Alpha-5 nicotinic acetylcholine receptor
The alpha-5 nicotinic acetylcholine receptor (α5 nAChR) also known as the α5 receptor is a type of ligand gated nicotinic acetylcholine receptor involved in pain regulation. One of the 5 transmembrane subunits of this receptor is the α5 subunit and is transcribed by the CHRNA5 gene. This receptor is commonly associated with nicotine addiction, immunotherapy, cancer, pain and attention.
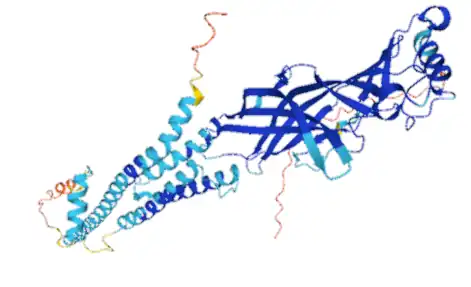
Overview
There are two major classes of acetylcholine receptors: nicotinic receptors, which bind to exogenous nicotine, and muscarinic receptors, which bind exogenous muscarine. Nicotinic acetylcholine receptors (nAChRs) were initially discovered through the application and binding of nicotine, however, endogenous acetylcholine is the ligand that binds under normal physiological conditions. The nAChRs are single channel ionotropic receptors found throughout the brain and body that allow for cations to flow in and out of cells. These receptors consist of five transmembrane subunits with the α5 nAChR defined by the presence of the α5 subunit. The α5 nAChR is located in various areas of the brain including the cortex, hippocampus, hypothalamus, inferior colliculus, medial habenula, olfactory bulb and striatum.[1] The α5 nAChR is involved in modulating chronic inflammation and peripheral nerve injury.[2]
| Alpha5 Nicotinic Acetylcholine Receptor | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | CHRNA5 | ||||||
| Alt. symbols | CHRNA5 | ||||||
| NCBI gene | 1138 | ||||||
| OMIM | 1188505 | ||||||
| RefSeq | NM_000745 | ||||||
| UniProt | P30532 | ||||||
| Other data | |||||||
| Locus | Chr. 15 q25.1 | ||||||
| |||||||
Acetylcholine binds in the cortex, hippocampus, hypothalamus, inferior colliculus, striatum and olfactory bulb.[3] CHRNA5 is located in a gene cluster on chromosome 15q24 along with CHRNA3 and CHRNB4.[4] Homopentameric receptors with five acetylcholine binding sites contain two a-subunits (a2-a4 or a6) and two non-a-subunits (B2 or B4). Alpha5 subunits tend to be the fifth that does not directly bind to acetylcholine and act as auxiliary subunits. Rather, they may be important for receptor targeting and localization on the cell membrane.[4] The alpha subunits normally assemble into both alpha3B4-containing and alpha4-beta2 containing nAChR assemblies. These receptors have been found on dopaminergic neurons in the rodent striatum and are involved in DA release upon nicotine stimulation. In addition to DA neurons, alpha5 subunits are also expressed on GABAergic neurons in the VTA and striatum.[4]
Development
The alpha5 subunit is important during the development and maturation of prefrontal pyramidal IV neurons. Cholinergic dysfunction during development causes attentional deficits observed in diseases such as schizophrenia, neurodevelopmental disorders, autism and epilepsy. Most cholinergic neurons are developed by the perinatal period in humans. Maturational changes that occur in dendrites during development are absent in alpha5 -/- mice indicating that the alpha5 subunit is necessary for proper maturation of prefrontal pyramidal cells.[5]
Nicotine addiction and withdrawal
Addiction to nicotine is modulated by the mesocorticolimbic dopamine reward system that drives the rewarding nature of nicotine; the mesocorticolimbic system is involved in self-stimulation and processing an environmental reward.[6][7] For example, this system is active while consuming highly caloric food or while gambling. Upon the administration of nicotine, there is increased firing rate mediated by midbrain dopamine neurons within this system. Through continuous exposure, dependence often occurs which is followed by withdrawal symptoms such as cravings, irritation, restlessness, sleep disturbances, weight gain, anxiety and difficulty concentrating.[8][9] Subunits involved with withdrawal syndrome include α5, α2, and B4 within the epithalamic habenular complex and its projections.[10] The medial habenula (MHb) and its projection to the interpeduncular nucleus (IPN) contain dense expression of α5 nAChR subunits.[11]
