Aristonectes
Aristonectes (meaning "best swimmer") is an extinct genus of large elasmosaurid plesiosaurs that lived during the Maastrichtian stage of the Late Cretaceous. Two species are known, A. parvidens and A. quiriquensis, whose fossil remains were discovered in what are now Patagonia and Antarctica. Throughout the 20th century, Aristonectes was a difficult animal for scientists to analyze due to poor fossil preparation, its relationships to other genera were uncertain. After subsequent revisions and discoveries carried out from the beginning of the 21st century, Aristonectes is now recognised as the type genus of the subfamily Aristonectinae, a lineage of elasmosaurids characterized by an enlarged skull and a reduced length of the neck.
| Aristonectes Temporal range: Late Cretaceous, Maastrichtian, ~ | |
|---|---|
 | |
| Life restoration of A. parvidens | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Superorder: | †Sauropterygia |
| Order: | †Plesiosauria |
| Family: | †Elasmosauridae |
| Subfamily: | †Aristonectinae |
| Genus: | †Aristonectes Cabrera, 1941 |
| Type species | |
| Aristonectes parvidens Cabrera, 1941 | |
| Other species | |
| |
| Synonyms | |
|
List
| |
Measuring more than 10 m (33 ft) long, Aristonectes is a notably imposing plesiosaur. A referred specimen discovered in Antarctica has an estimated size of more than 11 m (36 ft) long, which would make this genus one of the largest known plesiosaurs. The paddle-like limbs of Aristonectes are unusually large for an elasmosaurid, reaching almost 3 m (9.8 ft) in length, suggesting a total wingspan of around 7 m (23 ft) for the animal. The skull is ogival-shaped and is flattened, possessing sharp forward-facing teeth. According to estimates, 13 teeth would have been present in each premaxilla, 50 teeth in the maxilla and 50 to 63 or more teeth in the lower jaw.
According to its morphology, mainly cranial, Aristonectes fed by mixing prey and sediment in the benthic zones, like the modern gray whale. As in other plesiosaurs, gastroliths (stomach stones) would have been used by Aristonectes either to help digest its food or to aid in buoyancy, although there is little support for the first hypothesis. According to its geographical distribution in the fossil record, Aristonectes would have regularly migrated between Patagonia and Antarctica, which he encountered many bony fishes, cartilaginous fishes, invertebrates and other marine reptiles, including other plesiosaurs and mosasaurs.
Research history
A. parvidens
The first Aristonectes fossil was discovered long before the genus was named by Ángel Cabrera in 1941.[1] In 1848, the Franco-Chilean naturalist Claude Gay described the first known plesiosaur from South America, Plesiosaurus chilensis, on the basis of a single caudal vertebra discovered in Quiriquina Island, in Concepción Province, Chile.[2] In a book published in 1889, Richard Lydekker placed this species in the genus Cimoliosaurus.[3] In 1895, Wilhelm Deecke moved it to the pliosaurid genus Pliosaurus as Pliosaurus chilensis, and referred other fossils that have been discovered in the same locality to it. Among these fossils are some vertebrae, a femur and fragments of an ischium as well as ribs.[4] In 1918, the Russian paleontologist Pavel A. Pravoslavlev moved it into the related genus Elasmosaurus, as Elasmosaurus chilensis.[5] In 1941, Cabrera described a different specimen from Chubut Province, Argentina as the new genus and species Aristonectes parvidens. In the same publication, he also moved Elasmosaurus chilensis back into Plesiosaurus with doubts (therefore resurrecting its original name "Plesiosaurus" chilensis), and compared it with his new species A. parvidens.[6] In 1949, Edwin H. Colbert found that the holotype specimen of "P." chilensis (the caudal vertebra originally described by Gay) belongs to a pliosauroid, but seemed uncertain about its generic placement. Also in the article, he considers that the other fossil material previously attributed to "P." chilensis could come from elasmosaurids.[7][8] After a further analysis published in 2013 by José P. O'Gorman and colleagues, the holotype specimen of "P." chilensis was recognized as potentially belonging to A. parvidens. As this specimen merely consists of a single caudal vertebra, it was referred to as A. cf. parvidens to indicate uncertainty in its assignment.[1]
