Succinic semialdehyde dehydrogenase deficiency
Succinic semialdehyde dehydrogenase deficiency (SSADHD) is a rare autosomal recessive disorder[1] of the degradation pathway of the inhibitory neurotransmitter γ-aminobutyric acid, or GABA. The disorder has been identified in approximately 350 families, with a significant proportion being consanguineous families.[2] The first case was identified in 1981 and published in a Dutch clinical chemistry journal that highlighted a number of neurological conditions such as delayed intellectual, motor, speech, and language as the most common manifestations. Later cases reported in the early 1990s began to show that hypotonia, hyporeflexia, seizures, and a nonprogressive ataxia were frequent clinical features as well.[3]
| Succinic semialdehyde dehydrogenase deficiency | |
|---|---|
| Other names | 4-hydroxybutyric aciduria or Gamma-hydroxybutyric aciduria, |
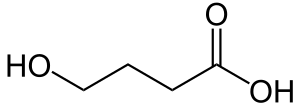 | |
| Gamma-Hydroxybutyric acid | |
SSADH deficiency is caused by an enzyme deficiency in GABA degradation. Under normal conditions, SSADH works with the enzyme GABA transaminase to convert GABA to succinic acid. Succinic acid can then be utilized for energy production via the Krebs cycle. However, because of the deficiency, the final intermediate of the GABA degradation pathway, succinic semialdehyde, accumulates and cannot be oxidized to succinic acid and is therefore reduced to gamma-hydroxybutyric acid (GHB) by gamma-hydroxybutyric dehydrogenase. This causes elevations in GHB and is believed to be the trademark of this disorder and cause for the neurological manifestations seen.[2]
Signs and symptoms
The symptoms of SSADH deficiency fall into three primary categories: neurological, psychiatric, and ocular. The most constant features seen are developmental delay, hypotonia and intellectual disability. Nearly half of patients seen manifest ataxia, behavior problems, seizures, and hyporeflexia.[2]
The age of onset ranges from newborn period to 25 years. Problems unique to neonates can include prematurity, lethargy, decreased sucking, respiratory difficulty and hypoglycemia. Gastrointestinal symptoms have been seen primarily in this population and are usually related to increased feeding.
Ocular problems related to the disorder include strabismus, nystagmus, retinitis, disc pallor, and oculomotor apraxia.[4]
Over half of the patients with SSADH deficiency have seizures. These include absence, tonic clonic, and convulsive status epilepticus. It is unclear whether decreased levels of GABA or elevated levels of GHB are responsible for these seizures but alterations in these neurotransmitters and their receptor binding or neurotransmitter transport is hypothesized to play a role in the pathogenesis of the seizures in this population.[5]
Symptoms associated with SSADHD may be mild, moderate or severe and often vary greatly from case to case. The symptoms of SSADH are caused by the accumulation of GHB in the brain and include the following manifestations (Defined as: common, > 70% of patients; frequent 30-70% of patients;unusual, < 30% of patients):
Common manifestations include:
- Delayed gross motor development
- Delayed mental development
- Delayed fine motor skill development
- Delayed speech and language development
- Hypotonia
Frequent manifestations include:
- Seizures
- Hyporeflexia
- Ataxia
- Behavioral problems
- Hyperkinesis
Unusual manifestations include:
- Neonatal problems
- EEG abnormalities
- Psychoses
- MRI or X-ray computed tomography abnormalities
- Oculomotor apraxia
- Microcephaly
- Macrocephaly
- Hyperreflexia
- Somnolence
- Choreoathetosis
- Myopathy
Genetics

SSADH deficiency is inherited in an autosomal recessive fashion. Such diseases are caused by an error in a single DNA gene. Because the disease is autosomal, the defective gene is found on an autosome (chromosome 6), rather than the sex-linked 23rd chromosome. Being a recessive disorder, the disease can only be inherited from both parents since the disorder can only occur when a person has two copies of the gene.