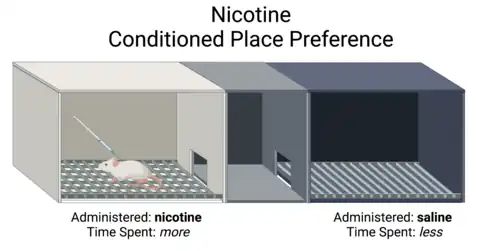
In vivo studies
Studies have shown that removing the α5 nAChR subunits from mice (α5 nAChR null) will make them less sensitive to acute effects of nicotine.[12] The mice showed decreased locomotion in an open field test and fewer nicotine-induced seizures.[11] Other studies have shown that α5 nAChR null mice display fewer signs of dependency and reduced anxiety-like behaviors.[11] Because the knockout mice show less aversion to increased nicotine intake, they tend to self-administer at much higher doses than wildtype mice. However, reintroduction of the alpha 5 subunit in the medial habenula in knockout mice restored normal levels of nicotine self-administration. This demonstrates that the expression of the alpha5 subunit in the medial habenula is sufficient to reinstate nicotine aversion.[13] In contrast, the targeted knockdown of α5 subunits in the habenulo-interpeduncular pathway in wildtype mice did not change the stimulatory effects observed in knockout mice demonstrating that the subunit is not necessary for nicotine aversion, and that other areas of the brain can compensate for this behavior.[12]
In a conditioned place preference study (CPP), researchers trained mice to associate nicotine administration with one chamber and saline administration in an adjacent chamber. At low doses of nicotine, alpha5 knockout mice and wildtype mice both showed preference for the nicotine chamber. However, at high doses of nicotine, only the α5 knockout mice preferred the nicotine chamber[13] demonstrating that the alpha5 knockout mice still experienced the rewarding aspects of nicotine, but not the normal aversive behaviors with nicotine overdose.
Studies from Tuesta et al. 2011 have shown that the dose-response curve is similar when comparing knockout mice to wildtype mice however the knockout mice consumed greater amounts of nicotine which resulted in the descending portion of the dose-response curve to descend declined slower in the knockout mice. There has been shown an increased response to nicotine in the ascending portion of the curve demonstrating the greater rewarding properties.[12]
Human studies
Nicotine is commonly consumed by people for its rewarding properties resulting in dependence, addiction and withdrawal.[14] Human studies have shown that people with a single nucleotide polymorphism (SNP) within the α5 nAChR encoding gene (CHRNA5, rs16969968) correlates with an increased risk of nicotine dependency and pleasure along with more heavy smoking.[11][15][16][17] This particular SNP results in an aspartic acid to asparagine substitution at amino acid residue 398 (D398N).[12] The rs16969968 within CHRNA5 causes a reduction in the receptor function that is involved in producing withdrawal symptoms. Individuals with this SNP are commonly found in those of European descent; there is about a 30% greater risk of nicotine dependence in individuals carry a single copy of the variant and about a 50% greater risk in individuals with two copies.[12] Other studies have shown that people with this SNP develop nicotine dependence at a younger age.[12] Researchers conclude that fewer aversive effects of nicotine would promote the rewarding effects or hedonic drive that would transition people from nicotine abuse to dependency.[11][18] Additionally, SNP variants within rs16969968 in CHRNA5 have been associated with smoking-related behaviors such has a higher prevalence of smokers vs nonsmokers.[19]
Attention and cognition
Attention is an important aspect of memory that allows for information to be held in the mind and maintain focus in the presence of distractions in order to achieve a goal directed behavior. Working memory is a similar aspect of learning, however, the main difference between the two is that working memory requires the mental manipulation of information as well. The structure most commonly associated with attention is the prefrontal cortex that mediates top down control of complex cognitive processes.[20] Acetylcholine is a neuromodulator that is closely studied for its role in learning and memory; it is involved in the acquisition, consolidation and recall.
