Caliciaceae
The Caliciaceae are a family of mostly lichen-forming fungi belonging to the class Lecanoromycetes in the division Ascomycota. Although the family has had its classification changed several times throughout its taxonomic history, the use of modern molecular phylogenetic methods have helped to establish its current placement in the order Caliciales. Caliciaceae contains 36 genera and about 600 species. The largest genus is Buellia, with around 300 species; there are more than a dozen genera that contain only a single species.
| Caliciaceae | |
|---|---|
 | |
| Calicium viride | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Fungi |
| Division: | Ascomycota |
| Class: | Lecanoromycetes |
| Order: | Caliciales |
| Family: | Caliciaceae Chevall. (1826) |
| Type genus | |
| Calicium Pers. (1794) | |
| Synonyms[1] | |
Most Caliciaceae grow on bark, dead wood, or rocks. Some members of this family, particularly those of the type genus, Calicium, are characterized by the presence of thin-walled and short-lasting asci (spore-bearing cells) and a mazaedium, which is an accumulation of loose, maturing spores covering the surface of the fruiting body. The resulting passive spore dispersal is relatively rare amongst the Ascomycota. The mazaedium, usually black, either sits atop a long thin stalk, or rests on (or is sometimes immersed within) the surface of the lichen substrate. Many other Caliciaceae species generate spores in an apothecium, which typically resembles a flattened black disc.
The family contains species with crustose (crusty), foliose (leafy), and, in a few instances, fruticose (shrubby) growth forms. The photobiont partner of Caliciaceae lichens is usually from the green algal genus Trebouxia. Collectively, the family has a cosmopolitan distribution, and can be found on all major land masses. Although the family is best represented in mountainous areas of temperate and tropical regions, a few hardy species can survive the harsh environment offered by Antarctica's McMurdo Dry Valleys by growing in cracks on the surface of rocks. Five Caliciaceae species are included in the International Union for Conservation of Nature's Red List of Threatened Species. Some air pollution-resistant species in the genus Pyxine have been investigated for use as biomonitors of heavy metal pollution. Several fossils of Caliciaceae found as inclusions in amber have been used to evaluate the evolutionary history of the family.
Systematics
Caliciaceae was circumscribed in 1826 by the French botanist François Fulgis Chevallier. He wrote about the family: "The Calicinees are a small group of plants, the growth of which has misled botanists. They are small parasitic fungi on the crusts of lichens, mainly on Variola and common porin, and on dead canes."[2] The type genus of the Caliciaceae is Calicium, originally circumscribed in 1794 by Christiaan Hendrik Persoon; this genus is itself typified by Calicium viride.[3] Buelliaceae and Pyxinaceae are historical families created to contain taxa that are now included in the Caliciaceae.[1]
Two other calicioid families with "Caliciaceae" in their name – Microcaliciaceae (order Pertusariales, class Lecanoromycetes) and Mycocaliciaceae (order Mycocaliciales, class Eurotiomycetes) – contain species formerly considered to be closely related to the Caliciaceae. The monogeneric Microcaliciaceae has non-lichenized, calicioid species that are parasites on lichens or free-living colonies of algae, while Mycocaliciaceae contains non-lichenized calicioid fungi lacking a mazaedium and utilising active spore dispersal.[4]

Classification
The Caliciaceae and other morphologically similar pin lichens with mazaedium-forming ascomata featuring passive spore dispersal used to be grouped together in the order Caliciales, which was for a long time considered to be a model example of a monophyletic grouping of taxa.[5] Historically, the family has also been placed by various authors in the orders Coryneliales, Helotiales, and the now-obsolete Sphaeriales, depending on which phenotypic characteristics were considered to be the most important.[6]
The Swedish lichenologist Leif Tibell spent much of his career studying calicioid lichens, and used a variety of techniques to help him understand relationships of taxa in this group,[7] including phenetic and cladistic analyses of morphology, and investigation of secondary chemistry. He suggested in 1984 that the order was polyphyletic (i.e., it did not originate from a single common ancestor) and that the main identifying features of the Caliciales had evolved independently in several different unrelated groups.[8] He restricted the order to three families (Caliciaceae, Mycocaliciaceae, and Sphinctrinaceae) that he considered to be the "core" of the group.[8][9] Preliminary phylogenetic analysis showed that Mycocaliciaceae and Sphinctrinaceae belonged to the class Eurotiomycetes, while Caliciaceae appeared to group with the order Lecanorales.[5] The proposition that mazaediate fungi are dispersed throughout the Ascomycota was confirmed later in several molecular phylogenetic studies.[5][10][4] The six genera that were included by Tibell in the Caliciaceae in his 1984 proposed classification of calicioid fungi (Acroscyphus, Calicium, Cyphelium, Texosporium, Thelomma, and Tholurna)[8] are still in the family today.[11]
Phylogenetics
