Cladoniaceae
The Cladoniaceae are a family of lichenized fungi in the order Lecanorales.[2] It is one of the largest families of lichen-forming fungi, with about 560 species distributed amongst 17 genera. The reindeer moss and cup lichens (Cladonia) belong to this family. The latter genus, which comprises about 500 species, forms a major part of the diet of large mammals in taiga and tundra ecosystems.[5] Many Cladoniaceae lichens grow on soil, but other can use decaying wood, tree trunks, and, in a few instances, rocks as their substrate. They grow in places with high humidity, and cannot tolerate aridity.
| Cladoniaceae | |
|---|---|
 | |
| Cladonia subuluata | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Fungi |
| Division: | Ascomycota |
| Class: | Lecanoromycetes |
| Order: | Lecanorales |
| Family: | Cladoniaceae Zenker (1827)[1] |
| Type genus | |
| Cladonia P.Browne (1756) | |
| Synonyms[2] | |
Many Cladoniaceae species are characterized by a thallus that has two distinct forms: a scaly or crust-like primary thallus that, depending on the species, can be permanent or temporary, and a secondary fruticose thallus called a podetium or pseudopodetium. Cladoniaceae members form symbiotic associations with green algae from the class Trebouxiophyceae, usually the genus Asterochloris. Five Cladoniaceae species are included in the International Union for Conservation of Nature's Red List of Threatened Species.
Taxonomy
Cladoniaceae was formally introduced to science in 1827 by German naturalist Jonathan Carl Zenker in a publication of Karl Goebel.[1] Zenker's initial concept of the family included genera that are now recognized as separate families, including Baeomycetaceae, Icmadophilaceae, and Stereocaulaceae.[6] William Nylander included 53 Cladonia species worldwide in his 1860 work Synopsis lichenum.[7] When Edvard August Vainio published his three-volume monograph on the Cladoniaceae (Monographia Cladoniarum universalis, 1887,[8] 1894,[9] and 1897[10]), he included 134 species and subspecies. In his circumscription of the family, the genera Pycnothelia, Cladia, and Cladina were included in the genus Cladonia.[11]
Cladoniaceae is now one of the largest families of lichen-forming fungi,[12] with about 560 species distributed amongst 18 genera. The type genus, after which the family was named, is Cladonia, circumscribed by Irish physician and botanist Patrick Browne in 1756. He included 8 species in his new genus. Of their occurrence, he wrote: "All these species are found in great abundance in the mountains of Liguanea: they grow mostly on the ground, among other sorts of moss, but a few ... species chiefly are found upon the decaying trunks of trees."[13]
Synonymy
Several phylogenetic studies have shown that Cladoniaceae is a member of the order Lecanorales, and is closely related to the family Stereocaulaceae.[14][15][16][17][18] The family Cetradoniaceae, which was created in 2002 to contain the endangered species Cetradonia linearis, was folded into the Cladoniaceae in 2006.[19]
In 2018, Kraichak and colleagues used a technique called temporal banding to reorganize the Lecanoromycetes, proposing a revised system of classification based on correlating taxonomic rank with geological (evolutionary) age. They synonymized the families Squamarinaceae and Stereocaulaceae with the Cladoniaceae, resulting in a large increase in the number of genera and species.[20] The Squamarinaceae had already been included in the Cladoniaceae by previous authors.[12] Although this reorganization has been used in some later publications,[2] the folding of the Stereocaulaceae into the Cladoniaceae was not accepted in a recent analysis. As Robert Lücking explained, "merging of the two families under the name Cladoniaceae is not possible without a conservation proposal because Cladoniaceae (Zenker, 1827) is antedated by Stereocaulaceae (Chevallier, 1826) by one year."[21]
Description
.jpg.webp)
The thallus of Cladoniaceae lichens are fruticose or foliose, and are often dimorphic–consisting of two distinct forms; this particular type of growth is also known as cladoniiform.[22] The primary thallus is ephemeral to persistent, crustose, foliose or squamulose, while the secondary thallus is typically vertical and holds the ascomata.[23] The secondary thallus ranges in height from a few millimetres to more than 25 cm (10 in). Some species, however, form neither a primary thallus nor any fruticose structures.[24]
The ascomata are in the form of an apothecium, and are biatorine, meaning they are of the lecideine type – light in colour and soft in consistency. They often have a reduced margin.[23] Their colour is typically dark brown (sometimes pale brown), red, ochraceous, or black.[24] The hamathecium (referring to all hyphae between the asci in the hymenium) consists of branched paraphyses, and is amyloid. The asci are somewhat fissitunicate, meaning they have two layers that separate during ascus dehiscence. The ascus structure consists of an apical dome and a tube (both of which are amyloid), which is cylindrical to clavate. Ascospores number eight per ascus, and they are usually non-septate, ellipsoid to more or less spherical in shape, hyaline, and non-amyloid.[23] Except for a few genera that produce septate ascospores (Calathaspis, Pycnothelia and Pilophorus), the hymenium does not generally have characters that are useful in taxonomy.[24] The conidiomata are pycnidia; the conidia are non-septate, usually filiform (thread-like), and hyaline.[23]
Photobionts
The symbiotic algal partner (photobiont) of most Cladoniaceae taxa are unicellular green algae, usually in the genus Asterochloris, but occasionally in the genus Chlorella;[25] both of these genera are in the class Trebouxiophyceae. Eleven species of Asterochloris have found to be associated with genus Cladonia; the algal genus – one of the most common lichen symbionts – occurs in the thalli of more than 20 lichen genera.[26] The most common photobionts in this genus that associate with Cladonia are A. glomerata, A. italiana, and A. mediterranea, with some lineages showing dominance in one or several climatic regions.[27] The algal genus Trebouxia, a very common lichen photobiont, has not been recorded associating with the Cladoniacae.[24] Some Pilophorus species associate with cyanobacterial symbionts (in addition to their association with green algae) in structures called cephalodia.[25] The cyanobacterial genera Nostoc and Stigonema are involved in these tripartite associations.[24]
Development



The development of several Cladoniaceae genera have been studied in detail,[28][29][30][31][32][33] although the interpretation of results has sometimes been controversial.[24] Cladoniaceae species begin development with the formation of a prothallus – a fungal layer upon which an algae-containing thallus will develop. It comprises the hyphae from the germination of an ascospore. After the protothallus contacts the alga, lichenization begins with the development of small squamules (scale-like thallus segments) that make up the primary thallus. Most Cladoniaceae have a mixed thallus, consisting of two parts: a base, parallel to the substrate, called the primary thallus and the other erect, the secondary thallus. The primary thallus is squamulose (scaly) or crustose (crustose-like). The secondary thallus consists of vertical structures that are shrubby and hollow, although they can be solid in rare cases. If these structures are made of generative tissue, they are called podetia; when they are made of vegetative tissue, they are called pseudopodetia. The morphology of these structures determines to a large part the taxonomy of the Cladoniaceae, which can range from simple to very complex branching patterns.[25] Cladonia minisaxicola, found in the mountains of Bahia (Brazil) is the only species in that large genus that is completely crustose and does not develop podetia.[34]
The tips of the podetia have a wide range of morphology in the Cladoniaceae. They can be straight, tapering from a wide base to a point (called subulate), or flaring on cup-shaped scyphi. The scyphi are sometimes closed, or have a central perforation, forming structures called funnels.[25] The podetia are slow-growing, with an annual growth rate generally ranging from 1 to 15 mm.[35]
