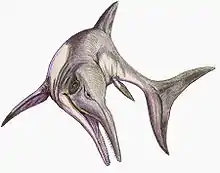
Cymbospondylus
Cymbospondylus (meaning "cupped vertebrae") is an extinct genus of large ichthyosaurs that lived in the Middle Triassic (Anisian), in what are now North America and Europe. The taxon was erected by Joseph Leidy in 1868 on the basis of vertebrae discovered in Nevada, before subsequent research found more complete fossil remains which were described by John Campbell Merriam in 1908. While many species have been assigned to the genus, only five are recognized as valid, the others being considered synonymous, doubtful or belonging to other genera.
| Cymbospondylus Temporal range: Middle Triassic, Middle to Late | |
|---|---|
 | |
| Partial holotype skeleton of C. buchseri (PIMUZ T 4351), on display at the Paleontological Museum of the University of Zurich, Switzerland. | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Order: | †Ichthyosauria |
| Family: | †Cymbospondylidae |
| Genus: | †Cymbospondylus Leidy, 1868 |
| Species | |
| Synonyms | |
As an ichthyosaur, Cymbospondylus had flippers for limbs and a fin on the tail. The different species of Cymbospondylus vary greatly in size, with the smallest reaching around 4 metres (13 ft) to 5 metres (16 ft) in length. The largest known species, C. youngorum, is estimated to be around 17 metres (56 ft) long, making Cymbospondylus one of the largest ichthyosaurs identified to date, but also one of the largest animals known of its time. The animal has a skull with a long, thin snout, proportionally small eye sockets, an elongated trunk, and a less pronounced tail than in later ichthyosaurs.
The genus was formerly classified as a representative of the Shastasauridae, but more recent studies consider it to be more basal, considered to be the only recognized member of the Cymbospondylidae. The teeth are conical and pointed, having longitudinal ridges, indicating a diet of fishes and cephalopods, and possibly other marine reptiles for larger species.
Research history
Discovery and identification
In 1868, American paleontologist Joseph Leidy described two new genera of ichthyosaurs dating from the Middle Triassic on the basis of fossil vertebrae discovered in several localities in Nevada, United States, all of which were transmitted through the geologist Josiah Dwight Whitney. One of the two genera he names is Cymbospondylus, to which he assigns two species. The first one is C. piscosus, which Leidy named on the basis of several more or less complete vertebrae having been discovered in different mountain ranges of the state.[1] The holotype of C. piscosus is a block containing five incomplete dorsal vertebrae that was discovered in the New Pass Range, northwest of the city of Austin.[2][3][4][lower-alpha 1] Leidy attributes two other specimens to the species, one being a series of eight caudal vertebrae discovered at Star Canyon in the Humboldt Range, and the other being a single vertebrae, probably also caudal, discovered in the Toiyabe Range, in the Reese River, northeast of Austin. The second species is C. petrinus, named from five large dorsal vertebrae discovered in the Humboldt Range.[1] The genus name Cymbospondylus derives from the Ancient Greek words κύμβη (kymbē, "cup") and σπόνδυλος (spondylos, "vertebra"), all taken together literally meaning cupped vertebrae, in reference to the rather obvious shape of this part of the skeleton.[5] As no type species was designated in the 1868 article, it was not until 1902 that John Campbell Merriam assigned C. piscosus to this title,[6] the latter being the first named in Leidy's official description of the genus.[1]
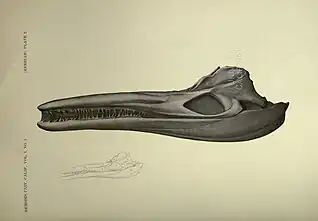
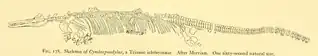
Between 1901 and 1907, the University of California sent ten expeditions across different corners of the United States to recover as many ichthyosaur fossils as possible dating from the Triassic, in order to be analyzed in more detail. These same expeditions were led by Merriam and were almost all financed by Annie Montague Alexander. The research finally collected more than fifty specimens, each preserving a significant portion of their skeletons.[7] Among these fossils are several specimens of excellent quality from C. petrinus, including an almost complete skeleton, cataloged as UCMP 9950, all discovered in the origin locality mentioned by Leidy. Like the other ichthyosaurs mentioned in the work, Merriam describes the taxon in more depth based on new known fossil material.[8][9] Merriam's anatomical descriptions of C. petrinus are still recognized as viable, and are even used in comparative anatomy in later studies of the genus Cymbospondylus, thus lending validity to the species.[3][10][4][11][12]
Unlike C. petrinus, no additional fossils of the type species C. piscosus have been discovered. In Merriam's works, the latter recognized C. piscosus as distinct on the basis of its smaller size and by the regularly concave faces of the vertebrae of the holotype specimen.[6][13] Only later did the validity of C. piscosus begin to be questioned, with authors mentioning the questionable nature of the fossils.[3] In their work published in 2003, Christopher McGowan and Ryosuke Motani assert that the fossil vertebrae attributed to C. piscosus do not present distinctive characteristics to prove its validity. This would therefore pose a taxonomic problem, because if the type species turns out to be a nomen dubium, the genus to which it is classified will also be. To try to resolve this problem, the two authors suggest that the most complete known skeleton of C. petrinus (UCMP 9950) could be designated as a neotype of C. piscosus.[14] However, as no formal ICZN appeal has been established to date, the name C. petrinus for the proposed neotype should be retained until further notice.[4] In 2017, Andrzej Stefan Wolniewicz referred all additional fossil material described by Merriam to C. piscosus and synonymized C. petrinus to the latter.[15] However, As the proposal remains restricted to a PhD thesis, it is defined as an unpublished work per Article 8 of the ICZN and therefore is not formally valid.[16][17] Therefore, C. piscosus is not included in most descriptions of the genus, although it is still recognized as the type species.[11][12]
Recognized species
In 1927, a partial skeleton of a large ichthyosaur was discovered in the Grenzbitumenzone Member at the Monte San Giorgio fossil site in Switzerland and was mentioned by Bernhard Peyer in 1944.[18] In 1964, Emil Kuhn-Schnyder published a photo of this same specimen and suggested that it shared affinities with Cymbospondylus, then only known from North America at that time.[19] The specimen in question, cataloged as PIMUZ T 4351, was formally described for the first time in 1989 under the name C. buchseri by Paul Martin Sander, thus confirming the presence of the genus in Europe. The specific epithet buchseri is named in honor of Fritz Buchser, a member of the Museum of Paleontology at the University of Zurich, the latter having prepared the holotype skeleton in 1931 as his first major professional achievement.[3]

While C. petrinus was for a time seen as the only valid species of the genus known from Nevada, it was not until the early 21th century that later discoveries contradicted this assertion. In 2006, Nadia Fröbisch and colleagues described the species C. nichollsi based on an incomplete skeleton, cataloged as FMNH PR 2251, which was discovered in the Augusta Mountains. The fossil was originally exhumed in the hope of finding a new specimen of C. petrinus, but the number of significant anatomical differences led researchers to establish a separate species. The species in question is named in honor to Elizabeth Nicholls, an American-Canadian paleontologist specializing in Triassic marine reptiles, who made a major contribution to the ichthyosaurs that lived during this period.[4][5] In their description, Fröbisch and his colleagues consider that an almost complete skull attributed to C. petrinus, cataloged as UCMP 9913,[lower-alpha 2] could in fact belong to C. nichollsi, because it presents similar characteristics. However, as the specimen has been relatively little described in the scientific literature, the authors do not know whether it would show intraspecific variations within C. petrinus, judging therefore that a more in-depth description is necessary.[4] Subsequent studies carried out on the genus Cymbospondylus nevertheless always refer specimen UCMP 9913 to C. petrinus,[21] but still mentioning some notable differences.[11]
In 2011, a notable new Cymbospondylus fossil was also discovered in the Augusta Mountains, then exhumed three years later in 2014. This discovery was a partial skeleton that was briefly mentioned in a 2013 study describing the large contemporary ichthyosaur Thalattoarchon,[22] as well as in a 2018 histological study, where it is among the specimens analyzed.[23] However, it was only later that the specimen, cataloged as LACM DI 158109, was formally designated a holotype for the species C. duelferi by Nicole Klein and colleagues in 2020. The species name duelferi was chosen in honor of Olaf Dülfer, fossil preparer who made many practical contributions to research on Mesozoic marine reptiles.[11][5]
In 1998, still in the Augusta Mountains, Sander discovered another notable specimen of Cymbospondylus, and exhumed it with his colleagues between 2014 and 2015.[21] After preparation of the fossils, the specimen, cataloged as LACM DI 157871, consists of a large complete skull, some cervical vertebrae, the right humerus as well as fragments of the shoulder girdle. It was in 2021, just one year after the identification of C. duelferi, that a new species of the genus was named from this specimen in Science Magazine by Sander and his colleagues.[12] This species, C. youngorum, is named in honor of Tom and Bonda Young,[12] these latter having financially supported the fossil exhumation project.[24] However, in their main article, the anatomical description of this taxon is limited and is mainly focused on the evolution of the size of the oldests ichthyosaurs,[12] the detailed anatomical descriptions being documented in a secondary article.[21]
Formerly assigned species
Although many valid and distinct species have been assigned to Cymbospondylus throughout its taxonomic history, some of these have been reassigned to different genera or are considered synonymous or even doubtful.[4] In his 1868 paper describing Cymbospondylus, Leidy also named another ichthyosaur as Chonespondylus grandis, based on a fragment of a caudal vertebra found at Star Canyon in the Humboldt Range.[1] In 1902, Merriam listed Leidy's discoveries, but having found no features distinguishing Chonespondylus from Cymbospondylus, he decided to synonymize the first name with the second, under the name C. (?) grandis.[6] In 1908, after the discovery of new very complete fossils from C. petrinus, Merriam decided to definitively synonymize C. (?) grandis with the latter.[25][4] In 1873, John Whitaker Hulke described a species of Ichthyosaurus, I. polaris, named after two sets of vertebrae associated with rib fragments that were discovered on Isfjorden, Spitsbergen, an island in Norway.[26] In 1902, the Russian paleontologist Nikolai Nikolajewitch Yakowlew moved the species within the genus Shastasaurus, referring an isolated vertebra to this taxon.[27] In 1908, Merriam in turn moved this species into the genus Cymbospondylus, under the name C. (?) polaris. Merriam still expresses some hesitation about this attribution, asserting that the true generic identity cannot be determined for this species due to the few known fossils.[28] In 1910, the species was moved to the newly erected genus Pessosaurus by Carl Wiman, as P. polaris,[29] to which it has always been referred by this name ever since. Although this taxon is declared as a nomen dubium according to studies published at the end of the 20th century, it is seen as a species inquirenda according to McGowan and Motani in 2003, i.e. a taxon under investigation, as numerous fossils that have since been referred to P. polaris could make it once again as valid.[30]
Still in his 1908 work, Merriam erects two new species of the genus, coming from the same locality from which C. petrinus is known. The first is C. nevadanus, named from fossils constituting a hind limb. Merriam distinguishes this species from C. petrinus on the basis of its larger size and the different proportions of some bones.[31] However, the C. nevadanus material is not sufficiently diagnostic to support the validity of this species, and is considered a species inquireda according to McGowan and Motani in 2003.[32][4] The second species erected by Merriam is C. natans, which he names from an isolated humerus, to which he attributes a radius, an ulna, carpals and a series of caudal vertebrae. In his article, he notes the similarity of these bones with those of Mixosaurus,[33] leading the author to rename the species to M (?) natans in 1911.[34] For much of the 20th century, M (?) natans was recognized as a valid species of Mixosaurus until 1999, when it was synonymized with M. nordenskioeldii.[35] Although M. nordenskioeldii itself has been considered a nomen dubium since 2005,[36] the fossil material concerned remains attributed to the family Mixosauridae and is no longer attributable to Cymbospondylus.[4]