It is believed that the genetic basis for SSADH deficiency resides in the SSADH human ALDH5A1 gene, which maps to chromosome 6p22. More than 47 disease-causing mutations have been identified for the disorder, all of which lead to absence of functional proteins through missense, nonsense, or splicing errors; no hotspots have been identified. Consanguinity is frequent; this suggests the rare occurrence of disease-causing alleles in the general population.[6]
Mechanism
GABA is a major inhibitory neurotransmitter in the central nervous system. It modulates the activity of several neurotransmitters including dopamine, serotonin, and norepinephrine. GABA is synthesized in a single step from its precursor glutamate by glutamic acid decarboxylase. GABA is metabolized by successive transamination and oxidation to yield succinic semialdehyde and succinic acid respectively via the catalyzing effects of GABA transaminase. The succinic semialdehyde can be converted into either succinic acid by SSADH or to GHB by the enzyme succinic semialdehyde reductase.[7] The absence of SSADH leads to a 30-fold increase of GHB and a 2-4 fold increase of GABA in the brains of patients with SSADH deficiency as compared to normal brain concentrations of the compounds. Elevations of GHB have been shown to induce spike and wave activity similar to that seen in generalized absence epilepsy in animal models as well, which has motivated researchers to increase their knowledge on the relationship between GHB and the neurological manifestations seen in SSADH deficiency.[8]
GABA acts via binding to its receptors which include the ligand gated ion channels, GABAA and GABAC and the G-protein couple receptors GABAB. The GABAB receptor has been found to be the most important of the three receptors for this disorder as it is vital in both GABA and GHB release. This receptor mediates the release through presynaptic effects through a voltage dependent inhibition of high voltage activation of calcium channels. Many experiments have been able to show that it is the increased levels of both GABA and GHB that seem to alter the function of GABAB receptor, which may further play a role in the tonic-clonic seizures that are often seen in patients with the disorder.[9]
In terms of intracellular signaling, GHB inhibits mitogen activated protein (MAP) kinase action via the GABAB receptor mechanism. MAP kinase is imperative for numerous physiological changes including regulation of cell division and differentiation, thus, down-regulation of this pathway may occur during the presence of too much GHB as found in SSADH deficiency.[9] In 2003, Ren and Mody et al. proved that repeated exposure of GHB to MAP kinase affected myelin expression. This is a critical finding since myelin is the electrical and insulating phospholipid layer that surrounds the axons of many neurons in the brain. Proper myelination is critical for carrying electrical signals, or data, from one nerve cell to the next. When myelin becomes damaged, it can cause numerous neurological problems, many of which are seen in patients with SSADH deficiency. Thus, Ren and Mody's work in the relationship between increased levels of GHB and myelin expression may further show the significance of this pathway in terms of the neurological deficits seen in SSADH deficiency.[10]
Glutamine metabolism may also play a role in the pathophysiology of SSADH deficiency. The major ionotropic glutamine receptors include the N-methyl-D-aspartate (NMDA) and alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA)/kainite receptor. High levels of GHB have been shown to depress both the NMDA and AMPA/kainite receptor mediated functions and may also alter glutamatergic excitatory synaptic transmission as well.[11] Decreased glutamine, coupled with elevated GABA, has also suggested disruption of the glutamine–glutamate shuttle which ultimately provides for astrocytic glutamine as a precursor for neuronal glutamate and GABA. This disruption has the potential to impair glutamate homeostasis and may lead to uncoupling of the normal balance between glutamatergic excitatory activity and GABAergic inhibition, and may be responsible for the convulsive seizures that are observed in this disorder.[12]
Finally, additional mitochondrial processes may also be affected by SSADH deficiency. Succinate semialdehyde is considered a reactive carbonyl and may lead to increased oxidative stress.[13] This stress is believed to contribute to the formation of free radicals in the brain tissue of animal models induced with SSADH deficiency, which further leads to secondary cell damage and death. Additionally, oxidative stress may be responsible for loss of striatal dopamine which may contribute to pathophysiology of the disease.[9]
Diagnosis
Neuroimaging
Cranial computed topography, magnetic resonance imaging, and flurodeoxyglucose positron emission topography are just some of the neuroimaging modalities that have been used to diagnose patients with SSADH deficiency. On the basis of 29 previously published cases that had imaging results available, there were some common abnormalities found. These included increased T2-weighted signal abnormalities involving the globus pallidi bilaterally and symmetrically as well as the presence of subcortical white matter. Similar abnormalities have been identified in the brainstem and cerebellar dentate nucleus.[2]
Signal intensity on a T2 image may be a result of edema or an inflammatory response. Because this type of imaging is a water detecting sequence, any form of calcification or mineralization would also appear dark, thus explaining why accumulation of extra blood or fluid would appear bright on a T2 image.[14] Another explanation for signal intensity may be demyelination since the globus pallidi are traversed by a number of myelinated axons, thus confirming Ren and Mody's 2003 work proving that repeated exposure of GHB to MAP kinase affected myelin expression, thus causing the numerous neurological dysfunctions seen in SSADH deficiency patients. Ultimately, because the globus pallidus is intimately linked with the basal ganglia and thalamus, it would be expected that some of the motor dysfunctions seen in SSADH patients such as ataxia and hyporeflexia would be common.[15]
Laboratory
Detection of the disorder is possible with an organic acid analysis of the urine. Patients with SSADH deficiency will excrete high levels of GHB but this can be difficult to measure since GHB has high volatility and may be obscured on gas chromatography or mass spectrometry studies by a high urea peak. Other GABA metabolites can also be identified in urine such as glycine. Finally, succinic semialdehyde dehydrogenase levels can be measured in cultured leukocytes of the patient. This occurs due to the accumulation of 4,5-dihydroxyhexanoic acid which is normally undetectable in mammalian tissues but is characteristic of SSADH deficiency. This agent can eventually compromise the pathways of fatty acid, glycine, and pyruvate metabolism, and then become detectable in patients' leukocytes. Such enzyme levels can also be compared to non-affected parents and siblings.[2]
Treatments
A number of pharmacological treatments have been suggested or tested for efficacy on Aldh5a1-/- mice and/or humans. Below is a small sampling of the most common treatments though to be therapeutic to patients with SSADH deficiency. Unfortunately, there is very little data to support the benefit of the following treatments since few controlled studies have been conducted in patients.[16]
Two hallmarks of SSADH disorder are the increased levels of both GHB and GABA. Potential treatment modalities into biochemical and neurological correction should aim to reduce one or both while not exacerbating the other.[17]
Vigabatrin
The most common therapeutic agent available for SSADH deficiency is one that reduces the levels of GHB via inhibition of GABA transaminase. Vigabatrin is an irreversible inhibitor of GABA transaminases which leads to decreased levels of GHB and elevation of GABA. Clinical results after use are diverse, ranging from improved ataxia and speech in some patients to worsening of symptoms in others. Lower doses (30–50 mg/kg per day) is associated with fewer side effects and greater improvement of clinical features at high doses of the therapeutic. Although vigabatrin has not been consistently successful in patients with SSADH deficiency, it has shown enhanced survival of Aldh5a1-/- mice at very high doses.[9]
Sodium valproate
Sodium valproate has been used for the treatment of generalized and partial seizures in humans for both epilepsy and bipolar disorder. Valproate enhances GABA synthesis and release leading to augmented GABAergic functions in some areas of the brain. Successful interventions with valproate have been noted, but no clinical trials have been conducted thus far.[9]
However, Valproate is usually contraindicated as it may inhibit residual SSADH enzyme activity.[18]
