رنا معزز
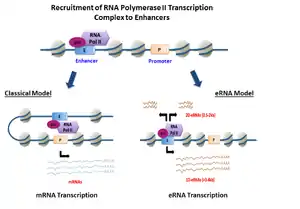
الرنا المعزز أو الرنا المحسِّن (بالإنجليزية: Enhancer RNA) هو قسم من جزيئات الرنا غير المشفرة القصيرة نسبيا (50-2000 نيوكليوتيد) وتُنسخ من تسلسلات الدنا الخاصة بالمعززات. تم اكتشافها أول مرة سنة 2010 عبر استخدام تقنيات عالية الإنتاج مثل سَلسَلة الرنا وسَلسَلة شيب.[1] يمكن تقسيم الرنا المعزز إلى قسمين فرعيين: الرنا المعزز 1D والرنا المعزز 2D ويختلفان في طولهما وحالة التذييل بعديد الأدينين واتجاهية النسخ.[2] يبدو أن التعبير عن رنا معزز معين له علاقة بنشاط المعزز الخاص به. تقترح دلائل متزايدة أن جزيئات الرنا المعزز تلعب دورا نشطا في تنظيم النسخ مقرون ومفروق ورغم أن آليات عملها مازالت غير واضحة، فقد تم اقتراح بعض الآليات.[2]
الاكتشاف
اكتُشِفت المعززات -وهي مواقع نسخ خارج جينية- أول مرة في دراسات الجينوم الكاملة والتي عرّفت المعززات على أنها مناطق شائعة لارتباط بوليميراز الرنا 2 ولنسخ جزيئات الرنا غير المشفرة.[3][1] وُجِد أن معدل تآثر بوليميراز الرنا 2 مع معزز ومعدل نسخ الرنا منه مختلفان بشكل كبير في هذه الدراسات الأولية. باستخدام ذروات توقيع الكروماتين (chromatin signature peaks) الصريح وُجِد أن نسبة معتبرة (~70%) من مواقع بدء الترجمة الخاصة ببوليميراز الرنا 2 تتداخل مع مواقع المعززات في الخلايا البلعمية الكبيرة للفئران.[3] من بين 12 ألف معزز عصبي في جينوم الفأر، وُجِد أن 25% من مواقع المعززات يرتبط بها بوليميراز الرنا 2 ويقوم بتخليق نُسَخ رنا معزز.[4] جزيئات الرنا المعزز هذه وعلى عكس جزيئات الرنا الرسول لا يتم تعديلها بإضافة ذيل عديد الأدنين وهي في العادة قصيرة وغير مشفرة للبروتين وتُنسخ في كلا الاتجاهين. كشفت دراسات لاحقة نوعا آخر من الرنا المعزز والذي يُنسَخ في اتجاه واحد وهو أطول ويملك ذيل عديد الأدينين.[5] فضلا عن ذلك، معدلات جزيئات الرنا المعزز تتناسب مع معدلات الرنا الرسول الخاصة بالجينات المجاورة، وهذا يشير إلى احتمال أن لجزيئات الرنا المعزز غير المشفرة وظيفة تنظيمية.[1]
مراجع
- Kim TK, Hemberg M, Gray JM, Costa AM, Bear DM, Wu J, Harmin DA, Laptewicz M, Barbara-Haley K, Kuersten S, Markenscoff-Papadimitriou E, Kuhl D, Bito H, Worley PF, Kreiman G, Greenberg ME (مايو 2010)، "Widespread transcription at neuronal activity-regulated enhancers"، Nature، 465 (7295): 182–7، doi:10.1038/nature09033، PMC 3020079، PMID 20393465.
- Natoli G, Andrau JC (2012)، "Noncoding transcription at enhancers: general principles and functional models"، Annual Review of Genetics، 46: 1–19، doi:10.1146/annurev-genet-110711-155459، PMID 22905871.
- De Santa F, Barozzi I, Mietton F, Ghisletti S, Polletti S, Tusi BK, Muller H, Ragoussis J, Wei CL, Natoli G (مايو 2010)، Mattick JS (المحرر)، "A large fraction of extragenic RNA pol II transcription sites overlap enhancers"، PLoS Biology، 8 (5): e1000384، doi:10.1371/journal.pbio.1000384، PMC 2867938، PMID 20485488.
- Heintzman ND, Stuart RK, Hon G, Fu Y, Ching CW, Hawkins RD, Barrera LO, Van Calcar S, Qu C, Ching KA, Wang W, Weng Z, Green RD, Crawford GE, Ren B (مارس 2007)، "Distinct and predictive chromatin signatures of transcriptional promoters and enhancers in the human genome"، Nature Genetics، 39 (3): 311–8، doi:10.1038/ng1966، PMID 17277777.
- Koch F, Fenouil R, Gut M, Cauchy P, Albert TK, Zacarias-Cabeza J, Spicuglia S, de la Chapelle AL, Heidemann M, Hintermair C, Eick D, Gut I, Ferrier P, Andrau JC (يوليو 2011)، "Transcription initiation platforms and GTF recruitment at tissue-specific enhancers and promoters"، Nature Structural & Molecular Biology، 18 (8): 956–63، doi:10.1038/nsmb.2085، PMID 21765417.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة تقانة
بوابة تقانة بوابة تقانة حيوية
بوابة تقانة حيوية بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي