BNIP3
BCL2/adenovirus E1B 19 kDa protein-interacting protein 3 is a protein found in humans that is encoded by the BNIP3 gene.[5]






BNIP3 is a member of the apoptotic Bcl-2 protein family. It can induce cell death while also assisting with cell survival. Like many of the Bcl-2 family proteins, BNIP3 modulates the permeability state of the outer mitochondrial membrane by forming homo- and hetero-oligomers inside the membrane.[6] Upregulation results in a decrease in mitochondrial potential, an increase in reactive oxygen species, mitochondrial swelling and fission, and an increase in mitochondrial turnover via autophagy.[7] Sequence similarity with Bcl-2 family members was not detected. Humans and other animals (Drosophila, Caenorhabditis), as well as lower eukaryotes (Dictyostelium, Trypanosoma, Cryptosporidium, Paramecium) encode several BNIP3 paralogues including the human NIP3L, which induces apoptosis by interacting with viral and cellular anti-apoptosis proteins.
Structure
The right-handed parallel helix-helix structure of the domain with a hydrogen bond-rich His-Ser node in the middle of the membrane, accessibility of the node for water, and continuous hydrophilic track across the membrane suggest that the domain can provide an ion-conducting pathway through the membrane. Incorporation of the BNIP3 transmembrane domain into an artificial lipid bilayer resulted in a pH-dependent conductivity increase. Necrosis-like cell death induced by BNIP3 may be related to this activity.[8]
Function
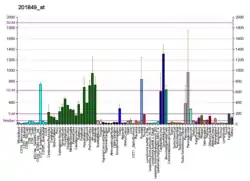
BNIP3 interacts with the E1B 19 kDa protein which is responsible for the protection of virally induced cell death, as well as E1B 19 kDa-like sequences of BCL2, also an apoptotic protector. This gene contains a BH3 domain and a transmembrane domain, which have been associated with pro-apoptotic function. The dimeric mitochondrial protein encoded by this gene is known to induce apoptosis, even in the presence of BCL2.[9] Change of BNIP3 expression along other members of the Bcl-2 family measured by qPCR captures important characteristics of malignant transformation, and are defined as markers of resistance toward cell death, a key hallmark of cancer.[10]
Transport reaction
The reaction catalyzed by BNIP3 is:
- small molecules (out) ⇌ small molecules (in)
Autophagy
Autophagy is important for recycling cellular contents and prolonging cell life. Hanna et al. show that BNIP3 and LC3 interact to remove endoplasmic reticulum and mitochondria.[11] When inactive BNIP3 is activated on the membrane of the mitochondria, they form homodimers where LC3 can bind to the LC3-interacting region (LIR) motif on BNIP3 and facilitates the formation of an autophagosome.[11][12] Interestingly, when disrupting BNIP3 and LC3 interaction, researchers found that autophagy was reduced but not completely erased. This suggests that BNIP3 is not the only receptor on the mitochondria and ER to promote autophagy.[11]
This relationship between autophagy and BNIP3 is widely supported in many studies. In ceramide- and arsenic trioxide- treated malignant glioma cells, increased BNIP3 expression led to mitochondrial depolarization and autophagy.[13][14]
Autophagic cell death
Increased expression of BNIP3 has been shown to induce cell death in different ways in multiple cell lines. BNIP3 can induce classical apoptosis through cytochrome c and caspase activation in some cells, while in others, cells have undergone autophagic cell death, occurring in the absence of apaf-1, caspase-1 or caspase 3, and without cytochrome c release.[7][15]
However, it still remains unclear if cell death is from excess autophagy itself or another mechanism. Cell death through excessive autophagy has only been shown experimentally and not in mammalian in vivo models. Kroemer and Levine believe that this name is a misnomer because cell death usually occurs with autophagy rather than by autophagy.[16]
NK cell memory formation
The innate immune system is generally not known to exhibit memory traits, but emerging research has proven otherwise. In 2017, O’Sullivan et al. found that BNIP3 and BNIP3L play a necessary role in promoting NK cell memory formation.[17] Expression of BNIP3 in NK cells is lowered upon viral infection as NK cell proliferation occurs but returns to its basal amounts by day 14 and through the contraction phase.[17] By using BNIP3-knockout mice, they found a significant decrease in surviving NK cells suggesting they are important to maintain survival of NK memory cells.[17] Additionally, by tracking mitochondria amounts and quality, they found that BNIP3 is necessary for clearing dysfunctional mitochondria with low membrane potential and reducing the build up of ROS to promote cell survival.[17] BNIP3L was also tested and was found to play a nonredundant role in cell survival.[17]
Activities in the mitochondrial membrane
Integration
Various stimuli like decreased intracellular pH, increased cytosolic calcium concentrations, and other toxic stimuli can induce BNIP3 integration into the outer mitochondrial membrane (OMM).[18] When integrated, its N-terminus remains in the cytoplasm while it stays anchored to the OMM via its C-terminal transmembrane domain (TMD).[19] The TMD is essential for targeting BNIP3 to the mitochondria, homodimerization, and pro-apoptotic function.[20][21][22] Its deletion results in the inability to induce autophagy.[11] Once integrated in the OMM, BNIP3 exists as an inactive monomer until activated.
Activation
Upon activation, BNIP3 can form heterodimers with BCL2 and BCL-XL and bind to itself.[15] Various conditions have been shown to induce activation and upregulation. Hypoxia has been shown to induce transcriptional upregulation of BNIP3 through an HIF1-dependent pathway in a p53-independent manner in HeLa cells, human skeletal muscle cells, and adult rat cardiomyocytes.[23]
Using BNIP3 phosphomimetics in HEK 293 cells, researchers found that phosphorylation of BNIP3's C-terminus is necessary to prevent mitochondrial damage and promote cell survival by allowing a significant amount of autophagy to occur without the induction of cell death.[7] Factors like cAMP and cGMP levels, calcium availability and growth factors like IGF and EGF can affect this kinase activity.[7]
References
- GRCh38: Ensembl release 89: ENSG00000176171 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000078566 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Boyd JM, Malstrom S, Subramanian T, Venkatesh LK, Schaeper U, Elangovan B, et al. (October 1994). "Adenovirus E1B 19 kDa and Bcl-2 proteins interact with a common set of cellular proteins". Cell. 79 (2): 341–51. doi:10.1016/0092-8674(94)90202-X. PMID 7954800. S2CID 38609845.
- Sassone J, Colciago C, Marchi P, Ascardi C, Alberti L, Di Pardo A, et al. (January 2010). "Mutant Huntingtin induces activation of the Bcl-2/adenovirus E1B 19-kDa interacting protein (BNip3)". Cell Death & Disease. 1 (1): e7. doi:10.1038/cddis.2009.6. PMC 3032515. PMID 21364626.
- Liu KE, Frazier WA (2015-06-23). "Phosphorylation of the BNIP3 C-Terminus Inhibits Mitochondrial Damage and Cell Death without Blocking Autophagy". PLOS ONE. 10 (6): e0129667. Bibcode:2015PLoSO..1029667L. doi:10.1371/journal.pone.0129667. PMC 4477977. PMID 26102349.
- Bocharov EV, Pustovalova YE, Pavlov KV, Volynsky PE, Goncharuk MV, Ermolyuk YS, et al. (June 2007). "Unique dimeric structure of BNip3 transmembrane domain suggests membrane permeabilization as a cell death trigger". The Journal of Biological Chemistry. 282 (22): 16256–66. doi:10.1074/jbc.M701745200. PMID 17412696.
- "Entrez Gene: BNIP3 BCL2/adenovirus E1B 19kDa interacting protein 3".
- Menyhárt O, Harami-Papp H, Sukumar S, Schäfer R, Magnani L, de Barrios O, Győrffy B (December 2016). "Guidelines for the selection of functional assays to evaluate the hallmarks of cancer". Biochimica et Biophysica Acta (BBA) - Reviews on Cancer. 1866 (2): 300–319. doi:10.1016/j.bbcan.2016.10.002. PMID 27742530.
- Hanna RA, Quinsay MN, Orogo AM, Giang K, Rikka S, Gustafsson ÅB (June 2012). "Microtubule-associated protein 1 light chain 3 (LC3) interacts with Bnip3 protein to selectively remove endoplasmic reticulum and mitochondria via autophagy". The Journal of Biological Chemistry. 287 (23): 19094–104. doi:10.1074/jbc.M111.322933. PMC 3365942. PMID 22505714.
- Birgisdottir ÅB, Lamark T, Johansen T (August 2013). "The LIR motif - crucial for selective autophagy". Journal of Cell Science. 126 (Pt 15): 3237–47. doi:10.1242/jcs.126128. PMID 23908376.
- Kanzawa T, Zhang L, Xiao L, Germano IM, Kondo Y, Kondo S (February 2005). "Arsenic trioxide induces autophagic cell death in malignant glioma cells by upregulation of mitochondrial cell death protein BNIP3". Oncogene. 24 (6): 980–91. doi:10.1038/sj.onc.1208095. PMID 15592527.
- Daido S, Kanzawa T, Yamamoto A, Takeuchi H, Kondo Y, Kondo S (June 2004). "Pivotal role of the cell death factor BNIP3 in ceramide-induced autophagic cell death in malignant glioma cells". Cancer Research. 64 (12): 4286–93. doi:10.1158/0008-5472.CAN-03-3084. PMID 15205343.