GABAB receptor antagonist: CGP-35348
The GABAB antagonist CGP-35348 (3-amino-propyl-(diethoxymethyl) phosphinic acid) has been used in Aldh5a1-/- mice with strong results. It has shown to reduce the frequency of absence seizures, though there have been some cases in which it worsened convulsive seizures.[9]
GABAB agonist: baclofen
Baclofen (β-p-chlorophenyl-GABA) has some analgesic properties and has been traditionally used for spasticity. Its pharmacological effects primarily take place via presynaptic GABAB receptors in the spinal cord, simultaneously releasing excitatory neurotransmitters onto motor neurons. Because the number and function of GABAB receptors has been shown to progressively diminish in Aldh5a1-/- mice, such a therapy may prove to be useful. However, no data on the efficacy of baclofen on Aldh5a1-/- mice or human patients has been reported.[9]
Taurine
Taurine is a non-protein sulfur amino acid that is found in high concentrations in human milk. It has been shown to have neuroprotective and neuromodulating properties. While it is an inhibitory neurotransmitter, its ability to cross the blood brain barrier is limited. There is a lot of literature that indicates that taurine acts as antagonist at GABAA and GABAB receptors which may further enhance its ability to treat patients with SSADH deficiency, but further pharmacological studies are yet to be conducted to see if taurine could serve a therapeutic purpose.[9]
Taurine has been successfully used in a single case open study in a child with SSADH deficiency; with resolving of brain lesions, and improvement in coordination and gait.[19]
Ketogenic diet
During prolonged periods of fasting, ketone bodies serve as the primary energy source for the brain. In 2006, Henderson et al. showed that there is a therapeutic effect of maintaining a ketogenic diet – a diet consisting of high fat/low carbohydrate meals – in children with epilepsy. Ketogenic diets have also been shown to have some neuroprotective effects in models of Parkinson's disease and hypoxia as well.[20] In a 2007 study conducted at the Hospital for Sick Children in Canada, researchers found that a ketogenic diet prolonged the lifespan of Aldh5a1-/- mice by greater than 300%, along with the normalization of ataxia and some improvement in various seizure types seen in SSADH deficient murine models.[21] These effects were in conjunction with "...a significant restoration of GABAergic synaptic activity and region-specific restoration of GABAA receptor associated chloride channel binding."[21] Ultimately, the data seen in the study indicated that a ketogenic diet may work in its ability to restore GABAergic inhibition. But further studies on murine models need to be conducted, ultimately leading to the possibility of conducting a controlled study on humans afflicted with the disorder.
There is speculation that a ketogenic diet may be harmful for humans with SSADH deficiency as it may cause elevated levels of GHB in the bloodstream.
Other interventions
Other therapeutic interventions[9] include:
- ethosuximide and other anticonvulsant drugs
- GHB receptor antagonist NCS-382
- GABAA receptor modulators
- uridine
- acamprosate
- dopaminergic agents
- dextromethorphan
- glutamine
- antioxidants
- Lamotrigine[22]
The GABA(B) receptor antagonist, SGS-742, is currently being tested as a potential therapeutic in an NIH phase II clinical trial (NCT02019667).[23]
Research
While SSADH deficiency has been studied for nearly 30 years, knowledge of the disorder and its pathophysiology remains unclear. However, the progress that has been made with both murine and human models of the disorder have provided a lot of insights into how the disease manifests itself and what more can be done in terms of therapeutic interventions. Much of the current research into SSADH has been led by a dedicated team of physicians and scientists, including Phillip L. Pearl, MD of the Boston Children's Hospital at Harvard Medical School and K. Michael Gibson, PhD of Washington State University College of Pharmacy. Both have contributed significant efforts to finding appropriate therapies for SSADH deficiency and have specifically spent most of their recent efforts into understanding the efficacy of the ketogenic diet for patients with SSADH deficiency. In addition, a lot of the research that was published in 2007 examined the pathogenesis for the disorder by examining the role of oxidative stress on tissues in various cerebral structures of Aldh5a1-/- mice.
