذائي القارت (آكل للحوم والنبات معا)، بالإضافة إلى أكل البذور (بعض الترودونتيدات)، وآكلة للحشرات (العديد من طيريات الأجنحة والألفاريزاصورات).
التكاثر والتطور

مثل جميع الديناصورات وكذلك طيور هذا اليوم، جميع وحشيات الأرجل تضع البيض. وقد تم اكتشاف العديد من الهياكل العظمية لأنواع من سارقات البيض وهي جالسة، في الماضي كان يُعتقد أنها آكلة للبيض، ومن هنا تمت التسمية "سارقات البيض" أما اليوم فقد تبين أنها تحتضن البيض. في الغالب تحتوي أعشاشها على 22 بيضة، وتجلس البالغة منها فوق منتصف العش وتمسك بذراعيها العش. كما تم العثور على أعشاش أحفورية أخرى لكن لم يتم تحديد لأي مجموعة تنتمي.
يتميز تطور وحشيات الأرجل بالنمو السريع، يزداد حجم الفريخ حت يبلغ بمقدار 10 إلى 15 مرة. كما أن الصغيرة منها تختلف في بنيتها الجسدية؛ لديها حجم الجمجمة مختلف مع فكين أصغر، ورقبة أقصر والأرجل خلفية أطول. لا تعرف الحيوانات الصغيرة إلا من خلال عدد قليل من الأنواع، على سبيل المثال من السيلوفايسيات الميغابنوصور والسيلوفايسس أو من التيرانوصور، ولا يزال الكثير من المعلومات حول التطور غامضة جدا.
تاريخ التطور

خلال أواخر العصر الثلاثي، كان هناك عدد من ديناصوريات وحشيات الأرجل البدائية ووحشيات الأرجل وقد تطورتا جنبا إلى جنب.
كانت أقدم وحشيات الأرجل وأكثرها بدائية الإيودروماوس والهيريراصوريات الآكلة للحوم وقد عاشت في الأرجنتين (وربما كذلك، الإيورابتور). وانتشرت الهيريراصوريات خلال أواخر الثلاثي المبكر (أواخر الكارني إلى النوري المبكر)، وعثر عليها في كل من أمريكا الشمالية وأمريكا الجنوبية وربما أيضا الهند وجنوب إفريقيا. تميزت الهيريراصوريات بالخصائص الفسيفسائية البدائية والمتقدمة. في الماضي اعتبر بعض علماء الأحافير أن الهيريراصوريات أحد أعضاء وحشيات الأرجل، في حين أن البعض الآخر افترض أن المجموعة هي سحليات ورك قاعدية، وربما تكون قد تطورت قبل انقسام سحليات وطيريات الورك. يبين تحليل تصنيف تفرعي بعد اكتشاف ديناصور تاوا الآخر من العصر الثلاثي أن أحتمال الهيريراصوريات كانت من وحشيات الأرجل المبكرة.[38]
تعتبر السيلوفايسيات أقدم وحشيات الأرجل والأكثر البدائية (أو بدلا من ذلك، "وحشيات الأرجل الحقيقية"). وكانت السيلوفايسيات عبارة عن مجموعة من الحيوانات المنتشرة وذات التجمعات الخفيفة والتي يحتمل أن تكون قطيعية. وكان من بينها الصيادات الصغيرة مثل السيلوفايسس و (ربما) حيوانات مفترسة أكبر مثل الدايلوفوصورات. استمرت هذه الحيوانات الناجحة بالانتشار من أواخر الكارني (أواخر العصر الثلاثي) وحتى خلال التورسي (أواخر الجوراسي المبكر). على الرغم من أنه تم تضمينها في التصنيف التفرعي المبكرة تحت السيراتوصوريات واعتبارها فرعا جانبيا من وحشيات الأرجل أكثر تقدما،[39] إلا أنها قد تكون أسلافا لجميع وحشيات الأرجل الأخرى (مما يجعلها مجموعة شبه عرقية).[40][41]
إلى حد ما تعتبر السيراتوصوريات الأكثر تقدما (بما فيها السيراتوصور والكارنوتوروس) وقد ظهرت في لوراسيا خلال الجوراسي المبكر واستمرت حتى الجوراسي المتأخر، وقد تنافست بجانب أقاربها صلبات الذيل المتقدمة تشريحيا —في شكل سلالة الأبليصوريات— في غندوانا واستمرت حتى نهاية العصر الطباشيري.

مرة أخرى أصبحت صلبات الذيل أكثر تخصصا مقارن بالسيراتوصورات، وتم توثيقها بشكل جيد، وقد تكون مجموعة وحيدة الأصل والتي ظهرت لأول مرة في الجوراسي المبكر. بصرف النظر عن العدد القليل للأنواع القاعدية، فإنه يمكن التعرف على خطين للتطور داخل مجموعة صلبات الذيل، هما الميجالوصوريات (أو السبينوصوريدات) ووحشيات الأرجل الطيرية.
كانت الميجالوصورات من الحيوانات المفترسة الأساسية خلال الفترة من الجوراسي الأوسط إلى الطباشيري المبكر، ومعظم بقايا أقاربها من السبينوصوريات في صخور الطباشيري المبكر والأوسط.