In a review of German ichthyosaurs published in 1916, Friedrich von Huene described two other species of Cymbospondylus whose fossils were discovered in the Muschelkalk of the town of Laufenburg in the state of Baden-Württemberg. The first is C. germanicus, which Huene names from a single vertebra associated with other vertebrae as well as a basioccipital.[37] Immediately afterwards, the validity of C. germanicus was questioned the same year by Ferdinand Broili, the latter citing that the fossils concerned did not present notable features to be recognized as distinct.[38][4] Additionally, the fossils appear to be very poorly preserved to be distinguished as a valid species,[3] and is therefore a nomen dubium.[39][40]

In 2002, paleontologists Chun Li and Hai-Lu You named a new species as C. asiaticus based on a complete skull discovered in the Xiaowa Formation, located in Guizhou Province, China, and it was considered as the most recent known representative of the genus.[41] In the official description of C. nichollsi published in 2006, the authors are doubtful regarding the attribution of this species to Cymbospondylus. They mention that the latter does not share any notable commonalities with the three then recognized species of the genus at the time, namely C. petrinus, C. buchseri and C. nichollsi, and suggest that it would in fact be a junior synonym of Guizhouichthyosaurus tangae.[4] The synonymy proposed by Fröbisch and colleagues was accepted in 2009 by Qing-Hua Shang and Li after the discovery of an almost complete skeleton of Guizhouichthyosaurus from the same formation. However, considering that Guizhouichthyosaurus is similar to Shastasaurus, they moved the species as S. tangae.[42] This synonymy was contested the following year, in 2010, in which Michael W. Maisch provisionally reclassified Guizhouichthyosaurus as a distinct genus.[43] In 2011, Sander and his colleagues considered that Guizhouichthyosaurus was distinct,[44] while a 2013 study by Shang and Li still synonymizes it with Shastasaurus.[45] However, numerous phylogenetic and morphological analyzes subsequently published consider Guizhouichthyosaurus to be distinct genus from Shastasaurus.[46][47][48][49]
Description

Ichthyosaurs like Cymbospondylus are marine reptiles, with two pairs of flippers for limbs and a fin on the tail.[50][51] Being one of the oldest and most basal (least derived) ichthyosaurs known from the fossil record, Cymbospondylus has an anatomy very different from those of the parvipelvians, the latter presenting the archetypal anatomy of this group of this now extinct marine reptiles (long and thin snout, large eye sockets and a homocercal caudal fin). The anatomy of Cymbospondylus, like other non-parvipelvians ichthyosaurs, is more "primitive", having a poorly developed or absent dorsal fin as well a less pronounced tail.[52] In its overall anatomy, Cymbospondylus has a long trunk and a hypocercal fin.[51][53] The anatomy of this type of ichthyosaur is similar to those of other marine vertebrates such as mosasaurs, marine crocodylomorphs, and archaeocete whales through convergent evolution.[54]
Size

The size of Cymbospondylus varies greatly between recognized species.[4][11][21] Possessing a skull which would measure a total of 65–70 cm (26–28 in), C. duelferi is the smallest known species of the genus, having a size estimated at 4.3 m (170 in) long in the study officially describing it,[11] before being estimated at 5 m (200 in) in the secondary article by Sander and colleagues in 2021.[21] The estimated size of the C. buchseri holotype is slightly larger, reaching a length of 5.5 m (220 in) with a skull 68 cm (27 in) long.[3][49] Estimating the size of C. nichollsi is more complex, because although the holotype specimen is known from a significant portion of the skeleton, the latter preserves only half of the skull. However, based on the comparison with C. buchseri and C. petrinus, the total length of the skull is estimated at 97 cm (38 in) for an animal measuring 7.6 m (300 in) long.[4][11][21]