Ultimately, the metabolic pathway of SSADH deficiency is known, but how the enzyme deficiency and accumulation of GABA and GHB contribute to the clinical phenotype is not. For the future however, treatment strategies should focus on both decreasing the total production of GHB and increasing the total concentration of GABA and further assessing whether the effects of these changes influences the neurological manifestations seen in patients afflicted with SSADH deficiency.
Response to treatment is variable and the long-term and functional outcome is unknown. To provide a basis for improving the understanding of the epidemiology, genotype/phenotype correlation and outcome of these diseases their impact on the quality of life of patients, and for evaluating diagnostic and therapeutic strategies a patient registry was established by the noncommercial International Working Group on Neurotransmitter Related Disorders (iNTD).[24]
Animal models
Several scientists have developed murine models of SSADH (Aldh5a1-/-) by typical gene methodology to create a uniform absence of the SSADH enzyme activity as well as accumulations of GHB and GABA in tissues and physiological fluids. The mice are born at the expected Mendelian frequencies for an autosomal recessive disorder. Most of the models include distinctive neurological phenotypes and exhibit hypotonia, truncal ataxia, generalized tonic-clonic seizures associated with 100% mortality. The mice uniformly die at 3–4 postnatal weeks. While this model is considered to be more severe than the phenotypes seen in humans, currently, it is the most highly regarded, valid, metabolic model to study potential therapeutic interventions for the disorder.[25]
Studies have shown that alterations of both the GABAA receptor and the GABAB receptor early in the life of the Aldh5a1-/- mice can increase levels of GHB and enhance GABA release. Besides these effects, it has also been shown that "...a developmental down-regulation of GABAA receptor mediated neurotransmission in Aldh5a1-/- mice likely contributes to the progression of generalized convulsive seizures seen in mutant animals."[9] Other studies have confirmed the relationship between elevated levels of GHB and MAP kinase in mutant animals contribute to profound myelin abnormalities.
See also
References
- Chambliss, K. L.; Hinson, D. D.; Trettel, F.; Malaspina, P.; Novelletto, A.; Jakobs, C.; Gibson, K. M. (1998). "Two exon-skipping mutations as the molecular basis of succinic semialdehyde dehydrogenase deficiency (4-hydroxybutyric aciduria)". American Journal of Human Genetics. 63 (2): 399–408. doi:10.1086/301964. PMC 1377305. PMID 9683595.
- Pearl, P. L.; Novotny, E. J.; Acosta, M. T.; Jakobs, C.; Gibson, K. M. (2003). "Succinic semialdehyde dehydrogenase deficiency in children and adults". Annals of Neurology. 54 (Suppl 6): S73–80. doi:10.1002/ana.10629. PMID 12891657. S2CID 34085564.
- Jakobs, C.; Jaeken, J.; Gibson, K. M. (1993). "Inherited disorders of GABA metabolism". Journal of Inherited Metabolic Disease. 16 (4): 704–715. doi:10.1007/BF00711902. PMC 3675887. PMID 8412016.
- Pearl, P. L.; Gibson, K. M. (Apr 2004). "Clinical aspects of the disorders of GABA metabolism in children". Current Opinion in Neurology. 17 (2): 107–113. doi:10.1097/00019052-200404000-00005. ISSN 1350-7540. PMID 15021235. S2CID 24090637.
- Gibson, K. M.; Schor, D. S.; Gupta, M.; Guerand, W. S.; Senephansiri, H.; Burlingame, T. G.; Bartels, H.; Hogema, B. M.; et al. (2002). "Focal neurometabolic alterations in mice deficient for succinate semialdehyde dehydrogenase". Journal of Neurochemistry. 81 (1): 71–79. doi:10.1046/j.1471-4159.2002.00784.x. PMID 12067239. S2CID 27277845.