Branching in the Cladoniaceae occurs on the podetium due to the growth dynamics of fungal meristem tissue at its apex. Two primary branching patterns exist: one where branches emerge from late divisions of a large meristem that alters its shape, and another where branches come from small meristems that split early but maintain their shape. These meristem growth dynamics are crucial for interpreting phylogeny in Cladoniaceae mycobionts, with the trend towards smaller, early-splitting meristems seen as an evolutionary advancement.[36] In cladoniiform lichens, especially within the Cladoniaceae, a shift in meristem growth from isotropous (having uniform properties in all directions) to anisotropous (having properties that differ depending on the direction in which they are measured) leads to pronounced lateral elongation of the apical meristem. This change offers developmental flexibility, transitioning from a symmetrical growth to a more varied, asymmetrical growth, hinting at evolutionary processes within the Cladoniaceae. Despite this variability, such morphogenetic activities appear to be highly conserved even among species that are presumably distantly related.[37]
Habitat and distribution
Cladoniaceae species have been recorded growing in many habitats and on a diversity of substrates, including soil, tree trunks, and rotten wood.[24] In a few cases, Cladoniaceae can grow on rocks, such as Cladonia salmonea which grows on the rock faces of vertical cliffs, or Cladonia pyxidata, which can grow on thin soil on rocks.[38] They are absent from very dry regions. The range of their habitats includes boreal forests, bogs, temperate forests, the tundra of the Arctic and Antarctic, man-made habitats (e.g. roadsides), tropical highlands, and the sandy tropical lowlands of the Amazon rainforest.[24] Twenty-six Cladoniaceae species (25 Cladonia and 1 Cladia) are known to occur in the Galapagos Islands. There, some species form mats on lava flows that have developed little soil.[39] A 2013 monograph of Northern European Cladoniaceae treated 100 species (95 Cladonia, 4 Pilophorus, and the monotypic genus Pycnothelia).[40] In the 2021 key to lichen species in Italy, 86 Cladoniaceae are included.[41]
A study on the distributional ecology of Cladina and Cladonia in western North America found that the Coast Mountains of British Columbia act as a key phytogeographic barrier. This results in distinct oceanic and continental taxa groupings on either side. The research also suggests that the southern boundaries of certain species may be determined more by historical rather than purely ecological factors, indicating possible range expansions.[42]
Conservation
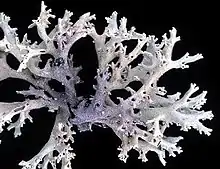
Five Cladoniaceae species have been assessed for the global IUCN Red List: Cetradonia linearis (vulnerable, 2015),[43] Cladonia appalachiensis (endangered, 2020),[44] Cladonia perforata (endangered, 2003),[45] Pilophorus fibula (endangered, 2020),[46] and Gymnoderme insulare (endangered, 2014).[47] On the red list of China's macrofungi, Cladonia delavayi (vulnerable), Cladonia pseudoevansii (critically endangered), Gymnoderma coccocarpum (endangered), and Gymnoderma insulare (endangered) are the representatives of the Cladoniaceae.[48]
Genera
After more than a century of discovery and research, including recent advances in understanding revealed by molecular phylogenetic studies, the Cladoniaceae encompass 17 genera and more than 550 species. This is a list of the genera contained within the Cladoniaceae, based on the Catalogue of Life; this includes taxa formerly classified in the Squamarinaceae, but does not include the Stereocaulaceae. Following the genus name is the taxonomic authority, year of publication, and the number of species:


- Calathaspis I.M.Lamb & W.A.Weber (1972)[49] – 1 sp.
- Carassea S.Stenroos (2002)[15] – 1 sp.
- Cetradonia J.C.Wei & Ahti (2002)[3] – 1 sp.
- Cladia Nyl. (1870)[50] – ca. 27 spp.
- Cladonia Hill ex P.Browne (1756)[13] – ca. 500 spp.
- Heteromyces Müll.Arg. (1889)[51] – 1 sp.
- Metus D.J.Galloway & P.James (1987)[52] – 3 spp.
- Muhria P.M.Jørg. (1987)[53] – 1 sp.
- Notocladonia S.Hammer (2003)[54] – 2 spp.
- Paralecia Brackel, Greiner, Peršoh & Rambold (2015)[55] – 1 sp.
- Pilophorus Th.Fr. (1857) – 17 spp.
- Pulchrocladia S.Stenroos, Pino-Bodas, Lumbsch & Ahti (2018)[24] – 3 spp.
- Pycnothelia Dufour (1821)[56] – 2 spp.