- Zhang J, Ney PA (July 2009). "Role of BNIP3 and NIX in cell death, autophagy, and mitophagy". Cell Death and Differentiation. 16 (7): 939–46. doi:10.1038/cdd.2009.16. PMC 2768230. PMID 19229244.
- Kroemer G, Levine B (December 2008). "Autophagic cell death: the story of a misnomer". Nature Reviews. Molecular Cell Biology. 9 (12): 1004–10. doi:10.1038/nrm2529. PMC 2727358. PMID 18971948.
- O'Sullivan TE, Johnson LR, Kang HH, Sun JC (August 2015). "BNIP3- and BNIP3L-Mediated Mitophagy Promotes the Generation of Natural Killer Cell Memory". Immunity. 43 (2): 331–42. doi:10.1016/j.immuni.2015.07.012. PMC 5737626. PMID 26253785.
- Graham RM, Thompson JW, Wei J, Bishopric NH, Webster KA (September 2007). "Regulation of Bnip3 death pathways by calcium, phosphorylation, and hypoxia-reoxygenation". Antioxidants & Redox Signaling. 9 (9): 1309–15. doi:10.1089/ars.2007.1726. PMID 17638546.
- Vande Velde C, Cizeau J, Dubik D, Alimonti J, Brown T, Israels S, et al. (August 2000). "BNIP3 and genetic control of necrosis-like cell death through the mitochondrial permeability transition pore". Molecular and Cellular Biology. 20 (15): 5454–68. doi:10.1128/mcb.20.15.5454-5468.2000. PMC 85997. PMID 10891486.
- Ray R, Chen G, Vande Velde C, Cizeau J, Park JH, Reed JC, et al. (January 2000). "BNIP3 heterodimerizes with Bcl-2/Bcl-X(L) and induces cell death independent of a Bcl-2 homology 3 (BH3) domain at both mitochondrial and nonmitochondrial sites". The Journal of Biological Chemistry. 275 (2): 1439–48. doi:10.1074/jbc.275.2.1439. PMID 10625696.
- Chen G, Ray R, Dubik D, Shi L, Cizeau J, Bleackley RC, et al. (December 1997). "The E1B 19K/Bcl-2-binding protein Nip3 is a dimeric mitochondrial protein that activates apoptosis". The Journal of Experimental Medicine. 186 (12): 1975–83. doi:10.1084/jem.186.12.1975. PMC 2199165. PMID 9396766.
- Kubli DA, Quinsay MN, Huang C, Lee Y, Gustafsson AB (November 2008). "Bnip3 functions as a mitochondrial sensor of oxidative stress during myocardial ischemia and reperfusion". American Journal of Physiology. Heart and Circulatory Physiology. 295 (5): H2025-31. doi:10.1152/ajpheart.00552.2008. PMC 2614576. PMID 18790835.
- Azad MB, Chen Y, Henson ES, Cizeau J, McMillan-Ward E, Israels SJ, Gibson SB (February 2008). "Hypoxia induces autophagic cell death in apoptosis-competent cells through a mechanism involving BNIP3". Autophagy. 4 (2): 195–204. doi:10.4161/auto.5278. PMC 3164855. PMID 18059169.
- Lamy L, Ticchioni M, Rouquette-Jazdanian AK, Samson M, Deckert M, Greenberg AH, Bernard A (June 2003). "CD47 and the 19 kDa interacting protein-3 (BNIP3) in T cell apoptosis". The Journal of Biological Chemistry. 278 (26): 23915–21. doi:10.1074/jbc.M301869200. PMID 12690108.
Further reading
- Kataoka N, Ohno M, Moda I, Shimura Y (September 1995). "Identification of the factors that interact with NCBP, an 80 kDa nuclear cap binding protein". Nucleic Acids Research. 23 (18): 3638–41. doi:10.1093/nar/23.18.3638. PMC 307259. PMID 7478990.
- Maruyama K, Sugano S (January 1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides". Gene. 138 (1–2): 171–4. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (October 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Chen G, Ray R, Dubik D, Shi L, Cizeau J, Bleackley RC, et al. (December 1997). "The E1B 19K/Bcl-2-binding protein Nip3 is a dimeric mitochondrial protein that activates apoptosis". The Journal of Experimental Medicine. 186 (12): 1975–83. doi:10.1084/jem.186.12.1975. PMC 2199165. PMID 9396766.