- Trettel, F.; Malaspina, P.; Jodice, C.; Novelletto, A.; Slaughter, C. A.; Caudle, D. L.; Hinson, D. D.; Chambliss, K. L.; Gibson, K. M. (1997). Human succinic semialdehyde dehydrogenase. Molecular cloning and chromosomal localization. Advances in Experimental Medicine and Biology. Vol. 414. pp. 253–260. doi:10.1007/978-1-4615-5871-2_29. ISSN 0065-2598. PMID 9059628.
- Nelson, D. L.; Cox M. M. (2004). Lehninger Principles of Biochemistry (4th ed.). W. H. Freeman. ISBN 978-0-7167-4339-2.
- Buzzi, A.; Wu, Y.; Frantseva, M. V.; Perez Velazquez, J. L.; Cortez, M. A.; Liu, C. C.; Shen, L. Q.; Gibson, K. M.; Snead, O. C. III (May 2006). "Succinic semialdehyde dehydrogenase deficiency: GABAB receptor-mediated function". Brain Research. 1090 (1): 15–22. doi:10.1016/j.brainres.2006.02.131. PMID 16647690. S2CID 10173411.
- Knerr, I.; Pearl, P. L.; Bottiglieri, T.; Snead, O. C.; Jakobs, C.; Gibson, K. M. (2007). "Therapeutic concepts in succinate semialdehyde dehydrogenase (SSADH; ALDH5a1) deficiency (γ-hydroxybutyric aciduria). Hypotheses evolved from 25 years of patient evaluation, studies in Aldh5a1-/- mice and characterization of γ-hyrodxybutyric acid pharmacology". Journal of Inherited Metabolic Disease. 30 (3): 279–294. doi:10.1007/s10545-007-0574-2. PMID 17457693. S2CID 20004337.
- Ren, X.; Mody, I. (2003). "Gamma-hydroxybutyrate reduces mitogen-activated protein kinase phosphorylation via GABAB receptor activation in mouse frontal cortex and hippocampus". Journal of Biological Chemistry. 278 (43): 42006–42011. doi:10.1074/jbc.M304238200. PMID 12923192. S2CID 53646170.
- Purves, Dale; Augustine, George J.; Fitzpatrick, David; Katz, Lawrence C.; LaMantia, Anthony-Samuel; McNamara, James O.; Williams, S. Mark (2001). "Glutamate Receptors". Neuroscience. 2nd Edition.
- Berton, F.; Brancucci, A.; Beghè, F.; Cammalleri, M.; Demuro, A.; Francesconi, W.; Gessa G. L. (1999). "Gamma-Hydroxybutyrate inhibits excitatory postsynaptic potentials in rat hippocampal slices". European Journal of Pharmacology. 380 (2–3): 109–116. doi:10.1016/S0014-2999(99)00515-4. PMID 10513569.
- Menduti, Giovanna; Vitaliti, Alessandra; Capo, Concetta Rosa; Lettieri-Barbato, Daniele; Aquilano, Katia; Malaspina, Patrizia; Rossi, Luisa (2020-06-19). "SSADH Variants Increase Susceptibility of U87 Cells to Mitochondrial Pro-Oxidant Insult". International Journal of Molecular Sciences. 21 (12): 4374. doi:10.3390/ijms21124374. ISSN 1422-0067. PMC 7352567. PMID 32575506.
- Liu, Chunlei; Li, Wei; Tong, Karen A.; Yeom, Kristen W.; Kuzminski, Samuel (2015). "Susceptibility-Weighted Imaging and Quantitative Susceptibility Mapping in the Brain". Journal of Magnetic Resonance Imaging. 42 (1): 23–41. doi:10.1002/jmri.24768. ISSN 1053-1807. PMC 4406874. PMID 25270052.
- Licht, Daniel MD; Stefanie Mason (2008). "Email Interview".
