5S ribosomal RNA
The 5S ribosomal RNA (5S rRNA) is an approximately 120 nucleotide-long ribosomal RNA molecule with a mass of 40 kDa. It is a structural and functional component of the large subunit of the ribosome in all domains of life (bacteria, archaea, and eukaryotes), with the exception of mitochondrial ribosomes of fungi and animals. The designation 5S refers to the molecule's sedimentation velocity in an ultracentrifuge, which is measured in Svedberg units (S).[1]
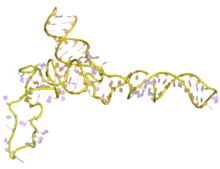
| 5S ribosomal RNA | |
|---|---|
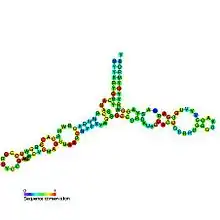 Predicted secondary structure and sequence conservation of 5S ribosomal RNA | |
| Identifiers | |
| Symbol | 5S_rRNA |
| Rfam | RF00001 CL00113 |
| Other data | |
| RNA type | Gene; rRNA |
| Domain(s) | Eukaryota; Bacteria; Archaea |
| GO | GO:0005840 GO:0003735 |
| SO | SO:0000652 |
| PDB structures | PDBe |
Biosynthesis
In prokaryotes, the 5S rRNA gene is typically located in the rRNA operons downstream of the small and the large subunit rRNA, and co-transcribed into a polycistronic precursor.[3] A particularity of eukaryotic nuclear genomes is the occurrence of multiple 5S rRNA gene copies (5S rDNA) clustered in tandem repeats, with copy number varying from species to species.[4][5] Eukaryotic 5S rRNA is synthesized by RNA polymerase III, whereas other eukaryotic rRNAs are cleaved from a 45S precursor transcribed by RNA polymerase I. In Xenopus oocytes, it has been shown that fingers 4–7 of the nine-zinc finger transcription factor TFIIIA can bind to the central region of 5S RNA.[6][7] Binding between 5S rRNA and TFIIIA serves to both repress further transcription of the 5S RNA gene and stabilize the 5S RNA transcript until it is required for ribosome assembly.[8]
Structure
The secondary structure of 5S rRNA consists of five helices (denoted I-V in roman numerals), four loops (B-E), and one hinge (A), which form together a Y-like structure. Loops C and D are terminal hairpins and loops B and E are internal.[4] According to phylogenetic studies, helices I and III are likely ancestral.[9] Helix III includes two highly conserved adenosines.[10] Helix V, with its hairpin structure, is thought to interact with TFIIIA.[4]
Location within the ribosome
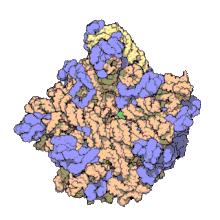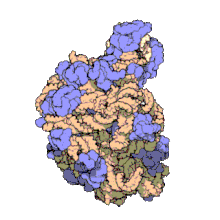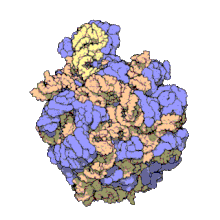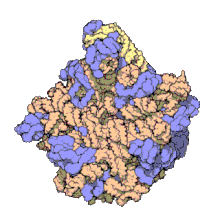
Using a variety of molecular techniques, including immuno-electron microscopy, cryo-electron microscopy, intermolecular chemical cross-linking, and X-ray crystallography, the location of the 5S rRNA within the large ribosomal subunit has been determined to great precision. In bacteria and archaea, the large ribosomal subunit (LSU) itself is composed of two RNA moieties, the 5S rRNA and another larger RNA known as 23S rRNA, along with numerous associated proteins.[3][12]
In eukaryotes, the LSU contains 5S, 5.8S, and 28S rRNAs and even more proteins.[13][14] The structure of LSU in 3-dimensions shows one relatively smooth surface and the opposite surface having three projections, notably the L1 protuberance, the central protuberance (CP), and the L7/L12 stalk. The L1 protuberance and L7/L12 stalk are arranged laterally surrounding CP. The 5S rRNA is located in the CP and participates in formation and structure of this projection. The other major constituents of the central protuberance include the 23S rRNA (or alternatively 28S in eukaryotes) and several proteins including L5, L18, L25, and L27.[15]
Ribosomal functions
The exact function of 5S rRNA is not yet clear. In Escherichia coli, 5S rRNA gene deletions reduce the protein synthesis rate and have a more profound detrimental effect on cell fitness than deletions of a comparable number of copies of other (16S and 23S) rRNA genes.[16] Crystallographic studies indicate that 5S rRNA-binding proteins and other proteins of the central protuberance of the LSU plays a role in binding tRNAs.[15] Also, the topographical and physical proximity between 5S rRNA and 23S rRNA, which forms the peptidyl transferase and GTPase-associating center, suggests that 5S rRNA acts as a mediator between the two functional centers of the ribosome by forming, together with 5S rRNA-binding proteins and other components of the central protuberance, intersubunit bridges and tRNA-binding sites.[15]
Roles in ribosomal assembly
In eukaryotes, the cytosolic ribosome is assembled from four rRNAs and over 80 proteins.[14][17] Once transcribed, the 3' ends of 5S rRNA can only be trimmed to mature length by functional homologues of RNase T, for example Rex1p in Saccharomyces cerevisiae.[18] The 60S and 40S ribosomal subunits are exported from the nucleus to the cytoplasm where they join to form the mature and translation-competent 80S ribosome. When exactly 5S rRNA is integrated into the ribosome remains controversial,[4] but it is generally accepted that 5S rRNA is incorporated into the 90S particle, which is a precursor to 60S particle, as part of a small ribosome-independent RNP complex formed by 5S rRNA and ribosomal protein L5.[17]
Interactions with proteins
Several important proteins which interact with 5S rRNA are listed below.