The Caliciaceae and the Physciaceae were shown to be closely related in molecular studies.[12][13] Both of these families were tentatively placed in the Teloschistales as part of the suborder Physciineae.[14] Since the mazaedia-producing species of the Caliciaceae were nested within the genera Dirinaria, Pyxine, and Physcia (all once contained in the family Physciaceae), some authors treated all the members of the Caliciaceae and Physciaceae as one family, and the name Physciaceae was proposed for conservation in 2002.[15] With the appearance of additional phylogenetic studies since then, however, a two-family concept of Caliciaceae and Physciaceae has been preferred by most authorities. In 2012, the suborder Physciineae was promoted to ordinal status and the name Caliciales was resurrected.[16] In its modern circumscription, the order Caliciales contains these two families.[17] A large molecular study of the Caliciaceae-Physciaceae clade published in 2016 has helped to sort out natural relationships in this group, and more clearly define generic delimitations.[18]
The genus Culbersonia, previously classified in the Physiaceae because of its morphological resemblance to Physconia, was shown to be a member of the Caliciaceae in 2019. In phylogenetic analysis, it groups together in a clade with Pyxine and Dirinaria. These three genera are distinguished from other Caliciaceae in the appressed foliose growth form (like small leaves pressed flat against the substrate), the absence of a mazaedium, and ecologically by their predominance in the subtropics and tropics.[11]
Description

.jpg.webp)
The thallus, when present, is crustose, with a texture that is verrucose (warty), granular (grainy), or areolate. Its colour is typically grey, yellow, or greenish. Sometimes it is immersed within the outer layers of the bark of its host. The ascomata are more or less spherical or hemispherical, situated atop a long stalk in some species, but sessile in other species. When it is present, the stalk is usually melanized (blackened).[19] The form of the ascomata is either apotheciate (consisting of an apothecium) or mazaediate.[1] The apothecium consists of the asci (spore-bearing cells) and associated hymenium, and the sterile, structural, and supportive part of the apothecium, called the excipulum. In apotheciate Caliciaceae species, the apothecium is typically lecidiene (lacking a margin around the thallus, or having a margin made of the excipulum itself), rather than lecanorine (rounded with a protruding margin). The tissue structure of the excipulum is either prosoplectenchymatous (comprising long, narrow, wavy, parallel hyphae) or paraplectenchymatous (a cell arrangement where the hyphae are oriented in all directions), and is hyaline (translucent) to dark brown.[1] The genus Acroscyphus, which contains the single widespread but rare species A. sphaerophoroides, is a peculiar exception to the typical morphology of the Caliciaceae: it has a finger-like (dactyliform) thallus, immersed ascocarps on podetia (hollow stalks), and a yellow to orange medulla.[20]

The asci of Caliciaceae species are cylindrical to broadly club-shaped. They are attached to croziers, which are hook-shaped structures on cells that form at the base of the asci. The asci have thin walls and lack any internal structures at the tip. As a result, they last for only a short time before they degrade, and the ascospores within are released passively in a mazaedial mass.[19] The hamathecium (a term that refers to the tissues interspersed between the asci in the hymenium) consists of unbranched to slightly branched paraphyses that are amyloid.[1] There are usually eight spores per ascus, although sometimes this is reduced to four, or increased to 16–32 per ascus.[1] The ascospores are dark brown, with a surface that is either smooth or ornamented with remnants from the rupture of the outer wall layers. They have either zero or one septa.[19] The spores of Texosporium lichens have a unique ornamentation that is created by paraphyses that stick to the surface; this feature may help provide protection against desiccation or DNA-damaging radiation encountered when exposed in sunny habitats.[21] The conidiomata produced by Caliciaceae species are in the form of pycnidia. The conidia lack a septum, are rod-shaped (bacillar) to thread-shaped (filiform) in form (often curved), and are hyaline.[1]
The secondary chemistry of Caliciaceae species is variable. Chemicals that are commonly reported from the family are depsides (including atranorin), terpenes, depsidones (e.g., norstictic acid), and lichexanthone. Sometimes anthraquinones are present when the thallus is pigmented.[1] Acroscyphus is again an exception, as it contains secondary compounds not found in other Caliciaceae, including chloroatranorin, rugulosin, zeorin, and chrysophanic acid.[20]
Genera

.jpg.webp)


As of January 2021, Species Fungorum accepts 36 genera, 589 species, 2 subspecies, and 11 forms in the family Caliciaceae.[23] This is a list of the genera in the Caliciaceae, based on a 2020 review and summary of fungal classification by Wijayawardene and colleagues.[17] Following the genus name is the taxonomic authority (those who first circumscribed the genus; standardized author abbreviations are used), year of publication, and the number of species:

.jpg.webp)

.jpg.webp)
- Acolium (Ach.) Gray (1821)[24] – 5 spp.