- Rexiella S.Stenroos, Pino-Bodas & Ahti (2019)[57] – 1 sp.
- Sphaerophoropsis Vain. (1890)[58] – 2 spp.
- Squamella S.Hammer (2001)[59] – 1 sp.
- Thysanothecium Mont. & Berk. (1846)[60] – 3 spp.
Myelorrhiza was transferred from the Cladoniaceae to the Ramalinaceae by Kistenich and colleagues in 2018.[61] Neophyllis, originally classified in the Cladoniaceae, was transferred to Sphaerophoraceae in 1999.[62]
References
- Goebel, Karl Christian Traugott Friedemann; Kunze, G. (1827). Pharmaceutische Waarenkunde (in German). Eisenach: J.F. Bärecke. p. 124.
- Wijayawardene, Nalin; Hyde, Kevin; Al-Ani, LKT; Dolatabadi, S; Stadler, Marc; Haelewaters, Danny; et al. (2020). "Outline of Fungi and fungus-like taxa". Mycosphere. 11: 1060–1456. doi:10.5943/mycosphere/11/1/8.
- Wei, Jiang-Chun; Ahti Teuvo (2002). "Cetradonia, a new genus in the new family Cetradoniaceae (Lecanorales, Ascomycota)". The Lichenologist. 34 (1): 19–31. doi:10.1006/lich.2001.0354. S2CID 86545255.
- Hafellner, J. (1984). "Studien in Richtung einer natürlichen Gliederung der Sammelfamilien Lecanoracae und Lecideaceae". Beihefte zur Nova Hedwigia. 79: 241–371.
- Cannon, Paul F.; Kirk, Paul M. (2007). Fungal Families of the World. CAB International. p. 73. ISBN 978-0-85199-827-5.
- Ahti 2000, p. 3.
- Nylander, William (1860). Synopsis methodica lichenum (in Latin). Paris: Imprimerie de L. Martinet.
- Wainio, Edvard August (1887). Monographia Cladoniarum universalis: I. Acta Societatis pro Fauna et Flora Fennica. Vol. 4. pp. 1–509.
- Wainio, Edvard August (1894). Monographia Cladoniarum universalis: II. Acta Societatis pro Fauna et Flora Fennica. Vol. 10. pp. 1–499.
- Wainio, Edvard August (1897). Monographia Cladoniarum universalis: III. Acta Societatis pro Fauna et Flora Fennica. Vol. 14. pp. 1–268.
- Ahti 2000, p. 4.
- Lücking, Robert; Hodkinson, Brendan P.; Leavitt, Steven D. (2017). "The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota–Approaching one thousand genera". The Bryologist. 119 (4): 361–416. doi:10.1639/0007-2745-119.4.361. S2CID 90258634.
- Browne, Patrick (1756). Civil and natural history of Jamaica. London. p. 81.
- Wedin, Mats; Döring, Heidi; Ekman, Stefan (2000). "Molecular phylogeny of the lichen families Cladoniaceae, Sphaerophoraceae, and Stereocaulaceae (Lecanorales, Ascomycotina)". The Lichenologist. 32 (2): 171–187. doi:10.1006/lich.1999.0236. S2CID 86041209.
- Stenroos, Soili; Myllys, Leena; Thell, Arne; Hyvönen, Jaakko (2002). "Phylogenetic hypotheses: Cladoniaceae, Stereocaulaceae, Baeomycetaceae, and Icmadophilaceae revisited". Mycological Progress. 1 (3): 267–282. doi:10.1007/s11557-006-0024-9. S2CID 22243440.
- Arup, U.; Ekman, S.; Grube, M.; Mattsson, J.-E.; Wedin, M. (2007). "The sister group relation of Parmeliaceae (Lecanorales, Ascomycota)". Mycologia. 99 (1): 42–49. doi:10.1080/15572536.2007.11832599. PMID 17663122. S2CID 218587865.
- Ekman, Stefan; Andersen, Heidi L.; Wedin, Mats; Buckley, Thomas (2008). "The limitations of ancestral state reconstruction and the evolution of the ascus in the Lecanorales (lichenized Ascomycota)". Systematic Biology. 57 (1): 141–156. doi:10.1080/10635150801910451. PMID 18300027.