لم يتم الاعتراف بترابط الميجالوصوريات إلا في عام 1998، وتنقسم إلى:
- الميجالوصورات (صنف من وحشيات الأرجل المتوسطة الحجم من العصر الجوراسي، منها الميغالوصور والستريبتوبونديلوس الحقيقية).
- السبينوصوريات (مجموعة من العصر الطباشيري تتميز بالرأس الممدود المشابه للتمساح، قد يتكون أنواعها التي تتغذى على الأسماك).
تعتبر وحشيات الأرجل الطيرية كما يشير اسمها أكثر ارتباطًا بالطيور، وتتميز وحشيات الأرجل الطيرية بنافذة إضافية في الفك العلوي وميزات أخرى في الجمجمة والحوض والفقرات الذيلية، وهي تنقسم إلى نوعين فرعيين:
- السيلوروصوريات (مجموعة كبيرة جدا من الديناصورات المتنوعة تشمل الطيور).
- الكارنوصوريات.
وقد تم استخدام هذين الاسمين في ما سابق لتقسيم وحشيات الأرجل إلى نوعين أكبر وأصغر، وفي النظاميات الحديث فهي مصنفة بشكل محدود بدقة أكبر. تتميز الكارنوصوريات بأنها وحشيات أرجل كبيرة نوعا ما وتتميز بوجود بفتحات أنف متضخمة وتتألف من ثلاث مجموعات رئيسية.
ومن السينرابتورات التي انتشرت بشكل رئيسي في أواخر العصر الجوراسي في آسيا:
- الألوصوريات ومنها الألوصور الشهير.
- الكاركارودونتوصوريات ومنها أكبر وحشيات الأرجل المعروفة الكاركارودونتوصور والجيجانوتوصور.
وهكذا، خلال الجوراسي المتأخر، كان هناك ما لا يقل عن أربعة سلالات متميزة من وحشيات الأرجل —السيراتوصورات، والميجالوصورات، والألوصورات، والسيلوروصورات— والي كانت تتغذى على الديناصورات العاشبة الصغيرة والكبيرة. نجت المجموعات الأربع حتى حلول العصر الطباشيري، ونجت ثلاثة منها —السيراتوصورات، والسيلوروصورات، والألوصورات— حتى نهاية الفترة، حيث كانت منفصلة جغرافيا، كانت السيراتوصورات والألوصورات في غندوانا، والسيلوروصورات في لوراسيا.
كانت السيلوروصوريات الأكثر تنوعا من بين كل مجموعات وحشيات الأرجل، وهي الصنف التي بدأ يتطور الريش لديها وفقًا لسجل الحفريات.
بالإضافة إلى بعض الأنواع الأساسية مثل الأورنيثولستس فإن وبعض مجموعات السيلوروصوريات التي ازدهرت خلال العصر الطباشيري هي:
- التيرانوصوريات (وهي وحشيات الأرجل الكبيرة جدا ومنها التيرانوصور) حيث تتميز الأنواع اللاحقة بالرأس الضخم والأطراف الأمامية قصيرة..
- الدرومايوصوريات (بما في ذلك الفيلوسيرابتور والدينونيكوس، التي تشبه الأركيوبتركس أقدم طائر معروف[42][43]).
- الترودونتيدات وسارقات البيض التي تشبه الطيور.
- الأورنيثوميموصوريا (أو "ديناصورات النعام") والتي يُفترض أنها عاشبة أو قارتة.
- التريزينوصورات العاشبة ذات المخالب العملاقة الغريبة.
- المانيرابتورا التي تتميز بطول الأطراف والأيدي الأمامية، ومن بينها:
- سارقات البيض التي غالبا ما تشكل أحد أنواع الديناصورات التي بلا أسنان،
- التريزينوصوريا التي لديها مخالب طويلة جدا على الأرجل الأمامية وقد تكون من آكلات النباتات،
- الداينونيكوصورات الأصغر نوعا ما، والمفترسة وتتميز بمخلب كبير على إصبع القدم الثاني، وتشمل الفيلوسيرابتور والدينونيكوس.
- طيريات الأجنحة التي تشمل الطيور الحديثة وهي سلالة الديناصورات الوحيدة التي نجت من حدث انقراض العصر الطباشيري-الباليوجيني.[44]
من نظرة علم الوراثة العرقي، فإن الطيور تنتمي أيضًا إلى المانيرابتورا. والمثير للجدل داخل مجموعة المانيرابتورا هو موضع الألفاريزصوريات التي تجمع ميزات شبيهة بالطيور والسمات البدائية، ولكن بسبب خصائصها، وقصر اذرعها فإنه ليس من السهل تصنيفها.
بينما كانت جذور المجموعات السابقة موجودة في الجوراسي الأوسط، إلا أنها أصبحت وفيرة فقط خلال الطباشيري المبكر. اقترح عدد قليل من علماء الأحافير مثل غريغوري باول أن بعض أو كل وحشيات الأرجل المتقدمة قد انحدرت بالفعل من الديناصورات الطائرة أو الطيور البدائية مثل الأركيوبتركس التي فقدت القدرة على الطيران وعادت إلى موطنها الأرضي.[45]
في 31 يوليو 2014، أبلغ العلماء عن تفاصيل تطور الطيور من ديناصورات وحشيات الأرجل الأخرى.[46][47][48] من بين السمات التي تربط الديناصورات وحشيات الأرجل بالطيور هي عظم الفريقة (عظم الترقوة)، وهي عظام مملوءة بالهواء، حضن البيض، والريش (في السيلوروصوريات، على الأقل).