La protein
Interaction of 5S rRNA with the La protein prevents the RNA from degradation by exonucleases in the cell.[19] La protein is found in the nucleus in all eukaryotic organisms and associates with several types of RNAs transcribed by RNA pol III. La protein interacts with these RNAs (including the 5S rRNA) through their 3' oligo-uridine tract, aiding stability and folding of the RNA.[4][20]
L5 protein
In eukaryotic cells, ribosomal protein L5 associates and stabilizes the 5S rRNA forming a pre-ribosomal ribonucleoprotein particle (RNP) that is found in both cytosol and the nucleus. L5 deficiency prevents transport of 5S rRNA to the nucleus and results in decreased ribosomal assembly.[4]
Other ribosomal proteins
In prokaryotes the 5S rRNA binds to the L5, L18 and L25 ribosomal proteins, whereas in eukaryotes 5S rRNA is only known to bind the L5 ribosomal protein.[21] In T. brucei, the causative agent of sleeping sickness, 5S rRNA interacts with two closely related RNA-binding proteins, P34 and P37, whose loss results in a lower global level of 5S rRNA.[4]
Presence in organelle ribosomes
| Permuted mitochondrial genome encoded 5S rRNA | |
|---|---|
| Identifiers | |
| Symbol | mtPerm-5S |
| Rfam | RF02547 CL00113 |
| Other data | |
| RNA type | Gene; rRNA |
| Domain(s) | Eukaryota; |
| GO | GO:0005840 GO:0003735 |
| SO | SO:0000652 |
| PDB structures | PDBe |
Translation machineries of mitochondria and plastids (organelles of endosymbiotic bacterial origin), and their bacterial relatives share many features but also display marked differences. Organelle genomes encode SSU and LSU rRNAs without exception, yet the distribution of 5S rRNA genes (rrn5) is most uneven. Rrn5 is easily identified and common in genomes of most plastids. In contrast, mitochondrial rrn5 initially appeared to be restricted to plants and a small number of protists.[22][23] Additional, more divergent organellar 5S rRNAs were only identified with specialized covariance models that incorporate information on the pronounced sequence composition bias and structural variation.[24] This analysis pinpointed additional 5S rRNA genes not only in mitochondrial genomes of most protist lineages, but also in genomes of certain apicoplasts (non-photosynthetic plastids of pathogenic protozoa such as Toxoplasma gondii and Eimeria tenella).
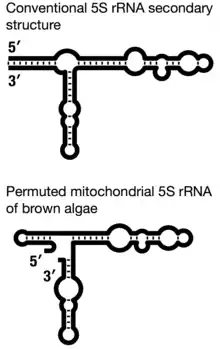
Mitochondrial 5S rRNAs of most stramenopiles comprise the largest diversity of secondary structures.[24] The permuted mitochondrial 5S rRNAs in brown algae represent the most unconventional case, where the closing helix I, which otherwise brings together the molecule's 5′ and 3′ ends, is replaced by a (closed) hairpin resulting in an open three-way junction.