In vivo studies
Researchers speculate that layer VI pyramidal neurons in the prefrontal cortex are important for holding attention in cognitively demanding tasks. These neurons send feedback projections to the thalamus and are highly responsive to acetylcholine. In vivo studies have shown that the presence of alpha5 subunits of nAChRs on layer VI pyramidal neurons in the PFC are important for visual attention.[21] In slice electrophysiology experiments, researchers have shown that alpha5 subunits enhance currents in the PFC of an adult mouse. In vivo, researchers use a five-choice serial reaction task. The animals are randomly given 1 of 5 light stimulus, and they need to encode and recall the location of the stimulus in order to receive a reward. Transgenic mice without the gene that encodes alpha5 subunits showed impaired performance on the five-choice serial reaction task. This indicates that the alpha5 null mice have attentional deficits.[5] Interestingly, the deletion of alpha5 subunits in mice results in an upregulation of muscarinic acetylcholine receptors as an excitatory compensation response to circuitry dysfunction. Because of the cognitive enhancing effects of alpha5 nAChR agonists, it is a common target for neurodegenerative disorders with cognitive deficits along with ADHD.[22]
Human studies
Due to technical limitations of invasive procedures, there are far fewer studies in about the role of the alpha5 nAChR subunit and cognition. Studies have performed microdialysis in subjects as they formed attention tasks and found significantly increased acetylcholine efflux.[5]
Clinical application
The α5 nAChR mediates acute effects of alcohol; a deletion in this gene affects alcohol intake under stressful conditions.[23] The α5 nAChR also mediates short term effects of nicotine.[24] Studies have shown that removing the alpha5 subunit in mice (α5 nAChR knockdown) increases nicotine intake which is rescued by reintroducing the gene.[25] Researchers have also shown that removing the α5 subunit only in the medial habenula increases nicotine self-administration, demonstrating that this subunit is necessary for normal nicotine intake and abnormalities within this subunit may contribute to the addictive properties of nicotine.[25] Additionally, the alpha5 nAChR subunit is associated with other forms of addiction such as cocaine.[26] Other studies have shown that α5 knockout mice shown impaired attentional performance.[27] During high frequency vagal stimulation, α5 nAChR knockout mice experience impaired cardiac parasympathetic ganglionic transmission. In vivo studies have also identified the alpha5 nAChR as a possible treatment for chronic inflammation and neuropathic pain.[28][29]
Ligands
| Ligand | Structure | Function | Use |
|---|---|---|---|
| Acetylcholine | Agonist | Endogenous | |
| Nicotine |  |
Agonist | Recreational drug Attention |
| Pozanicline | 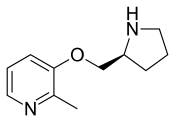 |
Partial agonist | Experimental drug for ADHD, [30] Alzheimer's disease,[31] and tobacco use disorder[32] |
| α-Conotoxin MII[33] |  |
Antagonist | |
| α-Conotoxin PnIA[33] | 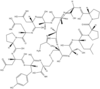 |
Antagonist | |
| α-Conotoxin GIC[33] | 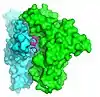 |
Antagonist | |
| α-Conotoxin TXIA[33] | 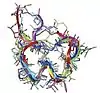 |
Antagonist |
References
- Brown RW, Collins AC, Lindstrom JM, Whiteaker P (October 2007). "Nicotinic alpha5 subunit deletion locally reduces high-affinity agonist activation without altering nicotinic receptor numbers". Journal of Neurochemistry. 103 (1): 204–215. doi:10.1111/j.1471-4159.2007.04700.x. PMID 17573823. S2CID 28394031.
- Bagdas D, AlSharari SD, Freitas K, Tracy M, Damaj MI (October 2015). "The role of alpha5 nicotinic acetylcholine receptors in mouse models of chronic inflammatory and neuropathic pain". Biochemical Pharmacology. Nicotinic Acetylcholine Receptors as Therapeutic Targets: Emerging Frontiers in Basic Research and Clinical Science (Satellite to the 2015 Meeting of the Society for Neuroscience) Oct 14-15, Chicago, IL USA. 97 (4): 590–600. doi:10.1016/j.bcp.2015.04.013. PMC 4600420. PMID 25931144.
- Brown RW, Collins AC, Lindstrom JM, Whiteaker P (October 2007). "Nicotinic alpha5 subunit deletion locally reduces high-affinity agonist activation without altering nicotinic receptor numbers". Journal of Neurochemistry. 103 (1): 204–215. doi:10.1111/j.1471-4159.2007.04700.x. PMID 17573823. S2CID 28394031.
- Greenbaum L, Lerer B (October 2009). "Differential contribution of genetic variation in multiple brain nicotinic cholinergic receptors to nicotine dependence: recent progress and emerging open questions". Molecular Psychiatry. 14 (10): 912–945. doi:10.1038/mp.2009.59. PMID 19564872. S2CID 9769700.
- Proulx E, Piva M, Tian MK, Bailey CD, Lambe EK (April 2014). "Nicotinic acetylcholine receptors in attention circuitry: the role of layer VI neurons of prefrontal cortex". Cellular and Molecular Life Sciences. 71 (7): 1225–1244. doi:10.1007/s00018-013-1481-3. PMC 3949016. PMID 24122021.