النظاميات
النظاميات الخارجية

تم وصف وحشيات الأرجل بتعريف التفرع الأساسي كـ"جميع الأصناف التي ترتبط ارتباطا وثيقا بالعصفور الدوري (عصفور المنزل) من السيتيوصور أوكسونيسيس (سحليات الأرجل)".[49] وهي تقليديا تصنف في مجموعة سحليات الورك والتي تشمل أيضا أشباه سحليات الأرجل. يمكن إظهار هذه السلالة في مخطط التصنيف التفرعي التالي:
| ديناصورات |
| ||||||||||||

هناك بعض الديناصورات البدائية أصبح تصنيفها التنظيمي مثيرا للجدل ومن الأمثلة عليها الإيورابتور والهيريراصوريات (منها الهيريراصور والستوريكوصور). هناك دراسات ذات نتائج مختلفة بالنسبة للأنواع الأولية من بداية العصر الثلاثي المتأخر، لذلك تعتبر أحيانا خارج منزلة الديناصورات، في بعض الأحيان أحد الأنواع القاعدية من وحشيات الأرجل وأحيانا أحد الأنواع القاعدية من سحليات الورك. أجرى العالم "ماكس لانجر"[50] دراسة لعلم الوراثة لهذه الأنواع المثيرة للجدل وصنفت على أنها سحليات الورك، لكنه يرى أن وحشيات الأرجل أكثر ارتباطًا بـأشباه سحليات الأرجل، وهذا هو سبب تصنيفها على أنها سحليات ورك قاعدية (وليس وحشيات الأرجل). بسبب عدم اكتمال المواد للنظاميات، فلا يوجد اتفاق نهائي عليها.[51]
في عام 2017 تم اقتراح نظام عرقي جديد للديناصورات. ووفقا لهذه الفرضية، لم تعد وحشيات الأرجل مرتبطة بسحليات الورك، ولكن مع طيريات الورك فإن الأورنيثوسكيليدات تشكل المجموعة الشقيقة لسحليات الأرجل:[52]
| ديناصورات |
| ||||||||||||||||||
النظاميات الداخلية
إذا احتسبت الأنواع المثيرة للجدل المذكورة في الأعلى من بين وحشيات الأرجل، كما أقترح العالم باول سيرينو.[53] حيث تعتبر وحشيات الأرجل المتبقية مجموعات شقيقة لتلك الأنواع فسيتم ضمها معا على أنها وحشيات الأرجل الحديثة.

بعيدا عن هذا، يمكن تقسيم وحشيات الأرجل بسهولة إلى مجموعتين، السيراتوصوريات وصلبات الذيل. وتنقسم السيراتوصوريات انقساما مبكرا مرة أخرى إلى صنفين، الفئة الأولى السيلوفايسيات، وهي مجموعة الديناصورات الأصغر والتي كانت منتشرة في العصر الثلاثي المتأخر والجوراسي المبكر، وبصرف النظر عن المجموعات المثيرة للجدل فهي تعتبر وحشيات الأرجل الأقدم. الفئة الثانية من السيراتوصوريات هي السيراتوصوريات الحديثة. بالإضافة إلى الأنواع الأوائل مثل السيراتوصور فهي تشمل الأبليصوريات، التي كانت في العصر الطباشيري هي الحيوانات المفترسة المهيمنة في جنوب القارة غندوانا. ولا تخلو النظاميات داخل السيراتوصوريات من الجدل، كما أنها غير واضحة حتى لو كانت من أصل واحد.