- Acroscyphus Lév. (1846)[25] – 1 sp.
- Allocalicium M.Prieto & Wedin (2016)[18] – 1 sp.
- Amandinea M.Choisy ex Scheid. & M.Mayrhofer (1993)[26] – 35 spp.
- Australiaena Matzer, H.Mayrhofer & Elix (1997)[27] – 1 sp.
- Baculifera Marbach & Kalb (2000)[28] – 14 spp.
- Buellia De Not. (1846)[29] – 300 spp.
- Burrowsia Fryday & I.Medeiros (2020)[30] – 1 sp.
- Caliciella Vain. (1927)[31] – 1 sp.
- Calicium Pers. (1794)[3] – ca. 30 spp.
- Chrismofulvea Marbach (2000)[32] – 4 spp.
- Ciposia Marbach (2000)[33] – 1 sp.[note 1]
- Cratiria Marbach (2000)[36] – ca. 20 spp.
- Culbersonia Esslinger (2000)[37] – 1 sp.
- Dermatiscum Nyl. (1867)[38] – 3 spp.
- Dermiscellum Hafellner, H.Mayrhofer & Poelt (1979)[39] – 1 sp.
- Dimelaena Norman (1852)[40] – 10 spp.
- Diploicia A.Massal. (1852)[41] – ca. 12 spp.
- Diplotomma Flot. (1849) [42] – ca. 30 spp.
- Dirinaria (Tuck.) Clem. (1909)[43] – ca. 35 spp.
- Endohyalina Marbach (2000)[44] – 10 spp.
- Fluctua Marbach (2000)[45] – 1 sp.
- Gassicurtia Fée (1825)[46] – 30 spp.
- Hypoflavia Marbach (2000)[47] – 3 spp.
- Monerolechia Trevis. (1857)[48] – 4 spp.
- Orcularia (Malme) Kalb & Giralt (2011)[49] – 4 spp.
- Pseudothelomma M.Prieto & Wedin (2016)[18] – 2 spp.
- Pyxine Fr. (1825)[50] – ca. 75 spp.
- Redonia C.W.Dodge (1973)[51] – 2 spp.
- Santessonia Hale & Vobis (1978)[52] – 10 spp.
- Sculptolumina Marbach (2000)[53] – 4 spp.
- Sphinctrinopsis Woron. (1927)[54] – 1 sp.
- Stigmatochroma Marbach (2000)[55] – 9 spp.
- Tetramelas Norman (1852)[40] – 16 spp.
- Texosporium Nádv. ex Tibell & Hofsten (1968)[56] – 1 sp.
- Thelomma A.Massal. (1860)[57] – 5 spp.
- Tholurna Norman (1861)[58] – 1 sp.
- Tylophoropsis Sambo (1938)[59] – 1 sp.[note 1]
Some genera that have been until relatively recently classified as members of the Caliciaceae have had their status reassessed in light of molecular phylogenetic studies. For example, Cyphelium Ach. was synonymized with Calicium in 2016 when it was discovered that its type species grouped with the latter genus.[18] Hafellia was placed in synonymy with Buellia when a proposal to replace the conserved type of Buellia, B. disciformis, with B. aethalea,[60] was declined by the Nomenclature Committee for Fungi.[61] That proposal had attempted to retain B. disciformis as the type for Hafellia, so that the name could continue to be used for a distinctive group of crustose lichens with thickened ascospore walls that had historically been treated in Buellia.[60]
Habitat and distribution
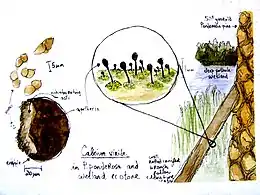
Collectively, Caliciaceae species have a cosmopolitan distribution, although they are especially predominant in temperate and tropical montane areas. Most species form lichens that grow on bark or dead wood,[19] although some grow on rocks.[1] In contrast, Texosporium is part of the biological soil crust communities found in grasslands, and grows on cow dung, soil, and detritus.[18] One rare species known only from a single locale in Sweden, Calicium episcalare, is parasitic on the common lichen Hypocenomyce scalaris.[62] Like most calicioid species, the Caliciaceae tend to avoid competition with other lichens and often grow in microhabitats neglected by other lichens. Examples include: the side of a tree not inhabited by other corticolous species, in the cracks of deep fissures in bark, or woodpecker holes. Because of this preference for neglected locales, and their generally small size, the calicioid lichens tend to be overlooked by collectors and the group in general is poorly known.[63]
Caliciaceae is one of the families whose species are most often found in mangrove forests of Calabarzon (Philippines).[64] Similar results were reported in studies on mangrove forests in India,[65] and the Gulf of Thailand,[66] where the genera Dirinaria and Pyxine were found to be amongst the most common foliose lichens. The lichens found in the mangrove ecosystem tend to be salt-tolerant and moisture sensitive.[64] Using next-generation sequencing techniques to detect the genetic signature of organisms, Caliciaceae species were identified as some of the most common endolithic lichens (crustose lichens that grow inside solid rock) found in the McMurdo Dry Valleys in Antarctica. This region features one of the most harsh environments on earth, with wide variations in temperature, extreme aridity, limited nutrients, and high levels of solar and UV radiation.[67] The photobiont partner of the Caliciaceae is usually from the green algal genus Trebouxia,[19] although Stichococcus has been recorded with Calicium.[68]
Conservation

Caliciaceae species that have been assessed for the global IUCN Red List include the following: Buellia asterella (critically endangered, 2015),[69] Buellia gypsyensis (vulnerable, 2020),[70] Buellia sharpiana (vulnerable, 2020),[71] Calicium sequoiae (endangered,[72] 2021), Texosporium sancti-jacobi (endangered, 2020),[73] and Thelomma carolinianum (endangered, 2017).[74] On the red list of China's macrofungi, Acroscyphus sphaerophoroides (endangered, 2020) is the sole representative of the Caliciaceae.[75]
Human uses
Although there are no Caliciaceae species that are known to have economic significance,[19] some of them have been exploited for scientific purposes. In a 1992 Swedish study, Allocalicium adaequatum (then a member of Calicium) was proposed as one of several bioindicator species that could be used to help assess forest continuity in boreal coniferous forests.[76] A similar study conducted in the Acadian forest ecoregion of Atlantic Canada found several Caliciaceae species appropriate for determining continuity in old-growth forests.[77]
Studies conducted in India suggest that Pyxine cocoes, a common, pollution-tolerant foliose lichen, is a candidate for biomonitoring of local air pollution.[78] It bioaccumulates toxic heavy metals that it acquires from the air and retains the pollutants in the thallus, which can then be sampled and assayed to determine their concentration.[79] Other Pyxine species in India with similar pollution-resistant properties are P. hispidula and P. subcinerea.[80][81]
Buellia frigida, a crustose species that grows on rocks in the harsh Antarctic climate, has often been used as a model organism in astrobiology research. This extremophile lichen has been exposed to conditions simulating those encountered in space and on celestial bodies like Mars, including vacuum, UV radiation, and extreme dryness. B. frigida has demonstrated resilience to these space-related stressors, making it a candidate for studying how life can adapt to and potentially survive in extraterrestrial environments.[82]
Fossil record
A fossil belonging to Calicium was described from Baltic amber dating to 55–35 million years ago (Mya).[83] This and other fossils were used to estimate the date of the split between the Caliciales (i.e., the Caliciaceae-Physciaceae clade as proposed by Gaya and colleagues in 2012[16]) and the Teloschistales. In their analysis, this occurred in the Middle Jurassic, around 171 Mya.[18] Other fossils have since been found. In 2018, three fossils assigned to Calicium were reported from European Paleogene amber.[84] A Baltic amber fungus fossil, originally collected by Robert Caspary and assigned to the genus Stilbum (family Chionosphaeraceae) in 1886, was reassessed as a member of the Caliciales in 2019, as Calicium succini. Because it is considered to be "well-preserved and reliably identifiable", it is valuable for use in evolutionary studies that estimate the divergence times of fungal lineages.[85]
Notes
- According to Index Fungorum,[34][35] the genera Ciposia and Tylophoropsis are not correct names, as their publications were predated by homonyms, the plant genera Ciposia (Myrtaceae) and Tylophoropsis (Apocynaceae). The earliest-published names have precedence according to the Principle of Priority.
References
- Jaklitsch, Walter; Baral, Hans-Otto; Lücking, Robert; Lumbsch, H. Thorsten (2016). Frey, Wolfgang (ed.). Syllabus of Plant Families: Adolf Engler's Syllabus der Pflanzenfamilien. Vol. 1/2 (13 ed.). Berlin Stuttgart: Gebr. Borntraeger Verlagsbuchhandlung, Borntraeger Science Publishers. pp. 118–119. ISBN 978-3-443-01089-8. OCLC 429208213.
- Chevallier, François-Fulgis (1826). Flore Générale des Environs de Paris (in French). Vol. 1. Paris: Chez Ferra Jeune, librairie, rue des Grands-Augustins. p. 314.
- Persoon, C.H. (1794). "Einige Bemerkungen über die Flechten: Nebst Beschreibungen einiger neue Arten aus dieser". Annalen der Botanik (in German). 7: 1–32.
- Prieto, Maria; Baloch, Elisabeth; Tehler, Anders; Wedin, Mats (2013). "Mazaedium evolution in the Ascomycota (Fungi) and the classification of mazaediate groups of formerly unclear relationship". Cladistics. 29 (3): 296–308. doi:10.1111/j.1096-0031.2012.00429.x. PMID 34818827. S2CID 85219307.
- Wedin, Mats; Tibell, Leif (1997). "Phytogeny and evolution of Caliciaceae, Mycocaliciaceae, and Sphinctrinaceae (Ascomycota), with notes on the evolution of the prototunicate ascus". Canadian Journal of Botany. 75 (8): 1236–1242. doi:10.1139/b97-837.
- Kimbrough, James W. (1970). "Current Trends in the Classification of Discomycetes". Botanical Review. 36 (2): 96–161 (see pp. 125–126. doi:10.1007/BF02858958. JSTOR 4353790. S2CID 11463562.
- Wedin, Mats. "Acharius Medallists: Leif Tibell". International Association for Lichenology. Retrieved 20 January 2021.
- Tibell, Leif (1984). A Reappraisal of the Taxonomy of Caliciales. Beihefte zur Nova Hedwigia. Vol. 79. J. Cramer. pp. 597–713.
- Tibell, Leif (1996). Caliciales. Flora Neotropica. Vol. 69. New York Botanical Garden Press. pp. 1–78. JSTOR 4393866.
- Gargas, A.; DePriest, P.; Grube, M.; Tehler, A. (1995). "Multiple origins of lichen symbioses in fungi suggested by SSU rDNA phylogeny". Science. 268 (5216): 1492–1495. Bibcode:1995Sci...268.1492G. doi:10.1126/science.7770775. PMID 7770775.
- Aptroot, André; Maphangwa, Khumbudzo Walter; Zedda, Luciana; Tekere, Memory; Alvarado, Pablo; Sipman, Harrie J. M. (2019). "The phylogenetic position of Culbersonia is in the Caliciaceae (lichenized ascomycetes)". The Lichenologist. 51 (2): 187–191. doi:10.1017/S0024282919000033. S2CID 150264979.
- Wedin, Mats; Döring, Heidi; Nordin, Anders; Tibell, Leif (2000). "Small subunit rDNA phylogeny shows the lichen families Caliciaceae and Physciaceae (Lecanorales, Ascomycotina) to form a monophyletic group". Canadian Journal of Botany. 78 (2): 246–254. doi:10.1139/b99-185.
- Wedin, Mats; Baloch, Elisabeth; Grube, Martin (2002). "Parsimony analyses of mtSSU and nITS rDNA sequences reveal the natural relationships of the lichen families Physciaceae and Caliciaceae". Taxon. 51 (4): 655–660. doi:10.2307/1555020. JSTOR 1555020.
- Miadlikowska, J.; Kauff, F.; Hofstetter, V.; Fraker, E.; Grube, M.; Hafellner, J.; et al. (2007). "New insights into classification and evolution of the Lecanoromycetes (Pezizomycotina, Ascomycota) from phylogenetic analyses of three ribosomal RNA- and two protein-coding genes". Mycologia. 98 (6): 1088–1103. doi:10.1080/15572536.2006.11832636. PMID 17486983. S2CID 218587855.
- Wedin, Mats; Grube, Martin (2002). "(1555) Proposal to conserve Physciaceae nom. cons. against an additional name Caliciaceae (Lecanorales, Ascomycota)". Taxon. 51 (4): 802. doi:10.2307/1555043. JSTOR 1555043.
- Gaya, Ester; Högnabba, Filip; Holguin, Ángela; Molnar, Katalin; Fernández-Brime, Samantha; Stenroos, Soili; Arup, Ulf; Søchting, Ulrik; Boom, Pieter Van den; Lücking, Robert; Sipman, Harrie J.M.; Lutzoni, François (2012). "Implementing a cumulative supermatrix approach for a comprehensive phylogenetic study of the Teloschistales (Pezizomycotina, Ascomycota)". Molecular Phylogenetics and Evolution. 63 (2): 374–387. doi:10.1016/j.ympev.2012.01.012. PMID 22306043.
- Wijayawardene, Nalin; Hyde, Kevin; Al-Ani, LKT; Dolatabadi, S; Stadler, Marc; Haelewaters, Danny; et al. (2020). "Outline of Fungi and fungus-like taxa". Mycosphere. 11: 1060–1456. doi:10.5943/mycosphere/11/1/8.
- Prieto, Maria; Wedin, Mats (2016). "Phylogeny, taxonomy and diversification events in the Caliciaceae". Fungal Diversity. 82 (1): 221–238. doi:10.1007/s13225-016-0372-y.
- Cannon, Paul F.; Kirk, Paul M. (2007). Fungal Families of the World. CAB International. pp. 48–49. ISBN 978-0851998275.
- Joneson, Suzanne; Glew, Katherine A. (2003). "Acroscyphus (Caliciaceae) in North America". The Bryologist. 106 (3): 443–446. doi:10.1639/10. S2CID 86756131.
- McCune, Bruce; Rosentreter, Roger (1992). "Texosporium sancti-jacobi, a rare western North American lichen". The Bryologist. 95 (3): 329–333. doi:10.2307/3243492. JSTOR 3243492.