- de Kloet SF, Mansvelder HD, De Vries TJ (October 2015). "Cholinergic modulation of dopamine pathways through nicotinic acetylcholine receptors". Biochemical Pharmacology. 97 (4): 425–438. doi:10.1016/j.bcp.2015.07.014. PMID 26208783.
- Demeter E, Sarter M (January 2013). "Leveraging the cortical cholinergic system to enhance attention". Neuropharmacology. 64 (1): 294–304. doi:10.1016/j.neuropharm.2012.06.060. PMC 3445745. PMID 22796110.
- Wang JC, Kapoor M, Goate AM (2012-09-22). "The genetics of substance dependence". Annual Review of Genomics and Human Genetics. 13 (1): 241–261. doi:10.1146/annurev-genom-090711-163844. PMC 3474605. PMID 22703173.
- Wittenberg RE, Wolfman SL, De Biasi M, Dani JA (October 2020). "Nicotinic acetylcholine receptors and nicotine addiction: A brief introduction". Neuropharmacology. 177: 108256. doi:10.1016/j.neuropharm.2020.108256. PMC 7554201. PMID 32738308.
- Antolin-Fontes B, Ables JL, Görlich A, Ibañez-Tallon I (September 2015). "The habenulo-interpeduncular pathway in nicotine aversion and withdrawal". Neuropharmacology. 96 (Pt B): 213–222. doi:10.1016/j.neuropharm.2014.11.019. PMC 4452453. PMID 25476971.
- De Biasi M, Dani JA (2011-07-21). "Reward, addiction, withdrawal to nicotine". Annual Review of Neuroscience. 34 (1): 105–130. doi:10.1146/annurev-neuro-061010-113734. PMC 3137256. PMID 21438686.
- Tuesta LM, Fowler CD, Kenny PJ (October 2011). "Recent advances in understanding nicotinic receptor signaling mechanisms that regulate drug self-administration behavior". Biochemical Pharmacology. 82 (8): 984–995. doi:10.1016/j.bcp.2011.06.026. PMC 3163076. PMID 21740894.
- Stoker AK, Markou A (August 2013). "Unraveling the neurobiology of nicotine dependence using genetically engineered mice". Current Opinion in Neurobiology. 23 (4): 493–499. doi:10.1016/j.conb.2013.02.013. PMC 3735838. PMID 23545467.
- De Biasi M, Dani JA (2011-07-21). "Reward, addiction, withdrawal to nicotine". Annual Review of Neuroscience. 34 (1): 105–130. doi:10.1146/annurev-neuro-061010-113734. PMC 3137256. PMID 21438686.
- Improgo MR, Scofield MD, Tapper AR, Gardner PD (September 2010). "From smoking to lung cancer: the CHRNA5/A3/B4 connection". Oncogene. 29 (35): 4874–4884. doi:10.1038/onc.2010.256. PMC 3934347. PMID 20581870.
- Improgo MR, Scofield MD, Tapper AR, Gardner PD (October 2010). "The nicotinic acetylcholine receptor CHRNA5/A3/B4 gene cluster: dual role in nicotine addiction and lung cancer". Progress in Neurobiology. 92 (2): 212–226. doi:10.1016/j.pneurobio.2010.05.003. PMC 2939268. PMID 20685379.
- Russo P, Cesario A, Rutella S, Veronesi G, Spaggiari L, Galetta D, et al. (2010-12-31). "Impact of genetic variability in nicotinic acetylcholine receptors on nicotine addiction and smoking cessation treatment". Current Medicinal Chemistry. 18 (1): 91–112. doi:10.2174/092986711793979715. PMID 21110812.
- Greenbaum L, Lerer B (October 2009). "Differential contribution of genetic variation in multiple brain nicotinic cholinergic receptors to nicotine dependence: recent progress and emerging open questions". Molecular Psychiatry. 14 (10): 912–945. doi:10.1038/mp.2009.59. PMID 19564872. S2CID 9769700.
- Ware JJ, van den Bree M, Munafò MR (November 2012). "From men to mice: CHRNA5/CHRNA3, smoking behavior and disease". Nicotine & Tobacco Research. 14 (11): 1291–1299. doi:10.1093/ntr/nts106. PMC 3482013. PMID 22544838.
- Bloem B, Poorthuis RB, Mansvelder HD (2014-03-11). "Cholinergic modulation of the medial prefrontal cortex: the role of nicotinic receptors in attention and regulation of neuronal activity". Frontiers in Neural Circuits. 8: 17. doi:10.3389/fncir.2014.00017. PMC 3949318. PMID 24653678.