التصنيف
تاريخ التصنيف

صاغ أوثنييل مارش اسم وحشيات الأرجل لأول مرة في عام 1881.[54] في البداية صنف مارش وحشيات الأرجل كرتيبة لتشمل فصيلة الألوصوريات، لكنه وسع نطاقها فيما بعد، وأعاد ترتيبها لتصبح رتبة تشمل مجموعة واسعة من فصائل الديناصورات "الآكلة للحوم"، وتشمل ميجالوصورات، وأنيقات الفك، والأورنيثوميموصوروات، والبلاتيوصوريات والأنشيصور (يُعرف الآن بأشباه سحليات الأرجل العاشبة) والهالوبويات (كشف عنها لاحقا كأقارب للتمساحيات). بسبب نطاق رتب وحشيات الأرجل التي صنفها مارش، فقد جاءت لتحل محل مجموعة تصنيفية سابقة أنشأها إدوارد كوب في عام 1866 للديناصورات الآكلة للحوم أطلق عليها (Goniopoda) ("مزوية الأرجل").[55]
مع بداية القرن العشرين، رفض بعض علماء الأحافير مثل فريدريك فون هيون أعتبار الديناصورات الآكلة للحوم قد شكلت مجموعة طبيعية. وقد تخلى فريدريك عن اسم "وحشيات الأرجل"، وبدلاً من ذلك استخدم رتبة سحليات الورك لهاري سيلي التي قسمها هيون إلى رتيبتين السيلوروصوريات والبكابودصوريات. وضع فريدريك معظم مجموعات وحشيات الأرجل الصغيرة في السيلوروصوريات، ووحشيات الأرجل الكبيرة والبروسوروبودات في البكابودصوريات، التي اعتبرها أسلاف سحليات الأرجل (كان لا يزال يُعتقد أن البروسوروبودات آكلة للحوم في ذلك الوقت، بسبب الارتباط غير الصحيح بين جماجم وأسنان تمساحيات راو مع أجسام البروسوروبود في الحيوانات مثل التيراتوصور).[55] في وصف ويليام ديلر ماثيو وبارنوم براون في عام 1922 لأول الدرومايوصوريات المعروفة (درومايوصور البرتنسس[56])، أصبحا أول علماء الحفريات اللذين يستبعدون البروسوروبودات من الديناصورات الآكلة للحوم، وقد حاولا إحياء اسم "Goniopoda" ("مزوية الأرجل") لتلك المجموعة، لكن لم يقبل العلماء الآخرين أيًا من هذه الاقتراحات.[55]

في عام 1956، عاد مصطلح "وحشيات الأرجل" إلى الاستخدام -كأصنوفة تحتوي على الديناصورات الآكلة للحوم وسلالتها- عندما أعاد ألفرد رومر تصنيف رتبة سحليات الورك إلى رتيبتين: وحشيات الأرجل و سحليات الأرجل. بقي هذا التقسيم الأساسي حتى حلول علم الحفريات الحديث، ما عدا البروسوروبودات مرة أخرى، الذي أدرجها رومر باعتبارها دون رتبة لوحشيات الأرجل. وقد حافظ رومر أيضا على الانقسام بين السيلوروصوريات والكارنوصوريات (والذي صنفها أيضا على أنها دون رتبة). في عام 1969 أخل هذا الانقسام باكتشاف الدينونيكوس والدينوكيروس، بحيث أنه لا يمكن تصنيف أي منهما بسهولة على أنهما "كارنوصور" أو "سيلوروصور". في ضوء هذه الاكتشافات وغيرها، أنشأ رينشن بارسبولد بحلول أواخر العقد 1970 سلسلة جديدة من دون رتب وحشيات الأرجل: السيلوروصوريات، والداينونيكوصورات، وسارقات البيض، الكارنوصوريات، الأورنيثوميموصوريا، والدينوكيروصوريات.[55]
مع ظهور التصنيف التفرعي والتسميات الحيوي في العقد 1980، وتطورها في العقد 1990 والعقد 2000، بدأت صورة علاقات وحشيات الأرجل تتضح. في عام 1986 قام جاك غوتييه بتسمية العديد من المجموعات الرئيسية لوحشيات الأرجل، بما في ذلك الفرع الحيوي صلبات الذيل لفرع واحد من انقسام وحشيات الأرجل الأساسية مع مجموعة السيراتوصوريات الأخرى. مع ظهور مزيد من المعلومات حول ارتباط الديناصورات مع الطيور، تم تجميع وحشيات الأرجل الأكثر شبها بالطيور في الفرع الحيوي المانيرابتورا (تم تسميتها أيضا بواسطة غوتييه في عام 1986). جاءت هذه التطورات الجديدة مع اعتماد معظم العلماء بأن الطيور نشأت مباشرة من وحشيات الأرجل المانيرابتوراتية، وعلى التخلي عن رتبة في التصنيف التفرعي، مع إعادة تقييم الطيور كمجموعة فرعية من ديناصورات وحشيات الأرجل التي نجت من انقراضات حقبة الحياة الوسطى وعاشت حتى الوقت الحاضر.[55]
المجموعات الرئيسية




.jpg.webp)
فيما يلي تصنيف مبسط لمجموعات وحشيات الأرجل استنادا إلى علاقاتها التطورية، وتم تنظيمها استنادا إلى قائمة أنواع الديناصورات من حقبة الحياة الوسطى التي قدمها هولتز (2007).[58] يمكن العثور على نسخة أكثر تفصيلاً في تصنيف الديناصورات. تستخدم العلامة (†) للدلالة على مجموعات التي ليس بها أعضاء على قيد الحياة.