- McCarthy, John W.; Clayden, Stephen R.; Ahti, Teuvo. "Tholurna dissimilis (Caliciaceae) confirmed as occurring in eastern North America". Opuscula Philolichenum. 19: 1–8.
- Species Fungorum. "Caliciaceae". Catalog of Life. Retrieved 15 January 2021.
- Gray, Samuel F. (1821). A Natural Arrangement of British Plants. Vol. 1. London: Baldwin, Cradock, and Joy. p. 482.
- Léveillé, J.H. (1846). "Descriptions des champignons de l'herbier du Muséum de Paris". Annales des Sciences Naturelles Botanique. 3 (in French). 5: 262.
- Scheidegger, Christoph (2007). "A revision of European saxicolous species of the genus Buellia De Not. and formerly included genera". The Lichenologist. 25 (4): 315–364. doi:10.1006/lich.1993.1001. S2CID 85578016.
- Matzer, M.; Mayrhofer, H.; Elix, J.A. (2007). "Australiaena streimannii, a new genus and species in the Physciaceae from tropical Australasia". The Lichenologist. 29 (1): 39–79. doi:10.1017/S0024282997000078.
- Marbach 2000, p. 113.
- De Notaris, G. (1846). "Framennti lichenographici di un lavoro inedito". Parlatore, Giornale Botanico Italiano (in Italian). 2: 174–224.
- Fryday, Alan M.; Medeiros, Ian D.; Siebert, Stefan J.; Pope, Nathaniel; Rajakaruna, Nishanta (2020). "Burrowsia, a new genus of lichenized fungi (Caliciaceae), plus the new species B. cataractae and Scoliciosporum fabisporum, from Mpumalanga, South Africa". South African Journal of Botany. 132: 471–481. doi:10.1016/j.sajb.2020.06.001. S2CID 221143820.
- Vainio, Edvard August (1927). Lichenographia Fennica III. Coniocarpaceae (PDF). Acta Societatis pro Fauna et Flora Fennica. Vol. 57. Helsinki: J. Simelii arvingars boktryckeri a.b. p. 52.
- Marbach 2000, p. 151.
- Marbach 2000, p. 158.
- Kirk, Paul (ed.). "Record Details: Ciposia Marbach". Index Fungorum. Retrieved 15 January 2021.
- Kirk, Paul (ed.). "Record Details: Tylophoropsis Sambo". Index Fungorum. Retrieved 15 January 2021.
- Marbach 2000, p. 160.
- Esslinger, Theodore L. (2000). "Culbersonia americana, a rare new lichen (Ascomycota) from Western America". The Bryologist. 103 (4): 771–773. doi:10.1639/0007-2745(2000)103[0771:CAARNL]2.0.CO;2. S2CID 86045065.
- Nylander, W. (1867). "Circa genus Lichenum Dermatiscum". Botanische Zeitung (in Latin). 25: 133.
- Hafellner, J.; Mayrhofer, H.; Poelt, J. (1979). "Die Gattungen der Flechtenfamilie Physciaceae". Herzogia (in German). 5 (1–2): 39–79. doi:10.1127/herzogia/5/1979/39. S2CID 249722749.
- Norman, J.M. (1852). "Conatus praemissus redactionis novae generum nonnullorum Lichenum in organis fructificationes vel sporis fundatae". Nytt Magazin for Naturvidenskapene (in Latin). 7: 213–252 (see pp. 231, 236).
- Massalongo, Abramo Bartolomeo (1852). Ricerche sull'autonomia dei licheni crostosi e materiali pella loro naturale ordinazione (in Italian). Verona: Dalla tipografia di A. Frizierio. p. 86. OCLC 23511705.
- Flotow, J.V. (1849). "Lichenes florae silesiae". Jahresbericht der Schlesischen Gesellschaft für Vaterländische Kultur (in German). 27: 130.
- Clements, Frederic E. (1909). The Genera of Fungi. Minneapolis: The H. W. Wilson Co. p. 84.
- Marbach 2000, p. 201.
- Marbach 2000, p. 207.
- Fée, Antoine Laurent Apollinaire (1825). Essai sur les cryptogames des écorces exotiques officinales (in French). Paris: Firmin Didot père et fils. p. 100.
- Marbach 2000, p. 291.
- Trevisan, Victore (1856). "Brigantiaea, novum Lichenum genus". Linnaea (in Latin). 28: 283–298.
- Kalb, K.; Giralt, M. (2011). "Orcularia, a segregate from the lichen genera Buellia and Rinodina (Lecanoromycetes, Caliciaceae)". Phytotaxa. 38: 53–60. doi:10.11646/phytotaxa.38.1.8.
- Fries, Elias M. (1825). Systema Orbis Vegetabilis (in Latin). Vol. 1. Lundin: Typographia Academica. p. 267.