The first specimen formally identified as A. parvidens was collected by Cristian S. Petersen in collaboration with local resident Victor Saldivia, in the Cañadón de los Loros, near the locality of Paso del Sapo in Chubut Province, Argentina.[6] The specimen was discovered in the Lefipán Formation and was dated to the Maastrichtian.[9][10][11] In September 1940, Pablo Groeber sent this specimen to the Museo de La Plata, as a donation for the Dirección de Minas y Geología del Ministerio de Agricultura (Directorate of Mines and Geology of the Ministry of Agriculture). A vertebra and incomplete phalanges from the same region had previously been donated to the Museo de La Plata by Mario Reguiló, and were subsequently referred to the holotype, catalogued as MLP 40-XI-14-6, as they most likely came from the same specimen.[6] The holotype consists of a partial skull attached to the mandible, the atlas and axis, 21 cervical vertebrae (including 16 anterior cervicals which are articulated), 8 caudal vertebrae and an incomplete left forelimb. The specimen is interpreted as an adult, or even an elderly individual.[9][12] Cabrera described the animal as Aristonectes parvidens in his article published in 1941 by the museum that houses the fossil material.[6] The genus name Aristonectes comes from the Ancient Greek words ἄριστος (áristos, "best", "superior") and νηκτός (nêktós, "swimmer"), and may be translated as "best swimmer", so named to indicate the best specimen of plesiosaur found at that time in Argentina. The specific epithet parvidens literally means "small teeth", in reference to the rather small size of the tooth sockets of this taxon.[13]
Since the unknown parts of the original specimen were reconstructed from added casts of the fossils, the precise anatomy of Aristonectes remained uncertain throughout the 20th century.[14][15][16] In 2003, Spanish paleontologist Zulma Gasparini and colleagues re-prepared the holotype skull along with the atlas and axis, in an attempt to clarify its anatomy.[9] In a doctoral thesis published in 2013, O'Gorman informally assigns several caudal vertebrae to A. parvidens.[10] In 2016, the same author revised the holotype again and described new details not reported in previous studies concerning this species.[11]
A. quiriquensis
The first discovery of the second known species, A. quiriquinensis, dates back to the late 1950s, when Argentinian paleontologist Rodolfo Casamiquela discovered a partial skeleton in Las Tablas Bay, located north of Quiriquina Island, Chile. The specimen, cataloged as SGO.PV.260, was found in Maastrichtian beds of the Quiriquina Formation, and first consisted of five vertebrae, a humerus fragment, and the distal end of a limb. The remarkable hardness of the rocks containing the fossils made subsequent preparation difficult. After preparation, the fossil contains all four limbs, a posterior portion of the neck, most of the trunk, and a complete tail. It is interpreted as a juvenile. This specimen was first analyzed in 2012 and identified as an indeterminate aristonectine,[17] before being assigned to the genus Aristonectes in 2013.[18] In 2014, the specimen was referred to as A. quiriquinensis.[12]
In 2001, Chilean paleontologist Mario E. Suárez collected a partial skull, mandibular fragments and 12 anterior cervical vertebrae from a beach near Cocholgüe, located north of Tomé. This material was described in the following year and attributed to Aristonectes, but identification as a separate species was then impossible due to lack of preparation.[19] In early 2009, a second excavation was carried out independently at the same site by a team from the University of Concepción of Chile and the University of Heidelberg of Germany. This excavation recovered 119 blocks of sandstone, most of them with bone material, but some were damaged due to the degradation of the bones by the periodic immersion of sea water which transformed the most delicate parts into brittle surfaces. Additionally, several anatomical contacts were lost as blocks of sandstone were cut from the beach using a rock saw at low tide. The precise location of the find and the taphonomic distribution of the fossils show that they belong to the same specimen that was discovered in 2001. The bones were sent to the Chilean National Museum of Natural History for the preparation and scientific analysis. The completely prepared fossil consists of the skull, the axis and the atlas, 12 anterior cervical vertebrae, 23 middle to posterior cervical vertebrae, most of the trunk, the two almost complete forelimbs and a significant part of the right hind limb. The specimen was likely a young adult. After comparison with A. parvidens, the specimen, which is cataloged as SGO.PV.957, became the holotype of the new species A. quiriquinensis. The specific epithet is a reference to the formation of Quiriquina, which is the type locality of this taxon.[12]
In 2015, two other specimens from Quiriquina Island were referred to this species. These are composed of a complete left femur and a proximal part of a humerus, cataloged respectively as SGO.PV.135 and SGO.PV.169, which were previously referred to the now dubious genus Mauisaurus.[20] In 2018, the holotype specimen was redescribed after further preparation. In the same publication, another specimen, SGO.PV.94, consisting of a series of anterior caudal vertebrae, was also referred to A. quiriquensis.[21]
Description
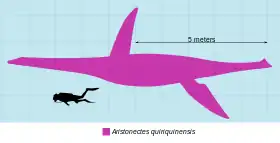