- Bailey CD, De Biasi M, Fletcher PJ, Lambe EK (July 2010). "The nicotinic acetylcholine receptor alpha5 subunit plays a key role in attention circuitry and accuracy". The Journal of Neuroscience. 30 (27): 9241–9252. doi:10.1523/jneurosci.2258-10.2010. PMC 3004929. PMID 20610759.
- Wallace TL, Ballard TM, Pouzet B, Riedel WJ, Wettstein JG (August 2011). "Drug targets for cognitive enhancement in neuropsychiatric disorders" (PDF). Pharmacology, Biochemistry, and Behavior. 99 (2): 130–145. doi:10.1016/j.pbb.2011.03.022. PMID 21463652. S2CID 25911863.
- Dawson A, Wolstenholme JT, Roni MA, Campbell VC, Jackson A, Slater C, et al. (August 2018). "Knockout of alpha 5 nicotinic acetylcholine receptors subunit alters ethanol-mediated behavioral effects and reward in mice". Neuropharmacology. 138: 341–348. doi:10.1016/j.neuropharm.2018.06.031. PMC 6400055. PMID 29944862.
- Salas R, Orr-Urtreger A, Broide RS, Beaudet A, Paylor R, De Biasi M (May 2003). "The nicotinic acetylcholine receptor subunit alpha 5 mediates short-term effects of nicotine in vivo". Molecular Pharmacology. 63 (5): 1059–1066. doi:10.1124/mol.63.5.1059. PMID 12695534. S2CID 97775995.
- Fowler CD, Lu Q, Johnson PM, Marks MJ, Kenny PJ (March 2011). "Habenular α5 nicotinic receptor subunit signalling controls nicotine intake". Nature. 471 (7340): 597–601. Bibcode:2011Natur.471..597F. doi:10.1038/nature09797. PMC 3079537. PMID 21278726.
- Forget B, Icick R, Robert J, Correia C, Prevost MS, Gielen M, et al. (February 2021). "Alterations in nicotinic receptor alpha5 subunit gene differentially impact early and later stages of cocaine addiction: a translational study in transgenic rats and patients" (PDF). Progress in Neurobiology. 197: 101898. doi:10.1016/j.pneurobio.2020.101898. PMID 32841724. S2CID 221235364.
- Bailey CD, De Biasi M, Fletcher PJ, Lambe EK (July 2010). "The nicotinic acetylcholine receptor alpha5 subunit plays a key role in attention circuitry and accuracy". The Journal of Neuroscience. 30 (27): 9241–9252. doi:10.1523/jneurosci.2258-10.2010. PMC 3004929. PMID 20610759.
- Antolin-Fontes B, Ables JL, Görlich A, Ibañez-Tallon I (September 2015). "The habenulo-interpeduncular pathway in nicotine aversion and withdrawal". Neuropharmacology. 96 (Pt B): 213–222. doi:10.1016/j.neuropharm.2014.11.019. PMC 4452453. PMID 25476971.
- Decker MW, Meyer MD, Sullivan JP (October 2001). "The therapeutic potential of nicotinic acetylcholine receptor agonists for pain control". Expert Opinion on Investigational Drugs. 10 (10): 1819–1830. doi:10.1517/13543784.10.10.1819. PMID 11772288. S2CID 24924290.
- "Safety and Tolerability Study of ABT-089 in Adults with Attention-Deficit/Hyperactivity Disorder (ADHD)". ClinicalTrials.gov.
- "Study of the Long-Term Safety of ABT-089 for Subjects with Mild-to-Moderate Alzheimer's Disease". ClinicalTrials.gov.
- Sullivan JP, Donnelly-Roberts D, Briggs CA, Anderson DJ, Gopalakrishnan M, Xue IC, et al. (October 1997). "ABT-089 [2-methyl-3-(2-(S)-pyrrolidinylmethoxy)pyridine]: I. A potent and selective cholinergic channel modulator with neuroprotective properties". The Journal of Pharmacology and Experimental Therapeutics. 283 (1): 235–246. PMID 9336329.
- Gotti C, Marks MJ, Millar NS, Wonnacott S (2019-09-16). "Nicotinic acetylcholine receptors (version 2019.4) in the IUPHAR/BPS Guide to Pharmacology Database". IUPHAR/BPS Guide to Pharmacology CITE. 2019 (4). doi:10.2218/gtopdb/F76/2019.4. ISSN 2633-1020. S2CID 203817110.