{{cite journal}}: Cite journal requires|journal=(help) - Didiasova, Miroslava; Banning, Antje; Brennenstuhl, Heiko; Jung-Klawitter, Sabine; Cinquemani, Claudio; Opladen, Thomas; Tikkanen, Ritva (2020-02-19). "Succinic Semialdehyde Dehydrogenase Deficiency: An Update". Cells. 9 (2): 477. doi:10.3390/cells9020477. ISSN 2073-4409. PMC 7072817. PMID 32093054.
- Pearl Phillip L.; Shukla Lovy; Theodore William H.; Jakobs Cornelis; Gibson K. Michael (2011). "Epilepsy in Succinic Semialdehyde Dehydrogenase Deficiency, a Disorder of GABA Metabolism". Brain and Development. 33 (9): 796–805. doi:10.1016/j.braindev.2011.04.013. PMC 4385391. PMID 21664777.
- Shinka, T.; Ohfu, M.; Hirose, S.; Kuhara, T. (Jul 2003). "Effect of valproic acid on the urinary metabolic profile of a patient with succinic semialdehyde dehydrogenase deficiency". Journal of Chromatography B. 792 (1): 99–106. doi:10.1016/S1570-0232(03)00276-9. ISSN 1570-0232. PMID 12829002.
- Saronwala, A.; Tournay, A.; Gargus, J. J. "Genetic inborn error of metabolism provides a unique window into molecular mechanisms underlying taurine therapy". Taurine in health and Disease. ISBN 978-81-7895-520-9.
- Henderson, C. B.; Filloux, F. M.; Alder, S. C.; Lyon, J. L.; Caplin, D. A. (2006). "Efficacy of the ketogenic diet as a treatment option for epilepsy: meta-analysis". Journal of Child Neurology. 21 (3): 193–198. doi:10.2310/7010.2006.00044. PMID 16901419. S2CID 4504870.
- Nylen, K.; Velazquez, J. L.; Likhodii, S. S.; Cortez, M. A.; Shen, L.; Leshchenko, Y.; Adeli, K.; Gibson, K. M.; Burnham, W. M.; Snead, O. C. III (2007). "A ketogenic diet rescues the murine succinic semialdehyde dehydrogenase deficient phenotype". Experimental Neurology. 210 (2): 449–457. doi:10.1016/j.expneurol.2007.11.015. PMC 2362105. PMID 18199435.
- Gibson, K. M.; Hoffmann, G. F.; Hodson, A. K.; Bottiglieri, T.; Jakobs, C. (1998). "4-Hydroxybutyric acid and the clinical phenotype of succinic semialdehyde dehydrogenase deficiency, an inborn error of GABA metabolism". Neuropediatrics. 29 (1): 14–22. doi:10.1055/s-2007-973527. PMID 9553943. S2CID 260243036.
- Parviz, M.; Vogel, K.; Gibson, K.M.; Pearl, P.L. (2014). "Disorders of GABA Metabolism: SSADH and GABA-transaminase Deficiencies". Journal of Pediatric Epilepsy. 3 (4): 217–227. doi:10.3233/PEP-14097. PMC 4256671. PMID 25485164.
- "Patient registry".
- Gropman, A. (2003). "Vigabatrin and newer interventions in succinic semialdehyde dehydrogenase deficiency". Annals of Neurology. 54: S66–S72. doi:10.1002/ana.10626. PMID 12891656. S2CID 22632304.
Further reading
- GeneReviews/NCBI/NIH/UW entry on Succinic Semialdehyde Dehydrogenase Deficiency
- Ricci L, Frosini M, Gaggelli N, Valensin G, Machetti F, Sgaragli G, Valoti M (2006). "Inhibition of rabbit brain 4-aminobutyrate transaminase by some taurine analogues: A kinetic analysis". Biochemical Pharmacology. 71 (10): 1510–1519. doi:10.1016/j.bcp.2006.02.007. PMID 16540097.
- NINDS - National Institute of Neurological Disorders and Stroke Archived 2016-12-16 at the Wayback Machine