- †هيريراصوريات (آكلات اللحوم بدائية ثنائية الأقدام)
- †سيلوفايسيات (وحشيات الأرجل صغيرة وبدائية، تشمل السيلوفايسزات وأقاربها)
- †دايلوفوصوريات (وحشيات الأرجل بدائية آكلة لحوم متوجة)
- †سيراتوصوريات (مقرن بشكل عام، آكلات لحوم جنوبية مقرنة عموما وسائدة في العصر الطباشيري)
- متيبسات الذيل (تشمل معظم وحشيات الأرجل)
- †ميجالوصوريات (مجموعة بدائية من آكلات اللحوم الكبيرة تشمل السبينوصورات الشبه مائية)
- †كارنوصوريات (ألوصورات وأقاربها مثل الكاركارودونتوصورات)
- سيلوروصوريات (وحشيات الأرجل بأحجام وكوات مختلفة)
- †كومبسوغناثيات (كويلوروصوريات بدائية ذات قوائم أمامية منخفضة)
- †تيرانوصوريات ( تيرانوصورات وأقاربها ذات قوائم أمامية منخفضة)
- †عظائيات محاكية الطيور ("محاكية النعام"؛ معظمها بلا أسنان وآكلة لحوم ومحتمل أنها عواشب)
- مانيرابتورا ("خاطفات باليد"؛ أذرعها وأصابعها طويلة ونحيفة)
- †ألفاريزصوريات (آكل حشرات صغيرة ذات قوائم أمامية منخفضة يحمل كل منها مخلب واحد ضخم)
- †عظائيات حاصدة (عواشب ثنائية الأقدام بمخالب كبيرة ورؤوس صغيرة)
- †مجنحات متسلقة ( مانيرابتورا صغيرة شجرية أصبعها الثالث طويل)
- †عظائيات سارقة البيض (معظمها بلا أسنان، نظامها الغذائي وأسلوب حياتهم غير مؤكد)
- †أركيوبتركسيات (طيور أولية صغير مجنح)
- †درومايوصوريات (وحشيات أرجل من صغيرة إلى متوسطة الحجم)
- †ترودونتيدات (وحشيات أرجل صغيرة ونحيلة)
- طيريات الأجنحة (طيور وأقاربها المنقرضة)
- †قارتات مجنحة (طيريات أجنحة بدائية كبيرة وقصيرة الذيل)
- †طيور كونفوشية (طيور صغيرة بلا أسنان)
- †طيور معاكسة وأشباهها (طائر محلق بدائي يسكن الأشجار)
- طيور حقيقية (طائر محلق متطور)
- †طيور يان وأشباهها (طيور صينية طباشيرية بلا أسنان)
- †طيور الغرب وأشباهها (طيور مائية غواصة متخصص)
- طيور (طيور منقارية عصرية وأقاربها المنقرضة)
التفرع الأصنوفي
تصنيف وحشيات الأرجل ليس كاملاً وفيه جدل. والعديد من المجموعات لديها الارتباطات القليلة، وقد وفرت الاكتشافات الجديدة رؤية جديدة تطلبت إعادة التنظيم. ولدى بعض الباحثين آراء وتصنيف مختلف اختلافا كبيرا عن بعضها البعض. أحد مخططات النسل لوحشيات الأرجل المحتمل (2004) كما يلي:[59]
| وحشيات الأرجل |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
وتوضح الشجرة التالية تركيبا لعلاقات مجموعات وحشيات الأرجل الرئيسية بناءً على دراسات مختلفة أجريت في العقد 2010.[60]
| وحشيات الأرجل |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



.png.webp)




.jpg.webp)
ظهرت أقدم وحشيات الأرجل المؤكدة (السيلوفايسيات) في العصر الثلاثي العلوي في مرحلة النوري (منذ حوالي 228 مليون سنة). من هذه النقطة عُرفت وحشيات الأرجل في حقبة الحياة الوسطى بأكملها تقريبا، وانقرضت وحشيات الأرجل غير الطيرية مع الانقراض الجماعي للديناصورات في نهاية العصر الطباشيري. (لأسباب هذا الانقراض، انظر حدود الطباشيري-الباليوجيني وانقراض الديناصورات.)
تطور الطيور

يفترض الغالبية العظمى من الباحثين أن الطيور بالتصنيف التفرعي هي وحشيات أرجل وبالتالي ديناصورات. هذا يعني، أن بعض وحشيات الأرجل مثل الداينونيكوصورات ترتبط ارتباطًا وثيقًا بالطيور أكثر من الأنواع الأخرى من هذه المجموعة وأن وحشيات الأرجل هي مجموعة شبه عرقية بدون الطيور، وأنها ليست كلها أحفادا لسلف مشترك. تشترك بعض أو حتى جميع وحشيات الأرجل بالعديد من الميزات الموجودة في الطيور الحديثة (مثل الريش، عظم الترقوة والعظام المجوفة الهوائية). في سياق تطورها تغيرت الطيور في بعض الخصائص -خاصة فيما يتعلق بالقدرة على الطيران- وتطورت بشكل كبير، على سبيل المثال في رافدة القص وفي اندماج عظام المشط مع المشط الرسغى. هناك العديد من الأشكال الفسيفسائية المعروفة (خاصة من العصر الطباشيري)، مما يجعل هذا التطور التدريجي واضحا، ومن الأمثلة على ذلك "الطائر البدائي" الأركيوبتركس، وطائر الكونفوشيوسورنيس، والطيور النقيضة والبعض الآخر. أيضًا تم دمج بعض بعض الأنواع في بعض التصنيفات مثل سارقات البيض أو الفاريزصوريات، والتي تعد وحشيات الأرجل غي طيرية.
يرى عدد قليل من الباحثين بما فيهم عالم الطيور القديمة "ألان فيدوتشيا"، بأن الطيور لم تكن من وحشيات الأرجل، ولكن قد تطورت من مجموعة قاعدية من الأركوصورات وبعض وحشيات الأرجل الشبيهة بالطيور ليست وحشيات الأرجل، لكنها طيور تكيفت مع التنقل لكن بدون طيران. وبسبب أوجه التشابه التشريحية الكثيرة، فإن غالبية العلماء يفترضون أن أصلها وحشيات الأرجل.