- Miadlikowska, Jolanta; Kauff, Frank; Högnabba, Filip; Oliver, Jeffrey C.; Molnár, Katalin; Fraker, Emily; et al. (2014). "A multigene phylogenetic synthesis for the class Lecanoromycetes (Ascomycota): 1307 fungi representing 1139 infrageneric taxa, 317 genera and 66 families". Molecular Phylogenetics and Evolution. 79: 132–168. doi:10.1016/j.ympev.2014.04.003. PMC 4185256. PMID 24747130.
- Zhou, Qi-Ming; Wei, Jiang-Chun; Ahti, Teuvo; Stenroos, Soili; Högnabba, Filip (2006). "The systematic position of Gymnoderma and Cetradonia based on SSU rDNA sequences". Journal of the Hattori Botanical Laboratory. 100: 871–880.
- Kraichak, Ekaphan; Huang, Jen-Pan; Nelsen, Matthew; Leavitt, Steven D.; Lumbsch, H. Thorsten (2018). "A revised classification of orders and families in the two major subclasses of Lecanoromycetes (Ascomycota) based on a temporal approach". Botanical Journal of the Linnean Society. 188 (3): 233–249. doi:10.1093/botlinnean/boy060.
- Lücking, Robert (2019). "Stop the abuse of time! Strict temporal banding is not the future of rank-based classifications in fungi (including lichens) and other organisms". Critical Reviews in Plant Sciences. 38 (3): 199–253. doi:10.1080/07352689.2019.1650517. S2CID 202859785.
- Ahti, T. (1982). "The morphological interpretation of cladoniiform thalli in lichens". The Lichenologist. 14 (2): 105–113. doi:10.1017/s0024282982000255. S2CID 86304863.
- Jaklitsch, Walter; Baral, Hans-Otto; Lücking, Robert; Lumbsch, H. Thorsten (2016). Frey, Wolfgang (ed.). Syllabus of Plant Families: Adolf Engler's Syllabus der Pflanzenfamilien. Vol. 1/2 (13 ed.). Berlin Stuttgart: Gebr. Borntraeger Verlagsbuchhandlung, Borntraeger Science Publishers. p. 121. ISBN 978-3-443-01089-8. OCLC 429208213.
- Stenroos, Soili; Pino‐Bodas, Raquel; Hyvönen, Jaakko; Lumbsch, Helge Thorsten; Ahti, Teuvo (2018). "Phylogeny of the family Cladoniaceae (Lecanoromycetes, Ascomycota) based on sequences of multiple loci". Cladistics. 35 (4): 351–384. doi:10.1111/cla.12363. hdl:10261/247495. PMID 34633698. S2CID 92664622.
- Burgaz, Ana Rosa; Ahti, Teuvo; Pino-Bodas, Raquel (2020). Mediterranean Cladoniaceae. Madrid: Spanish Lichen Society (SEL). ISBN 978-84-09-21610-9.
- Škaloud, Pavel; Steinová, Jana; Řídká, Tereza; Vančurová, Lucie; Peksa, Ondřej; De Clerck, O. (2015). "Assembling the challenging puzzle of algal biodiversity: species delimitation within the genus Asterochloris (Trebouxiophyceae, Chlorophyta)". Journal of Phycology. 51 (3): 507–527. doi:10.1111/jpy.12295. PMID 26986666. S2CID 25190572.
- Pino-Bodas, Raquel; Stenroos, Soili (2020). "Global biodiversitypatterns of the photobionts associated with the genus Cladonia (Lecanorales, Ascomycota)". Microbial Ecology. 82 (1): 173–187. doi:10.1007/s00248-020-01633-3. PMC 8282589. PMID 33150498.
- Jahns, H.M. (1970). Untersuchungen zur Entwicklungsgeschichte der Cladoniaceen unter besonderer Berücksichtigung des Podetien- Problems. Nova Hedwigia (in German). Vol. 20. pp. 1–177.