Current evidence indicates that mitochondrial DNA of only a few groups, notably animals, fungi, alveolates and euglenozoans lacks the gene.[24] The central protuberance, otherwise occupied by 5S rRNA and its associated proteins (see Figure 2), was remodeled in various ways. In the fungal mitochondrial ribosomes, 5S rRNA is replaced by LSU rRNA expansion sequences.[25] In kinetoplastids (euglenozoans), the central protuberance is made entirely of evolutionarily novel mitochondrial ribosomal proteins.[26] Lastly, animal mitochondrial ribosomes have coopted a specific mitochondrial tRNA (Val in vertebrates) to substitute the missing 5S rRNA.[27][28]
See also
References
- Szymanski M, Barciszewska MZ, Erdmann VA, Barciszewski J (January 2002). "5S Ribosomal RNA Database". Nucleic Acids Research. 30 (1): 176–178. doi:10.1093/nar/30.1.176. PMC 99124. PMID 11752286.
- Mueller F, Sommer I, Baranov P, Matadeen R, Stoldt M, Wöhnert J, et al. (April 2000). "The 3D arrangement of the 23 S and 5 S rRNA in the Escherichia coli 50 S ribosomal subunit based on a cryo-electron microscopic reconstruction at 7.5 A resolution". Journal of Molecular Biology. 298 (1): 35–59. doi:10.1006/jmbi.2000.3635. PMID 10756104.
- Kaczanowska M, Rydén-Aulin M (September 2007). "Ribosome biogenesis and the translation process in Escherichia coli". Microbiology and Molecular Biology Reviews. 71 (3): 477–494. doi:10.1128/MMBR.00013-07. PMC 2168646. PMID 17804668.
- Ciganda M, Williams N (2011). "Eukaryotic 5S rRNA biogenesis". Wiley Interdisciplinary Reviews. RNA. 2 (4): 523–533. doi:10.1002/wrna.74. PMC 3278907. PMID 21957041.
- Douet J, Tourmente S (July 2007). "Transcription of the 5S rRNA heterochromatic genes is epigenetically controlled in Arabidopsis thaliana and Xenopus laevis". Heredity. 99 (1): 5–13. doi:10.1038/sj.hdy.6800964. PMID 17487217.
- McBryant SJ, Veldhoen N, Gedulin B, Leresche A, Foster MP, Wright PE, et al. (April 1995). "Interaction of the RNA binding fingers of Xenopus transcription factor IIIA with specific regions of 5 S ribosomal RNA". Journal of Molecular Biology. 248 (1): 44–57. doi:10.1006/jmbi.1995.0201. PMID 7731045.
- Searles MA, Lu D, Klug A (August 2000). "The role of the central zinc fingers of transcription factor IIIA in binding to 5 S RNA". Journal of Molecular Biology. 301 (1): 47–60. doi:10.1006/jmbi.2000.3946. PMID 10926492.
- Pelham HR, Brown DD (July 1980). "A specific transcription factor that can bind either the 5S RNA gene or 5S RNA". Proceedings of the National Academy of Sciences of the United States of America. 77 (7): 4170–4174. Bibcode:1980PNAS...77.4170P. doi:10.1073/pnas.77.7.4170. PMC 349792. PMID 7001457.
- Sun FJ, Caetano-Anollés G (November 2009). "The evolutionary history of the structure of 5S ribosomal RNA". Journal of Molecular Evolution. 69 (5): 430–443. Bibcode:2009JMolE..69..430S. doi:10.1007/s00239-009-9264-z. PMID 19639237. S2CID 21572917.
- DiNitto JP, Huber PW (October 2001). "A role for aromatic amino acids in the binding of Xenopus ribosomal protein L5 to 5S rRNA". Biochemistry. 40 (42): 12645–12653. doi:10.1021/bi011439m. PMID 11601989.
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA (August 2000). "The complete atomic structure of the large ribosomal subunit at 2.4 A resolution". Science. 289 (5481): 905–920. Bibcode:2000Sci...289..905B. CiteSeerX 10.1.1.58.2271. doi:10.1126/science.289.5481.905. PMID 10937989.
- Tirumalai MR, Kaelber JT, Park DR, Tran Q, Fox GE (October 2020). "Cryo-electron microscopy visualization of a large insertion in the 5S ribosomal RNA of the extremely halophilic archaeon Halococcus morrhuae". FEBS Open Bio. 10 (10): 1938–1946. doi:10.1002/2211-5463.12962. PMC 7530397. PMID 32865340.
- Turowski TW, Tollervey D (2015). "Cotranscriptional events in eukaryotic ribosome synthesis". Wiley Interdisciplinary Reviews. RNA. 6 (1): 129–139. doi:10.1002/wrna.1263. PMID 25176256.
- Yusupova G, Yusupov M (February 2014). "High-resolution structure of the eukaryotic 80S ribosome". Annual Review of Biochemistry. 83: 467–486. doi:10.1146/annurev-biochem-060713-035445. PMID 24580643.