انظر أيضًا
مراجع
- Wilhelm Gemoll: Griechisch-Deutsches Schul- und Handwörterbuch. 9. Auflage, durchgesehene und erweiterte von Karl Vretska. Mit einer Einführung in die Sprachgeschichte von Heinz Kronasser. Freytag u. a., München u. a. 1965. Anmerkung: Der Wortstamm ποδ- des Substantivs πούς ist im Nominativ nicht erkennbar, aber im Genitiv ποδός.
- Frederick C. Mish (Hrsg.): Merriam-Webster's collegiate dictionary. 11th edition. Merriam-Webster, Springfield MA 2008, ISBN 0-87779-809-5, S. 1297.
- Alcober, Oscar A.؛ Martinez, Ricardo N. (19 أكتوبر 2010)، "A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina"، ZooKeys (63): 55–81، doi:10.3897/zookeys.63.550، PMC 3088398، PMID 21594020.
- Hendrickx, Christophe؛ Mateus, Octávio؛ Araújo, Ricardo (2015)، "A proposed terminology of theropod teeth (Dinosauria, Saurischia)"، Journal of Vertebrate Paleontology (Submitted manuscript)، 35 (e982797): e982797، doi:10.1080/02724634.2015.982797.
- Hendrickx, C؛ Mateus, O (2014)، "Abelisauridae (Dinosauria: Theropoda) from the Late Jurassic of Portugal and dentition-based phylogeny as a contribution for the identification of isolated theropod teeth"، Zootaxa، 3759: 1–74، doi:10.11646/zootaxa.3759.1.1، PMID 24869965، مؤرشف من الأصل في 9 أغسطس 2020.
- Geggel, Laura (28 يوليو 2015)، "One tough bite: T. rex's teeth had secret weapon"، Fox News، مؤرشف من الأصل في 31 أكتوبر 2018، اطلع عليه بتاريخ 01 أغسطس 2015.
- "Special Serrations Gave Carnivorous Dinosaurs an Evolutionary Edge"، مؤرشف من الأصل في 1 يناير 2019.
- "Developmental and evolutionary novelty in the serrated teeth of theropod dinosaurs"، مؤرشف من الأصل في 27 يوليو 2019.
- Senter, P.؛ Robins, J.H. (يوليو 2005)، "Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour"، Journal of Zoology, London، 266 (3): 307–318، doi:10.1017/S0952836905006989.
- Bonaparte, Novas, and Coria (1990). "Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia." Contributions in Science (Natural History Museum of Los Angeles County), 416: 41 pp.
- Göhlich, U.B.؛ Chiappe, L.M. (16 مارس 2006)، "A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago" (PDF)، Nature، 440 (7082): 329–332، doi:10.1038/nature04579، PMID 16541071، مؤرشف من الأصل (PDF) في 14 يوليو 2020.
- Czerkas, S.A., and Yuan, C. (2002). "An arboreal maniraptoran from northeast China." Pp. 63–95 in Czerkas, S.J. (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, U.S.A. PDF abridged version نسخة محفوظة 9 أغسطس 2020 على موقع واي باك مشين.
- Goehlich, U.B.؛ Tischlinger, H.؛ Chiappe, L.M. (2006)، "Juraventaor starki (Reptilia, Theropoda) ein nuer Raubdinosaurier aus dem Oberjura der Suedlichen Frankenalb (Sueddeutschland): Skelettanatomie und Wiechteilbefunde"، Archaeopteryx، 24: 1–26.
- Xu, X.؛ Zhao, Q.؛ Norell, M.؛ Sullivan, C.؛ Hone, D.؛ Erickson, G.؛ Wang, X.؛ Han, F.؛ Guo, Y. (فبراير 2009)، "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin"، Chinese Science Bulletin، 54 (3): 430–435، doi:10.1007/s11434-009-0009-6. Abstract
- R. O. Prum (1999): Development and evolutionary origin of feathers. In: Journal of Experimental Zoology. Nr. 285, S. 291–306
R. O. Prum und A. H. Brush: Which came first, the feather or the bird? In: Scientific American. Nr. 288 (2003), S. 84–93 - Xing Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao, Chengkai Jia: Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. In: طبيعة. Bd. 431, Nr. 7009, 2004, S. 680–684, دُوِي:10.1038/nature02855, PMID 15470426.
- Therrien, F.؛ Henderson, D. M. (2007)، "My theropod is bigger than yours...or not: estimating body size from skull length in theropods"، Journal of Vertebrate Paleontology، 27 (1): 108–115، doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- dal Sasso, C.؛ Maganuco, S.؛ Buffetaut, E.؛ Mendez, M. A. (2005)، "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities"، Journal of Vertebrate Paleontology (Submitted manuscript)، 25 (4): 888–896، doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2.
- Conservation International (Content Partner); Mark McGinley (Topic Editor). 2008. "Biological diversity in the Caribbean Islands." In: Encyclopedia of Earth. Eds. Cutler J. Cleveland (Washington, D.C.: Environmental Information Coalition, National Council for Science and the Environment). [First published in the Encyclopedia of Earth May 3, 2007; Last revised August 22, 2008; Retrieved November 9, 2009]. <http://www.eoearth.org/article/Biological_diversity_in_the_Caribbean_Islands> نسخة محفوظة 2013-05-23 على موقع واي باك مشين.