Aristonectes is a plesiosaur that has been particularly difficult for scientists to analyze since its official description in 1941, because many missing parts of its fossils prevented a concrete analysis of its anatomy,[15][16] to the point of being declared as an incertae sedis.[14] Although the fossils are incomplete, more recent research carried out on A. parvidens and the discovery of the second species A. quiriquinensis have made it possible to redescribe its anatomy and classify it among the elasmosaurids.[9][11][12][21] However, some morphological differences between A. parvidens and A. quiriquinensis cannot be drawn, as many anatomical regions are not known between the two species in their respective fossils.[12][11] Aristonectes is a large representative of the plesiosauroids, the holotype of A. quiriquinensis having an estimated size of about or more than 10 metres (33 ft) long for a body mass of 4 metric tons (4.4 short tons).[21][22] A referred specimen to Aristonectes discovered in the López de Bertodano Formation in Antarctica, cataloged as MLP 89-III-3-1, is considered to be one of the largest and heaviest plesiosaurs identified to date, having an estimated size between 11–11.9 metres (36–39 ft) long for a body mass of 10.7–13.5 metric tons (11.8–14.9 short tons).[23][lower-alpha 1]
Skull
In the two known species, the skull of Aristonectes is flattened and ogive-shaped.[9][11][12][21] The size of the skull varies very slightly between the two species, that of the holotype of A. quiriquinensis having a proposed size between 65–70 cm (26–28 in), while from the holotype of A. parvidens measures 73.5 cm (28.9 in).[12] In A. quiriquinensis the squamosal bones extend well posterior to the occipital condyle. The pterygoid bones are large and project posterior to the neurocranium. The posterior extension of the pterygoid bones surrounding the axis and the atlas, this reduces the space between the skull and the vertebrae, and would therefore limit lateral movement at their joints.[21] Around 13 teeth were present in the premaxilla and 50 teeth present in the maxilla.[12][11] Among the unique characters of the skull of A. quiriquinensis is the presence of a mental boss (a ridge that appears in the mandibular symphysis), a feature not observed in the holotype of A. parvidens.[12]
The mandibular symphysis of Aristonectes is short.[6][9][11][12][21] The dentary bones, although being partially preserved in both species, would have a total of 50 teeth in A. quiriquinensis and 63 to more in A. parvidens according to estimates.[9][12][24][11][lower-alpha 2] The mandibular symphysis of the dentary bones in A. quiriquinensis is comparatively thicker and lacks the deep groove seen in ventral view in A. parvidens. The most posterior teeth of A. parvidens were about 60 mm (2.4 in) rostrally from the coronoid process. In A. quiriquinensis, there are no tooth sockets present although 130 mm (5.1 in) of the mandible is preserved rostral to the coronoid process. Therefore, it is likely that A. quiriquinensis would have around fewer teeth than A. parvidens. In A. parvidens, the labial contact between the angular bone and the surangular, anterior to the glenoid fossa, bears a deep cavity which is not present in A. quiriquinensis.[12]
The teeth of A. quiriquinensis have an oval cross-section, being very similar to elasmosaurids of the Northern Hemisphere and Weddellian Province, as in Kaiwhekea. The length of the root of complete teeth is slightly longer than that of the crown. The teeth of A. quiriquinensis, and most likely A. parvidens,[11][lower-alpha 3] is homodont, wich means that the teeth are all of identical shape in the jaw. The largest known tooth is very thin and pointed, having a crown bearing ridges on the lingual side. The smaller tooth is a short-rooted replacement tooth, having a similar morphology to the larger one.[12] All teeth are inclined toward the front and would not fit together when the jaws are closed.[21]
Postcranial skeleton
Although some postcranial remains are known in both species, they are mainly documented in A. quiriquinensis, the holotype specimen of this latter being significantly more complete than that of A. parvidens.[12][21] In 2018, Otero and his colleagues estimate that A. quiriquinensis would have had a total of 109 vertebrae, among which would be 43 cervical vertebrae, 3 pectoral vertebrae, 24 dorsal vertebrae, 3 sacral vertebrae and 35 caudal vertebrae.[21] The anterior cervical ribs are recurved in A. parvidens and fused distally,[6] while those of A. quiriquinensis are short and lack contacts.[12] The cervical ribs and cervical neural spines of Aristonectes are curved towards the head, similar to other members of the subfamily Aristonectinae.[12][21] This morphology would have allowed for maintaining the trunk higher than the head.[21] The neural spines present on the middle of the cervical part look like blades and are enlarged distally.[12]