- Jahns, H.M. (1970). "Induktion der Apothecienbildung bei Cladia aggregata (Sw.) Nyl". Berichte der Deutschen Botanischen Gesellschaft (in German). 83: 33–40. doi:10.1111/j.1438-8677.1970.tb02301.x. S2CID 82100599.
- Jahns, H.M.; Beltman, H.A. (2007). "Variations in the ontogeny of fruiting bodies in the genus Cladonia and their taxonomic and phylogenetic significance". The Lichenologist. 5 (5–6): 349–367. doi:10.1017/S0024282973000447. S2CID 86386885.
- Hammer, Samuel (1992). "Development in Cladonia ochrochlora". Mycologia. 85 (1): 84–92. doi:10.1080/00275514.1993.12026250.
- Hammer, Samuel (2018). "Primary tissue and the structure of the podetium in Cladonia". Mycologia. 87 (1): 46–53. doi:10.1080/00275514.1995.12026501.
- Grube, Martin; Hawksworth, David L. (2007). "Trouble with lichen: the re-evaluation and re-interpretation of thallus form and fruit body types in the molecular era". Mycological Research. 111 (9): 1116–1132. doi:10.1016/j.mycres.2007.04.008. PMID 17698333.
- Aptroot, André; da Silva Cáceres, Marcela Eugenia (2018). "New lichen species from Chapada Diamantina, Bahia, Brazil". The Bryologist. 121 (1): 67–79. doi:10.1639/0007-2745-121.1.067. S2CID 90659999.
- Ahti 2000, p. 12.
- Hammer, Samuel (2000). "Meristem growth dynamics and branching patterns in the Cladoniaceae". American Journal of Botany. 87 (1): 33–47. doi:10.2307/2656683.
- Hammer, Samuel (2001). "Lateral growth patterns in the Cladoniaceae". American Journal of Botany. 88 (5): 788–796. doi:10.2307/2657031.
- Ahti 2000, p. 23.
- Yánez, Alba; Ahti, Teuvo; Bungartz, Frank (2013). "The Family Cladoniaceae (Lecanorales) in the Galapagos Islands". Phytotaxa. 129 (1): 1–33. doi:10.11646/phytotaxa.129.1.1.
- Ahti, Teuvo; Stenroos, Soili; Moberg, Roland (2013). Nordic Lichen Flora. Cladoniaceae. Vol. 5. Uppsala: Museum of Evolution, Uppsala University. ISBN 978-91-85221-29-5.
- Gheza, Gabriele; Nimis, Per Luigi (22 July 2021). "Keys to the Lichens of Italy. 61) Cladoniaceae (Cladonia, Pilophorus, and Pycnothelia)". Italic 6.0, The Information System on Italian Lichens. Retrieved 22 March 2022.
- Goward, Trevor; Ahti, Teuvo (1997). "Notes on the distributional ecology of the Cladoniaceae (lichenized ascomycetes) in temperate and boreal western North America". Journal of the Hattori Botanical Laboratory. 82: 143–155.
- Allen, J.; Lendemer, J.; McMullin, T. (11 August 2015). "Rock Gnome Lichen. Cetradonia linearis". The IUCN Red List of Threatened Species 2020. Retrieved 19 February 2021.
- Lendemer, J.; Allen, J.; McMullin, T. (6 October 2019). "Cladonia appalachiensis". The IUCN Red List of Threatened Species 2020. Retrieved 19 February 2021.
- Yahr, R. (30 April 2003). "Cladonia perforata". The IUCN Red List of Threatened Species 2003. Retrieved 19 February 2021.
- Lendemer, J.; Allen, J.; McMullin, T. (8 October 2019). "Appalachian Matchsticks. Pilophorus fibula". The IUCN Red List of Threatened Species 2020. Retrieved 19 February 2021.
- Ohmura, Y.; Nadyeina, O.; Scheidegger, C. (14 August 2014). "Gymnoderme insulare". The IUCN Red List of Threatened Species 2014. Retrieved 19 February 2021.