- Borenstein, Seth (31 يوليو 2014)، "Study traces dinosaur evolution into early birds"، أسوشيتد برس، مؤرشف من الأصل في 23 أكتوبر 2018، اطلع عليه بتاريخ 3 أغسطس 2014.
- Zoe Gough (31 يوليو 2014)، "Dinosaurs 'shrank' regularly to become birds"، بي بي سي، مؤرشف من الأصل في 09 سبتمبر 2018.
- W. Desmond Maxwell, جون أستروم: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 4, 1995, ISSN 0272-4634, S. 707–712, دُوِي:10.1080/02724634.1995.10011256.
- كينيث كاربنتر: Variation in Tyrannosaurus rex. In: Kenneth Carpenter, فيليب جيه. كوري (Hrsg.): Dinosaur Systematics. Approaches and Perspectives. Cambridge University Press, Cambridge u. a. 1990, ISBN 0-521-36672-0, S. 141–145.
- Hutchinson, J.R. (مارس–أبريل 2006)، "The evolution of locomotion in archosaurs"، Comptes Rendus Palevol، 5 (3–4): 519–530، doi:10.1016/j.crpv.2005.09.002.
- Newman, BH (1970)، "Stance and gait in the flesh-eating Tyrannosaurus"، Biological Journal of the Linnean Society، 2 (2): 119–123، doi:10.1111/j.1095-8312.1970.tb01707.x، مؤرشف من الأصل في 4 أغسطس 2020.
- K. Padian, P.E. Olsen, (1989). "Ratite footprints and the stance and gait of Mesozoic theropods." Pp. 231–241 in: D.D. Gillette, M.G. Lockley (Eds.), Dinosaur Tracks and Traces, Cambridge University Press, Cambridge.
- Paul, G.S. (1998)، "Limb design, function and running performance in ostrich-mimics and tyrannosaurs"، Gaia، 15: 257–270.
- Farlow, J.O.؛ Gatesy, S.M.؛ Holtz, Jr.؛ Hutchinson, J.R.؛ Robinson, J.M. (2000)، "Theropod locomotion"، Am. Zool.، 40 (4): 640–663، doi:10.1093/icb/40.4.640.
- R. McNeill Alexander: Estimates of speeds of dinosaurs. In: Nature. Bd. 261, Nr. 5556, 1976, S. 129–130, دُوِي:10.1038/261129a0.
- Richard A. Thulborn: Speeds and gaits of dinosaurs. In: Palaeogeography, Palaeoclimatology, Palaeoecology. Bd. 38, Nr. 3/4, 1982, ISSN 0031-0182, S. 227–256, دُوِي:10.1016/0031-0182(82)90005-0.
- James A. Hopson: Relative brain size in dinosaurs; implications for dinosaurian endothermy. In: Roger D. K. Thomas, Everett Claire Olson (Hrsg.): A Cold Look at Warm-Blooded-Dinosaurs (= American Association for the Advancement of Science Symposium. Nr. 28). Published by Westview Press for the American Association for the Advancement of Science, Boulder CO 1980, ISBN 0-89158-464-1, S. 287–310.
- Angaben zum Enzephalsationsquotient der Theropoda نسخة محفوظة 2017-02-07 على موقع واي باك مشين.
- David E. Fastovsky, Joshua B. Smith: Dinosaur Paleoecology. In: Weishampel, Dodson, Osmólska: The Dinosauria. 2004, S. 620–624.
- John H. Ostrom: Deinonychus, the ultimate Killing Machine. In: Gary D. Rosenberg, Donald L. Wolberg (Hrsg.): Dino Fest (= Paleontological Society. Special Publication. Nr. 7, قالب:ZDB). Proceedings of a Conference for the General Public held March 24–26, 1994 at Indianapolis, Indiana. Paleontological Society u. a., Knoxville TN u. a. 1994, S. 127–138.
- Longrich, Nicholas R.؛ Currie, Philip J. (فبراير 2009)، "Albertonykus borealis, a new alvarezsaur (Dinosauria: Theropoda) from the Early Maastrichtian of Alberta, Canada: Implications for the systematics and ecology of the Alvarezsauridae"، Cretaceous Research، 30 (1): 239–252، doi:10.1016/j.cretres.2008.07.005.
- Holtz, T.R.؛ Jr؛ Brinkman, D.L.؛ Chandler, C.L. (1998)، "Dental morphometrics and a possibly omnivorous feeding habit for the theropod dinosaur Troodon"، GAIA، 15: 159–166.
- Hendrickx, C.؛ Mateus, O. (2014)، "Abelisauridae (Dinosauria: Theropoda) from the Late Jurassic of Portugal and dentition-based phylogeny as a contribution for the identification of isolated theropod teeth"، Zootaxa، 3759: 1–74، doi:10.11646/zootaxa.3759.1.1، PMID 24869965، مؤرشف من الأصل في 08 أغسطس 2020.