The dorsal vertebrae are larger and taller than the cervical vertebrae and are also taller than they are wide. Dorsal vertebrae have centra which exhibit constrictions and which have amphicelous articular surfaces.[12] Between the glenoid cavities, the pectoral girdle of A. quiriquinensis have a width of nearly one meter. The ribs appear thickened on their distal sides. The gastralia are pachyostotic and have a moderate length. The caudal vertebrae of A. quiriquinensis are wider than tall or long. The neural spines of the latter become progressively blunt and short, possessing a thick dorsal apex with a flat surface, this suggesting the probable existence of a strong ligament all along the dorsal part of the tail.[21]
The limbs of A. quiriquinensis are very long for an elasmosaurid, the best-preserved swim paddle being about 3 m (9.8 ft) long. Combined with the aforementioned width of the pectoral girdle, this indicates a total wingspan of 7 m (23 ft). In aquatic tetrapods, such proportions in the limbs are only observable in the rhomaleosaurid from the Early Jurassic Meyerasaurus and in the modern humpback whale.[21] A. parvidens would also have had similar fin size. In both species, the phalanges of their limbs have large articular facets, featuring elongated coil-shaped bones. In A. quiriquinensis the ulna is slightly shorter than the radius and the tibia is slightly wider than the fibula.[12]
Classification

Because of the wrong preparation of fossils, Aristonectes had a classification that changed many times throughout its taxonomic history.[12][11] In 1941, Cabrera classifies with doubts Aristonectes within the Elasmosauridae on the basis of observations of the rare cranial elements bringing it closer with this group,[6] a classification that will be recognized as such several decades later.[25][26][21] In 1962, Samuel P. Welles considered Aristonectes to be an indeterminate pliosaur based on its rather enigmatic anatomy for the time.[14] In 1960, Per O. Persson classified Aristonectes in the Cimoliasauridae, a proposal which he reaffirmed in 1963 on the basis of new observations on the skull reconstructed in cast.[15] In 1981, David S. Brown moved Aristonectes within the Cryptoclididae, a classification which he justified by the presence of between 6 and 15 teeth in the premaxilla.[16] From the first revision carried out on the genus in 2003, Gasparini and his colleagues reclassified again Aristonectes in the family Elasmosauridae,[9] but this classification is rejected the same year by F. Robin O'Keefe and William Wahl Jr., putting the genus in the Cimoliasauridae.[27] In 2009, O'Keefe and Hallie P. Street reviewed the validity of Cimoliasauridae, and found that this taxon is a junior synonym of the Elasmosauridae. Therefore, they moved the genera Aristonectes, Kaiwhekea, Kimmerosaurus and Tatenectes into the new erected family Aristonectidae.[28] In 2011, two years later, O'Keefe and his colleagues were skeptical about the classification of Aristonectes and Kaiwhekea with Tatenectes, because their morphologies did not seem to correspond together to form a monophyletic group (natural group).[29] Otero and his colleagues erected in 2012 the subfamily Aristonectinae and classified this taxon within the Elasmosauridae. Each representative of this group is characterized by having a very enlarged skull compared to the width of the body, a moderately short neck and more than 25 teeth in the maxilla,[17] a higher number than of other elasmosaurids.[24] Along with the Elasmosaurinae, the Aristonectinae represent one of the two recognized subfamilies within the elasmosaurids.[30] In many phylogenetic analyses, Aristonectes is the most derived genus within the Aristonectinae.[25][26][30][31]
The following cladogram is modified from Otero & Acuña, (2020):[31]
| Elasmosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Gastroliths
As with most other plesiosaur fossils, gastroliths have been found in some specimens of Aristonectes. Among these are 5 elements interpreted as gastroliths who have been discovered during the second exhumation of the holotype of A. quiriquinensis,[12] and over 793 listed in the largest known specimen of the genus, MLP 89-III-3-1, discovered in Antarctica.[32][lower-alpha 4] The function of gastroliths in plesiosaurs is still controversial and has generated several hypotheses about their function.[34] The most frequently cited hypotheses are either their use in buoyancy control or in the digestive process, the latter being the most widely accepted by recent studies.[35][36][37] In the analysis carried out on the specimen MLP 89-III-3-1, the researchers conclude that Aristonectes would not ingest the gastroliths individually, but would pick them up randomly, an affirmation which is based according to their consistencies. However, as this specimen was disarticulated when it was discovered (i.e., the bones had moved out of their original anatomical position), it is likely that not all of its gastrolithes have been found. The gastroliths do not indicate whether they were used in this genus for digestion.[32]