- Yasuda M, Theodorakis P, Subramanian T, Chinnadurai G (May 1998). "Adenovirus E1B-19K/BCL-2 interacting protein BNIP3 contains a BH3 domain and a mitochondrial targeting sequence". The Journal of Biological Chemistry. 273 (20): 12415–21. doi:10.1074/jbc.273.20.12415. PMID 9575197.
- Chen G, Cizeau J, Vande Velde C, Park JH, Bozek G, Bolton J, et al. (January 1999). "Nix and Nip3 form a subfamily of pro-apoptotic mitochondrial proteins". The Journal of Biological Chemistry. 274 (1): 7–10. doi:10.1074/jbc.274.1.7. PMID 9867803.
- Yasuda M, Han JW, Dionne CA, Boyd JM, Chinnadurai G (February 1999). "BNIP3alpha: a human homolog of mitochondrial proapoptotic protein BNIP3". Cancer Research. 59 (3): 533–7. PMID 9973195.
- Ohi N, Tokunaga A, Tsunoda H, Nakano K, Haraguchi K, Oda K, et al. (April 1999). "A novel adenovirus E1B19K-binding protein B5 inhibits apoptosis induced by Nip3 by forming a heterodimer through the C-terminal hydrophobic region". Cell Death and Differentiation. 6 (4): 314–25. doi:10.1038/sj.cdd.4400493. PMID 10381623.
- Ray R, Chen G, Vande Velde C, Cizeau J, Park JH, Reed JC, et al. (January 2000). "BNIP3 heterodimerizes with Bcl-2/Bcl-X(L) and induces cell death independent of a Bcl-2 homology 3 (BH3) domain at both mitochondrial and nonmitochondrial sites". The Journal of Biological Chemistry. 275 (2): 1439–48. doi:10.1074/jbc.275.2.1439. PMID 10625696.
- Vande Velde C, Cizeau J, Dubik D, Alimonti J, Brown T, Israels S, et al. (August 2000). "BNIP3 and genetic control of necrosis-like cell death through the mitochondrial permeability transition pore". Molecular and Cellular Biology. 20 (15): 5454–68. doi:10.1128/MCB.20.15.5454-5468.2000. PMC 85997. PMID 10891486.
- Lee SM, Li ML, Tse YC, Leung SC, Lee MM, Tsui SK, et al. (September 2002). "Paeoniae Radix, a Chinese herbal extract, inhibit hepatoma cells growth by inducing apoptosis in a p53 independent pathway". Life Sciences. 71 (19): 2267–77. doi:10.1016/S0024-3205(02)01962-8. PMID 12215374.
- Lamy L, Ticchioni M, Rouquette-Jazdanian AK, Samson M, Deckert M, Greenberg AH, Bernard A (June 2003). "CD47 and the 19 kDa interacting protein-3 (BNIP3) in T cell apoptosis". The Journal of Biological Chemistry. 278 (26): 23915–21. doi:10.1074/jbc.M301869200. PMID 12690108.
- Kothari S, Cizeau J, McMillan-Ward E, Israels SJ, Bailes M, Ens K, et al. (July 2003). "BNIP3 plays a role in hypoxic cell death in human epithelial cells that is inhibited by growth factors EGF and IGF". Oncogene. 22 (30): 4734–44. doi:10.1038/sj.onc.1206666. PMID 12879018.
- Okami J, Simeone DM, Logsdon CD (August 2004). "Silencing of the hypoxia-inducible cell death protein BNIP3 in pancreatic cancer". Cancer Research. 64 (15): 5338–46. CiteSeerX 10.1.1.326.628. doi:10.1158/0008-5472.CAN-04-0089. PMID 15289340. S2CID 16163067.
- Giatromanolaki A, Koukourakis MI, Sowter HM, Sivridis E, Gibson S, Gatter KC, Harris AL (August 2004). "BNIP3 expression is linked with hypoxia-regulated protein expression and with poor prognosis in non-small cell lung cancer". Clinical Cancer Research. 10 (16): 5566–71. doi:10.1158/1078-0432.CCR-04-0076. PMID 15328198.
- Shen XY, Zacal N, Singh G, Rainbow AJ (2005). "Alterations in mitochondrial and apoptosis-regulating gene expression in photodynamic therapy-resistant variants of HT29 colon carcinoma cells". Photochemistry and Photobiology. 81 (2): 306–13. doi:10.1562/2004-07-22-RA-242. PMID 15560738.
External links
- Human BNIP3 genome location and BNIP3 gene details page in the UCSC Genome Browser.
PDB gallery | |
|---|---|
|
As of this edit, this article uses content from "1.A.20 The BCL2/Adenovirus E1B-interacting Protein 3 (BNip3) Family", which is licensed in a way that permits reuse under the Creative Commons Attribution-ShareAlike 3.0 Unported License, but not under the GFDL. All relevant terms must be followed.