- Yijian, Yao; Jiangchun, Wei; Wenying, Zhuang; Tiezheng, Wei; Yi, Li; Xinli, Wei; et al. (2020). "Threatened species list of China's macrofungi". Biodiversity Science. 28 (1): 20–25. doi:10.17520/biods.2019174.
- Lamb, I.M.; Weber, W.A. (1974). "Calathaspis, a new genus of lichens (Cladoniaceae) from New Guinea". Occasional Papers of the Farlow Herbarium of Cryptogamic Botany. 4: 1–12.
- Nylander, William (1870). Recognitio Monographica Ramalinarum (in French). Caen. p. 69.
{{cite book}}: CS1 maint: location missing publisher (link) - Müller, J. (1889). "Lichenologische Beiträge XXXII". Flora (Regensburg) (in German). 72 (5): 505–508.
- Galloway, D.J.; James, P.W. (1987). "Metus, a new austral lichen genus and notes on an Australian species of Pycnothelia". Notes from the Royal Botanical Garden Edinburgh. 44 (3): 561–579.
- Jørgensen, P.M.; Jahns, H.M. (1987). "Muhria, a remarkable new lichen genus from Scandinavia". Notes from the Royal Botanical Garden Edinburgh. 44: 581–599.
- Hammer, Samuel (2003). "Notocladonia, a new genus in the Cladoniaceae". The Bryologist. 106 (1): 162–167. doi:10.1639/0007-2745(2003)106[0162:NANGIT]2.0.CO;2. JSTOR 3244806. S2CID 86169366.
- Liu, Jian Kui; Hyde, Kevin D.; Jones, E. B. Gareth; Ariyawansa, Hiran A.; Bhat, Darbhe J.; Boonmee, Saranyaphat; et al. (2015). "Fungal diversity notes 1–110: taxonomic and phylogenetic contributions to fungal species". Fungal Diversity. 72 (1): 1–197. doi:10.1007/s13225-015-0324-y. S2CID 256069348.
- Dufour, J.L.M. (1821). "Révision des genres Cladonia, Scyphophorus, Helopodium et Baeomyces de la flore française". Annales Générales des Sciences Physiques (in French). 8: 41–73.
- Stenroos, Soili; Pino‐Bodas, Raquel; Ahti, Teuvo (2019). "Rexiella, a new name for Rexia S. Stenroos, Pino‐Bodas & Ahti (2018), non Rexia D. A. Casamatta, S. R. Gomez & J. R. Johansen (2006)". Cladistics. 35 (5): 603. doi:10.1111/cla.12401. S2CID 201197971.
- Vainio, E.A. (1890). "Étude sur la classification naturelle et la morphologie des Lichens du Brésil. Pars prima". Acta Societatis Pro Fauna et Flora Fennica (in Latin). 7 (1): 7.
- Hammer, Samuel (2001). "Additions to the lichen family Cladoniaceae in Australia". The Bryologist. 104 (4): 560–575. doi:10.1639/0007-2745(2001)104[0560:ATTLFC]2.0.CO;2. S2CID 86076701.
- Montagne, C.; Berkeley, M.J. (1846). "On Thysanothecium, a new genus of lichens". Hooker's Journal of Botany. 5: 257–258.
- Kistenich, Sonja; Timdal, Einar; Bendiksby, Mika; Ekman, Stefan (2018). "Molecular systematics and character evolution in the lichen family Ramalinaceae (Ascomycota: Lecanorales)". Taxon. 67 (5): 871–904. doi:10.12705/675.1. hdl:10852/67955.
- Wedin, Mats; Döring, Heidi (1999). "The phylogenetic relationship of the Sphaerophoraceae, Austropeltum and Neophyllis (lichenized Ascomycota) inferred by SSU rDNA sequences". Mycological Research. 103 (9): 1131–1137. doi:10.1017/S0953756298008223.
Cited literature
- Ahti, Teuvo (2000). Cladoniaceae. Flora Neotropic Monograph. Vol. 78. Bronx, New York: New York Botanical Garden Press. ISBN 978-0-89327-431-3.