Feeding
A. quiriquinensis has a morphology of the swimming paddles and the tail close to various modern cetaceans, and postcranial characteristics which would limit the lateral movement of the head. In addition, the animal's skull has an enlarged mouth that would have allowed the engulfment of a large volume of water. This indicates that A. quiriquinensis fed in benthic zones, mixing prey and sediment at the same time. This type of feeding pattern is also documented in modern gray whales. The presence of decapod crustaceans and fishes within the Quiriquina Formation further reinforces this assertion.[21]
Growth
After the discovery of A. quiriquinensis in 2014, several specimens were subsequently referred to this latter. In a study published in 2015 by Otero and his colleagues, it was noted that during their growth, the humerus and femur go from a flat capitulum in juveniles to hemispherical shaped heads of the two bones at adulthood.[20]
Paleoecology

Aristonectes is known from various geological formations dating from the Maastrichtian of Patagonia and Antarctica. It is more precisely known from Quiriquina[12][21] and Dorotea formations in Chile,[38] of the Lefipán Formation in Argentina[11] and in the López de Bertodano Formation in Antarctica.[39][23] The presence of Aristonectes within these formations shows that it would have been endemic in the Weddellian Province, a geographical area that appeared after the isolation of Antarctica.[38] This also indicates that the genus could have migrated regularly between Antarctica and Patagonia, like many current cetaceans.[21] Many bony fishes, cartilaginous fishes, crustaceans and molluscs are known from most localities from which Aristonectes is listed.[40][38][21][41]
The diversification of plesiosaurs seems to vary from formation to formation. In the Lefipán Formation, only Aristonectes is known,[9][11] while in the Quiriquina Formation it was contemporary with its close relative Wunyelfia.[31] In Dorotea and López de Bertodano formations, in addition to Aristonectes, there are many indeterminate elasmosaurids.[38][39] Exclusively in the López de Bertodano Formation, numerous genera of contemporary mosasaurs have been identified. These include Kaikaifilu, Moanasaurus, Mosasaurus, Liodon and Plioplatecarpus, although the validity of some of these genera is disputed as being primarily based on isolated teeth.[42][43] Some of these mosasaurs could have attacked the contemporary plesiosaurs of the formation, including Aristonectes.[44]
Notes
- The body mass estimate per O'Gorman and his colleagues is based on their assumption that Cryptoclidus and Aristonectes would have had similar body proportions.[23]
- In the original 1941 description by Cabrera, the number of teeth in the lower jaw of A. parvidens was estimated at 58,[6] before being proposed at between 60 and 65 in the first revision of the genus carried out by Gasparini et al., (2003).[9] In 2016, O'Gorman reduced this estimate slightly to at least 63 teeth, but did not rule out the possibility that the true number could have been between 63 and 65.[11]
- Only 9 teeth are known in the fossils of A. quiriquinensis,[12][21] while none are known from A. parvidens.[9][11]
- Before the study concerning this specimen published in 2014, only 560 gastroliths were found.[33][12]
References
- José P. O'Gorman; Zulma Gasparini; Leonardo Salgado (2013). "Postcranial morphology of Aristonectes (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of Patagonia and Antarctica". Antarctic Science. 25 (1): 71–82. Bibcode:2013AntSc..25...71O. doi:10.1017/S0954102012000673. S2CID 128417881.
- Claude Gay (1848). "Reptiles Fosiles". Zoologia, Vol. 2. Historia Física y Política de Chile [Physical and Political History of Chile] (in Spanish). Vol. 2. Paris: Imprenta Maulde y Renou. pp. 130–136.
- Richard Lydekker (1889). Catalogue of the fossil Reptilia and Amphibia in the British Museum. Part II. London: British Museum. p. 222.
- Wilhelm Deecke (1895). "Ueber Saurierreste aus den Quiriquina−Schichten" [Concerning Dinosaur Remains from the Quiriquina Strata]. Beiträge zur Geologie und Palaeontologie von Südamerika (in German). 14: 32–63.
- Pavel Pravoslavlev (1918). "Геологическое распространенiе эласмозавровъ" [Geological distribution of Elasmosaurus]. Bulletin of the Russian Academy of Sciences. VI (in Russian). 12 (17): 1955–1978. Archived from the original on 2017-12-01. Retrieved 2017-11-18.
- Ángel Cabrera (1941). "Un Plesiosaurio nuevo del Cretáceo del Chubut" [A new Plesiosaur of the Cretaceous of Chubut]. Revista del Museo de La Plata (in Spanish). 2 (8): 113–130.
- Edwin H. Colbert (1949). "A new Cretaceous plesiosaur from Venezuela". American Museum Novitates (1420): 1–22. CiteSeerX 10.1.1.1033.3285. hdl:2246/2347.
- Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers (2010). "Presence of Mauisaurus in the Maastrichtian (Late Cretaceous) of central Chile". Acta Palaeontologica Polonica. 55 (2): 361–364. doi:10.4202/app.2009.0065. S2CID 130819450.
- Zulma Gasparini; Nathalie Bardet; James E. Martin; Marta Fernandez (2003). "The elasmosaurid plesiosaur Aristonectes Cabrera from the latest Cretaceous of South America and Antarctica". Journal of Vertebrate Paleontology. 23 (1): 104–115. doi:10.1671/0272-4634(2003)23[104:TEPACF]2.0.CO;2. JSTOR 4524298. S2CID 85897767.
- José P. O'Gorman (2013). Plesiosaurios del Cretácico Superior de Patagonia y la península Antártica [Plesiosaurs from the Upper Cretaceous of Patagonia and the Antarctic Peninsula] (PhD) (in Spanish). Universidad Nacional de La Plata. hdl:10915/26140. S2CID 127759354.
- José P. O'Gorman (2016). "New Insights on the Aristonectes parvidens (Plesiosauria, Elasmosauridae) Holotype: News on an Old Specimen". Ameghiniana. 52 (4): 397–417. doi:10.5710/AMGH.24.11.2015.2921. S2CID 89238379.
- Rodrigo A. Otero; Sergio Soto-Acuña; Frank Robin O'Keefe; José P. O’Gorman; Wolfgang Stinnesbeck; Mario E. Suárez; David Rubilar-Rogers; Christian Salazar; Luis Arturo Quinzio-Sinn (2014). "Aristonectes quiriquinensis, sp. nov., a new highly derived elasmosaurid from the upper Maastrichtian of central Chile". Journal of Vertebrate Paleontology. 34 (1): 100–125. Bibcode:2014JVPal..34..100O. doi:10.1080/02724634.2013.780953. hdl:10915/128448. JSTOR 24523254. S2CID 84729992.
- Ben Creisler (2012). "Ben Creisler's Plesiosaur Pronunciation Guide". Oceans of Kansas. Retrieved 2021-06-26.
- Samuel P. Welles (1962). "A new species of elasmosaur from the Aptian of Colombia and a review of the Cretaceous plesiosaurs" (PDF). University of California Publications in Geological Sciences. 44 (1): 1–96.
- Per Ove Persson (1963). "A revision of the classification of the Plesiosauria with a synopsis of the stratigraphical and geographical distribution of the group" (PDF). Lunds Universitets Arsskrift. 59 (1): 1–59.
- David S. Brown (1981). "The English Late Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria". Bulletin of the British Museum (Natural History), Geology. 35 (4): 253–347.
- Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers (2012). "A postcranial skeleton of an elasmosaurid plesiosaur from the Maastrichtian of central Chile, with comments on the affinities of Late Cretaceous plesiosauroids from the Weddellian Biogeographic Province". Cretaceous Research. 37: 89–99. Bibcode:2012CrRes..37...89O. doi:10.1016/j.cretres.2012.03.010. S2CID 129841690.
- Rodrigo A. Otero; José P. O'Gorman (2013). "Identification of the first postcranial skeleton of Aristonectes Cabrera (Plesiosauroidea, Elasmosauridae) from the upper Maastrichtian of the south-eastern Pacific, based on a bivariate graphic analysis". Cretaceous Research. 41: 86–89. Bibcode:2013CrRes..41...86O. doi:10.1016/j.cretres.2012.11.001. S2CID 53485462.
- Mario E. Suárez; Omar Fritis (2002). "Nuevo registro de Aristonectes sp. (Plesiosauroidea incertae sedis) del Cretácico Tardío de la Formación Quiriquina, Cocholgüe, Chile" [New record of Aristonectes sp. (Plesiosauroidea incertae sedis) from the Late Cretaceous of the Quiriquina Formation, Cocholgüe, Chile]. Boletín de la Sociedad de Biología de Concepción (in Spanish). 73: 87–93.
- Rodrigo A. Oterio; José P. O'Gorman; Norton Hiller (2015). "Reassessment of the upper Maastrichtian material from Chile referred to Mauisaurus Hector, 1874 (Plesiosauroidea: Elasmosauridae) and the taxonomical value of the hemispherical propodial head among austral elasmosaurids". New Zealand Journal of Geology and Geophysics. 58 (3): 252–261. doi:10.1080/00288306.2015.1037775. S2CID 129018714.