- Dodge, C.W. (1973). Lichen flora of the Antarctic continent and adjacent islands. Canaan, New Hampshire: Phoenix Publishing. ISBN 978-0914016014.
- Hale, Mason E.; Vobis, Gernot (1978). "Santessonia, a new lichen genus from Southwest Africa". Botaniska Notiser. 131 (1): 1–5.
- Marbach 2000, p. 296.
- Woronichin, N.N. (1927). "Contribution à la flore mycologique du Caucase". Travaux du Musée Botanique de l'Académie des Sciences de Russie (in French). 21: 87–243 (see p. 103).
- Marbach 2000, p. 304.
- Tibell, Leif; Hofsten, Angelica van (1968). "Spore evolution of the lichen Texosporium sancti-jacobi (=Cyphelium sancti-jacobi)". Mycologia. 60 (3): 553–558. doi:10.1080/00275514.1968.12018604. JSTOR 3757422.
- Massalongo, Abramo Bartolomeo (1860). "Esame comparativo di alcune genere di licheni". Atti dell'Imperial Regio Istituto Veneto di Scienze (in Italian). 5 (3): 247–276.
- Norman, J.M. (1861). "Descriptio speciei novae lichenis, quam detexit et sub nomine Tholurna dissimilis proponit". Flora (Regensburg) (in Latin). 44: 409.
- Cengia Sambo, Maria (1938). "Licheni del Kenia e del Tanganica raccotti dai rev. Padri della Consolata". Nuovo Giornale Botanico Italiano (in Italian). 45 (3): 364–387. doi:10.1080/11263503809438758.
- Moberg, R.; Nordin, A.; Scheidegger, C. (1999). "(1384) Proposal to change the listed type of the name Buellia, nom. cons. (Physciaceae, lichenised Ascomycota)". Taxon. 48 (1): 14. doi:10.2307/1224634. JSTOR 1224634.
- Gams, Walter (2004). "Report of the Committee for Fungi: 11". Taxon. 53 (4): 1067–1069. doi:10.2307/4135578. JSTOR 4135578.
- Tibell, Leif; Knutsson, Tommy (2016). "Calicium episcalaris, (Caliciaceae), a new lichen species from Sweden" (PDF). Symbolae Botanicae Upsaliensis. 38: 49–52.
- Selva, Steven B. (2013). "The calicioid lichens and fungi of the Acadian Forest Ecoregion of northeastern North America, I. New species and range extensions". The Bryologist. 116 (3): 248–256. doi:10.1639/0007-2745-116.3.248. S2CID 86640485.
- Lucban, M.C.; Paguirigan, J.A.G. (2019). "Occurrence of manglicolous lichens in Calabarzon, Philippines". Studies in Fungi. 4 (1): 263–273. doi:10.5943/sif/4/1/28.
- Sethy, P. (2012). "Lichens on mangrove plants in Andaman Islands, India". Mycosphere. 3 (4): 476–484. doi:10.5943/mycosphere/3/4/11.
- Rangsiruji, Achariya; Meesim, Sanya; Buaruang, Kawinnat; Boonpragob, Kansri; Mongkolsuk, Pachara; Binchai, Sutheewan; Pringsulaka, Onanong; Parnmen, Sittiporn (2020). "Molecular systematics and species distribution of foliose lichens in the Gulf of Thailand mangroves with emphasis on Dirinaria picta species complex" (PDF). Songklanakarin Journal of Science and Technology. 42 (3): 504–514. doi:10.14456/sjst-psu.2020.65.
- Coleine, Claudia; Pombubpa, Nuttapon; Zucconi, Laura; Onofri, Silvano; Stajich, Jason E.; Selbmann, Laura (2020). "Endolithic fungal species markers for harshest conditions in the McMurdo Dry Valleys, Antarctica". Life. 10 (2): 13. Bibcode:2020Life...10...13C. doi:10.3390/life10020013. PMC 7175349. PMID 32041249.
- Rambold, Gerhard; Friedl, Thomas; Beck, Andreas (1998). "Photobionts in lichens: possible indicators of phylogenetic relationships?". The Bryologist. 101 (3): 392–397. doi:10.2307/3244177. JSTOR 3244177.
- Spribille, T.; Bilovitz, P.; Printzen, C.; Haugan, R.; Timdal, E. (2015). "Buellia asterella". IUCN Red List of Threatened Species. 2015. Retrieved 15 January 2021.
- Fryday, A. (2020). "Buellia gypsyensis". IUCN Red List of Threatened Species. 2020. Retrieved 15 January 2021.
- Allen, J.; Lendemer, J.; McMullin, T. (2020). "Buellia sharpiana". IUCN Red List of Threatened Species. 2020. Retrieved 15 January 2021.
- Reese Næsborg, R. (5 May 2021). "Calicium sequoiae". IUCN Red List of Threatened Species. 5 May 2021. Retrieved 5 September 2021.