- Gongadze GM (December 2011). "5S rRNA and ribosome". Biochemistry. Biokhimiia. 76 (13): 1450–1464. doi:10.1134/S0006297911130062. PMID 22339598. S2CID 41669737.
- Ammons D, Rampersad J, Fox GE (January 1999). "5S rRNA gene deletions cause an unexpectedly high fitness loss in Escherichia coli". Nucleic Acids Research. 27 (2): 637–642. doi:10.1093/nar/27.2.637. PMC 148226. PMID 9862991.
- Henras AK, Soudet J, Gérus M, Lebaron S, Caizergues-Ferrer M, Mougin A, Henry Y (August 2008). "The post-transcriptional steps of eukaryotic ribosome biogenesis". Cellular and Molecular Life Sciences. 65 (15): 2334–2359. doi:10.1007/s00018-008-8027-0. PMID 18408888. S2CID 1934099.
- van Hoof A, Lennertz P, Parker R (March 2000). "Three conserved members of the RNase D family have unique and overlapping functions in the processing of 5S, 5.8S, U4, U5, RNase MRP and RNase P RNAs in yeast". The EMBO Journal. 19 (6): 1357–1365. doi:10.1093/emboj/19.6.1357. PMC 305676. PMID 10716935.
- Wolin SL, Cedervall T (2002). "The La protein". Annual Review of Biochemistry. 71: 375–403. doi:10.1146/annurev.biochem.71.090501.150003. PMID 12045101.
- Maraia RJ, Intine RV (2002). "La protein and its associated small nuclear and nucleolar precursor RNAs". Gene Expression. 10 (1–2): 41–57. PMC 5977531. PMID 11868987.
- Moore PB (March 2001). "The ribosome at atomic resolution". Biochemistry. 40 (11): 3243–3250. doi:10.1021/bi0029402. PMID 11258942.
- Bullerwell CE, Schnare MN, Gray MW (March 2003). "Discovery and characterization of Acanthamoeba castellanii mitochondrial 5S rRNA". RNA. 9 (3): 287–292. doi:10.1261/rna.2170803. PMC 1370395. PMID 12592002.
- Bullerwell CE, Burger G, Gott JM, Kourennaia O, Schnare MN, Gray MW (May 2010). "Abundant 5S rRNA-like transcripts encoded by the mitochondrial genome in amoebozoa". Eukaryotic Cell. 9 (5): 762–773. doi:10.1128/EC.00013-10. PMC 2863963. PMID 20304999.
- Valach M, Burger G, Gray MW, Lang BF (December 2014). "Widespread occurrence of organelle genome-encoded 5S rRNAs including permuted molecules". Nucleic Acids Research. 42 (22): 13764–13777. doi:10.1093/nar/gku1266. PMC 4267664. PMID 25429974.
- Amunts A, Brown A, Bai XC, Llácer JL, Hussain T, Emsley P, et al. (March 2014). "Structure of the yeast mitochondrial large ribosomal subunit". Science. 343 (6178): 1485–1489. Bibcode:2014Sci...343.1485A. doi:10.1126/science.1249410. PMC 4046073. PMID 24675956.
- Sharma MR, Booth TM, Simpson L, Maslov DA, Agrawal RK (June 2009). "Structure of a mitochondrial ribosome with minimal RNA". Proceedings of the National Academy of Sciences of the United States of America. 106 (24): 9637–9642. Bibcode:2009PNAS..106.9637S. doi:10.1073/pnas.0901631106. PMC 2700991. PMID 19497863.
- Brown A, Amunts A, Bai XC, Sugimoto Y, Edwards PC, Murshudov G, et al. (November 2014). "Structure of the large ribosomal subunit from human mitochondria". Science. 346 (6210): 718–722. Bibcode:2014Sci...346..718B. doi:10.1126/science.1258026. PMC 4246062. PMID 25278503.
- Greber BJ, Boehringer D, Leibundgut M, Bieri P, Leitner A, Schmitz N, et al. (November 2014). "The complete structure of the large subunit of the mammalian mitochondrial ribosome". Nature. 515 (7526): 283–286. Bibcode:2014Natur.515..283G. doi:10.1038/nature13895. hdl:20.500.11850/93239. PMID 25271403. S2CID 4402484.
External links
- Page for 5S ribosomal RNA at Rfam
- 5SData Archived 2010-04-27 at the Wayback Machine
- 5S+Ribosomal+RNA at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- 5S_rRNA human gene location in the UCSC Genome Browser.
- Halococcus morrhuae (archaebacterium) 5S rRNA