- Rodrigo A. Otero; Sergio Soto-Acuña; Frank R. O'keefe (2018). "Osteology of Aristonectes quiriquinensis (Elasmosauridae, Aristonectinae) from the upper Maastrichtian of central Chile". Journal of Vertebrate Paleontology. 38 (1): e1408638. Bibcode:2018JVPal..38E8638O. doi:10.1080/02724634.2017.1408638. JSTOR 44865847. S2CID 90977078.
- Gregory S. Paul (2022). The Princeton Field Guide to Mesozoic Sea Reptiles. Princeton University Press. p. 119. doi:10.2307/j.ctv2hnkc6h. ISBN 978-0-691-19380-9. S2CID 251553177.
- José P. O'Gorman; Sergio N. Santillana; Rodrigo A. Otero; Marcelo A. Reguero (2019). "A giant elasmosaurid (Sauropterygia; Plesiosauria) from Antarctica: New information on elasmosaurid body size diversity and aristonectine evolutionary scenarios" (PDF). Cretaceous Research. 102: 37–58. Bibcode:2019CrRes.102...37O. doi:10.1016/j.cretres.2019.05.004. S2CID 181725020.
- Sven Sachs; Benjamin P. Kear (2015). "Fossil Focus: Elasmosaurs". www.palaeontologyonline.com. Palaeontology Online. pp. 1–8. Archived from the original on February 1, 2018. Retrieved January 18, 2018.
- R. Araújo; Michael J. Polcyn; Johan Lindgren; L. L. Jacobs; A. S. Schulp; O. Mateus; A. Olimpio Gonçalves; M-L. Morais (2015). "New aristonectine elasmosaurid plesiosaur specimens from the Early Maastrichtian of Angola and comments on paedomorphism in plesiosaurs" (PDF). Netherlands Journal of Geosciences. 94 (1): 93–108. doi:10.1017/NJG.2014.43. S2CID 55793835. Archived from the original (PDF) on 2020-07-11.
- Rodrigo A. Otero (2016). "Taxonomic reassessment of Hydralmosaurus as Styxosaurus: new insights on the elasmosaurid neck evolution throughout the Cretaceous". PeerJ. 4: e1777. doi:10.7717/peerj.1777. PMC 4806632. PMID 27019781.
- F. Robin O’Keefe; William Wahl Jr. (2003). "Preliminary report on the osteology and relationships of a new aberrant cryptocleidoid plesiosaur from the Sundance Formation, Wyoming". Paludicola. 4 (2): 48–68.
- F. Robin O'Keefe; Hallie P. Street (2009). "Osteology Of The cryptoclidoid plesiosaur Tatenectes laramiensis, with comments on the taxonomic status of the Cimoliasauridae" (PDF). Journal of Vertebrate Paleontology. 29 (1): 48–57. doi:10.1671/039.029.0118. S2CID 31924376.
- F. Robin O'Keefe; Hallie P. Street; Benjamin C. Wilhelm; Courtney D. Richards; Helen Zhu (2011). "A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape among plesiosaurs". Journal of Vertebrate Paleontology. 31 (2): 330–339. doi:10.1080/02724634.2011.550365. S2CID 54662150.
- José P. O'Gorman (2020). "Elasmosaurid phylogeny and paleobiogeography, with a reappraisal of Aphrosaurus furlongi from the Maastrichtian of the Moreno Formation". Journal of Vertebrate Paleontology. 39 (5): e1692025. doi:10.1080/02724634.2019.1692025. S2CID 215756238.
- Rodrigo A. Otero; Sergio Soto-Acuña (2020). "Wunyelfia maulensis gen. et sp. nov., a new basal aristonectine (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of central Chile". Cretaceous Research. 188: 104651. Bibcode:2021CrRes.11804651O. doi:10.1016/j.cretres.2020.104651. S2CID 224975253.
- José P. O’Gorman; Eduardo B. Olivero; Sergio Santillana; Michael J. Everhart; Marcelo Reguero (2014). "Gastroliths associated with an Aristonectes specimen (Plesiosauria, Elasmosauridae), López de Bertodano Formation (upper Maastrichtian) Seymour Island (Is. Marambio), Antarctic Peninsula". Cretaceous Research. 50: 228–237. Bibcode:2014CrRes..50..228O. doi:10.1016/j.cretres.2014.03.011. hdl:11336/7400. S2CID 129616515.