- Stone, D.; Root, H.; Hollinger, J.; Rosentreter, R.; Chandler, A.; Allen, J. (2020). "Texosporium sancti-jacobi". IUCN Red List of Threatened Species. 2020. Retrieved 15 January 2021.
- Lendemer, J. (2020). "Thelomma carolinianum". IUCN Red List of Threatened Species. 2020. Retrieved 15 January 2021.
- Yijian, Yao; Jiangchun, Wei; Wenying, Zhuang; Tiezheng, Wei; Yi, Li; Xinli, Wei; et al. (2020). "Threatened species list of China's macrofungi". Biodiversity Science. 28 (1): 20–25. doi:10.17520/biods.2019174.
- Tibell, Leif (1992). "Crustose lichens as indicators of forest continuity in boreal coniferous forests". Nordic Journal of Botany. 12 (4): 427–450. doi:10.1111/j.1756-1051.1992.tb01325.x.
- Selva, Steven B. (2003). "Using calicioid lichens and fungi to assess ecological continuity in the Acadian Forest Ecoregion of the Canadian Maritimes". The Forestry Chronicle. 79 (3): 550–558. doi:10.5558/tfc79550-3. S2CID 84488632.
- Bajpai, Rajesh; Upreti, Dalip K.; Nayaka, S.; Kumari, B. (2010). "Biodiversity, bioaccumulation and physiological changes in lichens growing in the vicinity of coal-based thermal power plant of Raebareli district, north India". Journal of Hazardous Materials. 174 (1–3): 429–436. doi:10.1016/j.jhazmat.2009.09.071. PMID 19818555.
- Singh, Pramod Kumar; Bujarbarua, P.; Singh, K.P.; Tandon, P.K. (2019). "Report on the bioaccumulation of heavy metals by foliose lichen (Pyxine cocoes) from air polluted area near Nagaon Paper Mill in Marigaon, Assam, North-East India". Journal on New Biological Reports. 8 (1): 15–21. ISSN 2319-1104.
- Shukla, Vertika; Upreti, Dalip K. (2007). "Effect of metallic pollutants on the physiology of lichen, Pyxine subcinerea Stirton in Garhwal Himalayas". Environmental Monitoring and Assessment. 141 (1–3): 237–243. doi:10.1007/s10661-007-9891-z. PMID 17879139. S2CID 1325600.
- Shukla, Vertika; Upreti, Dalip K. (2010). "Changing lichen diversity in and around urban settlements of Garhwal Himalayas due to increasing anthropogenic activities". Environmental Monitoring and Assessment. 174 (1–4): 439–444. doi:10.1007/s10661-010-1468-6. PMID 20440642. S2CID 33101795.
- Meeßen, J.; Sánchez, F. J.; Brandt, A.; Balzer, E.-M.; de la Torre, R.; Sancho, L. G.; de Vera, J.-P.; Ott, S. (2013). "Extremotolerance and resistance of lichens: comparative studies on five species used in astrobiological research I. Morphological and anatomical characteristics". Origins of Life and Evolution of Biospheres. 43 (3): 283–303. Bibcode:2013OLEB...43..283M. doi:10.1007/s11084-013-9337-2. PMID 23868319. S2CID 254893830.
- Rikkinen, Jouko (2003). "Calicioid lichens from European Tertiary amber". Mycologia. 95 (6): 1032–1036. doi:10.1080/15572536.2004.11833019. PMID 21149012. S2CID 36366954.
- Rikkinen, Jouko; Meinke, S. Kristin L.; Grabenhorst, Heinrich; Gröhn, Carsten; Kobbert, Max; Wunderlich, Jörg; Schmidt, Alexander R. (2018). "Calicioid lichens and fungi in amber – Tracing extant lineages back to the Paleogene". Geobios. 51 (5): 469–479. Bibcode:2018Geobi..51..469R. doi:10.1016/j.geobios.2018.08.009. hdl:10138/308761. S2CID 135125977.
- Kettunen, Elina; Sadowski, Eva-Maria; Seyfullah, Leyla J.; Dörfelt, Heinrich; Rikkinen, Jouko; Schmidt, Alexander R.; Lomax, Barry (2019). "Caspary's fungi from Baltic amber: historic specimens and new evidence". Papers in Palaeontology. 5 (3): 365–389. doi:10.1002/spp2.1238. hdl:10138/309518. S2CID 92394285.
Cited literature
- Marbach, Bernhard (2000). Corticole und lignicole Arten der Flechtengattung Buellia sensu lato in den Subtropen und Tropen [A study of corticolous and lignicolous species of the lichen genus Buellia s.l. in subtropical and tropical regions]. Bibliotheca Lichenologica (in German). Vol. 74. J. Cramer. pp. 1–384. ISBN 978-3-443-58053-7.