- Nesbitt, S. J.؛ Smith, N. D.؛ Irmis, R. B.؛ Turner, A. H.؛ Downs, A.؛ Norell, M. A. (11 ديسمبر 2009)، "A complete skeleton of a Late Triassic saurischian and the early evolution of dinosaurs"، ساينس، 326 (5959): 1530–1533، doi:10.1126/science.1180350، PMID 20007898، S2CID 8349110، مؤرشف من الأصل في 16 ديسمبر 2020..
- Rowe, T., and Gauthier, J., (1990). "Ceratosauria." Pp. 151–168 in Weishampel, D. B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, University of California Press, Berkeley, Los Angeles, Oxford.
- Mortimer, M. (2001). "Rauhut's Thesis", Dinosaur Mailing List Archives, 4 Jul 2001. نسخة محفوظة 29 يوليو 2020 على موقع واي باك مشين.
- Carrano, M. T.؛ Sampson, S. D.؛ Forster, C. A. (2002)، "The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar"، Journal of Vertebrate Paleontology، 22 (3): 510–534، doi:10.1671/0272-4634(2002)022[0510:TOOMKA]2.0.CO;2.
- Ostrom, J.H. (1969)، "Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana"، Peabody Museum Natural History Bulletin، 30: 1–165.
- Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster Co. ((ردمك 0-671-61946-2))
- Dingus, L. and Rowe, T. (1998). The Mistaken Extinction: Dinosaur Evolution and the Origin of Birds. Freeman.
- Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 472 pp. ((ردمك 0-8018-6763-0))
- Borenstein, Seth (31 يوليو 2014)، "Study traces dinosaur evolution into early birds"، أسوشيتد برس، مؤرشف من الأصل في 23 أكتوبر 2018، اطلع عليه بتاريخ 3 أغسطس 2014.
- Zoe Gough (31 يوليو 2014)، "Dinosaurs 'shrank' regularly to become birds"، بي بي سي، مؤرشف من الأصل في 9 سبتمبر 2018.
- Lee, MichaelS.Y.؛ Cau, Andrea؛ Naish, Darren؛ Dyke, Gareth J. (01 أغسطس 2014)، "Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds"، ساينس، 345 (6196): 562–566، doi:10.1126/science.1252243، PMID 25082702، S2CID 37866029.
- „all taxa closer to Passer domesticus than to Cetiosaurus oxoniensis“. توماس ر. هولتز جونيور, Halszka Osmólska: Saurischis. In: Weishampel, Dodson, Osmólska: The Dinosauria. 2004, S. 22, bezugnehmend auf Arbeiten von K. Padian und C. L. May bzw. T. Maryańska.
- Max C. Langer: Basal Saurischia. In: Weishampel, Dodson und Osmólska (Hrsg.): The Dinosauria. (2004), S. 40–44.
- Das Problem der Diagnose urtümlicher Theropoden ausführlich und auf letztem Stand diskutiert in: Paul C. Sereno: The phylogenetic relationships of early dinosaurs: A comparative report. In: Historical Biology. Bd. 19, Nr. 1 = Special Issue: Early Dinosaur Evolution, 2007, ISSN 0891-2963, S. 145–155, دُوِي:10.1080/08912960601167435.
- Matthew G. Baron, David B. Norman, Paul M. Barrett: A new hypothesis of dinosaur relationships and early dinosaur evolution. In: Nature 543, 2017, S. 501–506, دُوِي:10.1038/nature21700.
- Paul C. Sereno, Catherine A. Forster, Raymond R. Rogers, Alfredo M. Monetta: Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. In: Nature. Bd. 361, Nr. 6407, 1993, S. 64–66, دُوِي:10.1038/361064a0.
- Marsh, O.C. (1881)، "Principal characters of American Jurassic dinosaurs. Part V."، The American Journal of Science and Arts، 3، 21 (125): 417–423، doi:10.2475/ajs.s3-21.125.417، S2CID 219234316.
- Rauhut, O.W. (2003). The Interrelationships and Evolution of Basal Theropod Dinosaurs. Blackwell Publishing, 213 pp. (ردمك 0-901702-79-X)
- Matthew, W. D.؛ Brown, B. (1922)، "The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta"، Bulletin of the American Museum of Natural History، 46: 367–385.
- Anderson, Ted R. (2006). Biology of the Ubiquitous House Sparrow: from Genes to Populations. Oxford: Oxford University Press. (ردمك 0-19-530411-X).
- Holtz, Thomas R. Jr. (2007) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix. نسخة محفوظة 29 أغسطس 2020 على موقع واي باك مشين.
- تبسيط وفقا لكا من ويشامبل (Weishampel)، ودودسون (Dodson) وأوسمولسكا (Osmólska): الديناصورات. 2004.
- Hendrickx, C.؛ Hartman, S.A.؛ Mateus, O. (2015)، "An Overview of Non- Avian Theropod Discoveries and Classification"، PalArch's Journal of Vertebrate Palaeontology، 12 (1): 1–73.
مشطيات القدم الطيرية (أشباه الديناصورات وأقاربها) | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||


| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||











 بوابة علم الأحياء
بوابة علم الأحياء بوابة طيور
بوابة طيور بوابة ديناصورات
بوابة ديناصورات بوابة زواحف وبرمائيات
بوابة زواحف وبرمائيات بوابة علم الأحياء القديمة
بوابة علم الأحياء القديمة