- José P. O’Gorman; Sergio Santillana; Marcelo Reguero; J. J. Moly (2012), "Primer registro de gastrolitos asociados a un especimen de Aristonectes sp. (Plesiosauria, Elasmosauridae), Isla Seymour (Is. Marambio), Antartida" [First record of gastroliths associated with a specimen of Aristonectes sp. (Plesiosauria, Elasmosauridae), Seymour Island (Is. Marambio), Antarctica], in Lepe, Mario; Aravena, Juan Carlos; Villa-Martínez, Rodrigo (eds.), Abriendo Ventanas al Pasado. Libro de Resúmenes del III Simposio - Paleontología en Chile [Opening Windows to the Past. Book of Abstracts of the III Symposium - Paleontology in Chile] (in Spanish), Centro de Estudios del Cuaternario y Antártica del Instituto Antártico Chileno, pp. 121–123, OCLC 1318661175
- Oliver Wings (2007). "A review of gastrolith function with implications for fossil vertebrates and a revised classification". Acta Palaeontologica Polonica. 52 (1): 1–16.
- Michael J. Everhart (2000). "Gastroliths Associated with Plesiosaur Remains in the Sharon Springs Member of the Pierre Shale (Late Cretaceous), Western Kansas". Transactions of the Kansas Academy of Science. 103 (1–2): 64–75. doi:10.2307/3627940. JSTOR 3627940. S2CID 88446437.
- David J. Cicimurri; Michael J. Everhart (2001). "An Elasmosaur with Stomach Contents and Gastroliths from the Pierre Shale (Late Cretaceous) of Kansas". Transactions of the Kansas Academy of Science. 104 (3–4): 129–143. doi:10.1660/0022-8443(2001)104[0129:AEWSCA]2.0.CO;2. JSTOR 3627807. S2CID 86037286.
- Ignacio A. Cerda; Leonardo Salgado (2008). "Gastrolitos en un plesiosaurio (Sauropterygia) de la Formación Allen (Campaniano-Maastrichtiano), provincia de Río Negro, Patagonia, Argentina". Ameghiniana (in Spanish). 45 (8): 529–536. ISSN 0002-7014.
- Rodrigo A. Otero; Sergio Soto-Acuña; Christian Salazar; José L. Oyarzún (2015). "New elasmosaurids (Sauropterygia, Plesiosauria) from the Late Cretaceous of the Magallanes Basin, Chilean Patagonia: Evidence of a faunal turnover during the Maastrichtian along the Weddellian Biogeographic Province". Andean Geology. 42 (2): 237–267. doi:10.5027/andgeoV42n1-a05. S2CID 54994060.
- José P. O'Gorman; Karen M. Panzeri; Marta S. Fernández; Sergio Santillana; Juan J. Moly; Marcelo Reguero (2018). "A new elasmosaurid from the upper Maastrichtian López de Bertodano Formation: new data on weddellonectian diversity". Alcheringa: An Australasian Journal of Palaeontology. 42 (4): 575–586. doi:10.1080/03115518.2017.1339233. S2CID 134265841.
- Wolfgang Stinnesbeck (1986). "Zu den faunistischen und palökologischen Verhältnissen in der Quiriquina Formation (Maastrichtium) Zentral-Chiles" [On the faunistic and paleoecological conditions in the Quiriquina Formation (Maastrichtian) of central Chile]. Palaeontographica Abteilung A (in German). 194 (4–6): 99–237.
- Alberto L. Cione; Sergio Santillana; Soledad Gouiric-Cavalli; Carolina Acosta Hospitaleche; Javier N. Gelfo; Guillermo M. Lopez; Marcelo Reguero (2018). "Before and after the K/Pg extinction in West Antarctica: New marine fish records from Marambio (Seymour) Island". Cretaceous Research. 85: 250–265. doi:10.1016/j.cretres.2018.01.004. hdl:10915/147537. S2CID 133767014.
- Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers; Carolina S. Gutstein (2017). "Kaikaifilu hervei gen. et sp. nov., a new large mosasaur (Squamata, Mosasauridae) from the upper Maastrichtian of Antarctica". Cretaceous Research. 70: 209–225. doi:10.1016/j.cretres.2016.11.002. S2CID 133320233.
- Pablo Gonzalez Ruiz; Marta S. Fernandez; Marianella Talevi; Juan M. Leardi; Marcelo A. Reguero (2019). "A new Plotosaurini mosasaur skull from the upper Maastrichtian of Antarctica. Plotosaurini paleogeographic occurrences". Cretaceous Research. 103: 104166. doi:10.1016/j.cretres.2019.06.012. hdl:11336/125124. S2CID 198418273.
- University of Chile (2016-11-07). "A giant predatory lizard swam in Antarctic seas near the end of the dinosaur age". ScienceDaily.
External links
- Aristonectes information and photos, The Plesiosaur Directory
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||






.png.webp)