Estimates of the size of C. petrinus have changed relatively little since 1908, mainly due to the almost complete skeleton of specimen UCMP 9950, considered the best known specimen of Cymbospondylus.[55] Merriam suggests that C. petrinus would reach a size exceeding 9.1 m (360 in) in length based on specimens UCMP 9947 and 9950.[56][3][4] In 2020, Klein and colleagues increased the size estimate of C. petrinus a little further, seeing them as reaching 9.3 m (370 in) for a 1.16 m (46 in) skull.[11] In the secondary article published by Sander and colleagues in 2021, C. petrinus is estimated to be 12.56 m (494 in) long for the same cranial measurements.[21] The holotype specimen of C. youngorum having a skull measuring almost 2 m (79 in) long and its humerus being the second largest bone of this type recorded in an ichthyosaur, the maximum size of the animal is therefore estimated at 17.65 m (57.9 ft) for a weight of 44.7 metric tons (49.3 short tons).[lower-alpha 3] This estimate not only makes C. youngourum one of the largest ichthyosaurians identified to date behind Shastasaurus sikanniensis, but also makes it one of the largest animals known of its time.[12] The antiquity as well as such an imposing size for an animal dating from the beginning of the Middle Triassic makes C. youngorum qualify as "the first aquatic giant" according to Lene Liebe Delsett and Nicholas David Pyenson.[24]
Evolution
Of particular interest is the rapid increase in body size undergone by ichthyosaurs early in their evolutionary history. Ichthyosaurs evolved from small (skull length 55 mm) ancestors such as Cartorhynchus to giant forms like C. youngorum in the span of only 2.5 million years. Cetaceans meanwhile, which originated under similar conditions (following the mass extinction at the end of the Cretaceous period eradicating much of marine tetrapod diversity) took notably longer to obtain similar sizes. Between Pakicetus (skull width 127 mm) and Basilosaurus isis (skull width 600 mm) a total of 10 to 14 million years passed, almost five times as much as between Cartorhynchus and Cymbospondylus youngorum. In toothed whales, the size increase likewise took notably longer with around 25 million years passing between the basal Simocetus and the massive Livyatan. In their 2021 study Sander, Griebeler, Schmiz and their colleagues determined that ichthyosaurs underwent rapid size increase early on in their evolutionary history, in particular among Cymbospondylus and early merriamosaurs, while whales obtained great sizes relatively slowly. Even when restricting the analysis to only observe the growth in members of Pelagiceti (fully aquatic whales) to account for the varying degrees of aquatic adaptations, cetaceans still reached large body sizes much slower than ichthyosaurs. Meanwhile, after this initial burst the overall size of ichthyosaurs slowed down significantly, while among crown group whales body sizes increased initially slowly before accelerating rapidly later in their history. Sander et al. (2021) suggests that this rapid increase in body size may have been favored by the swift recovery of conodonts and ammonites following the Permian–Triassic extinction event. They further suggest that the evolution of large eyes, a trait present in many ichthyosaurs, further assisted them in exploiting this food source efficiently.[12][57]
Classification
The exact placement of Cymbospondylus within Ichthyosauria is poorly understood with its position varying between different studies, sometimes being recovered as more and sometimes as less derived than mixosaurids.[58] However it is agreed upon that Cymbospondylus is a rather basal member of the clade. Early phylogenies placed Cymbospondylus within Shastasauridae.[59] In the analysis of Bindellini et al. (2021), Cymbospondylus is placed at the very base of Ichthyosauria, outside the more derived members of Hueneosauria (including Mixosauridae and Shastasauridae).[60] In the publication describing C. duelferi, Klein and colleagues recovered that all species from the Fossil Hill Member in Nevada form a clade with one another.[11] The description of C. youngorum further supports this Nevadan clade, recovering C. youngorum as its most derived member while C. buchseri from Europe sits at the base of the genus. Much like in the analysis by Bindellini and colleagues, shastasaurids and mixosaurids were recovered as more derived ichthyosaurs.[12]
Like in many analyses prior, the type species was not included in the dataset due to its questionable and fragmentary nature.[12] This causes Cymbospondylus to have a very convoluted taxonomy, with it being suggested that the type species should be neglected.[4][11] The 2020 study reviewed the skull morphology of C. nichollsi and found the species to be valid, as the skull morphology accords with that of C. petrinus but is distinct enough to be separate, such as the upper temporal fenestra shape being oval in C. nichollsi but triangular in C. petrinus. In their phylogenetic analysis the authors did not recover a definite placement for C. buchseri, leading them to state that further study was needed to determine whether the Swiss species belonged to the genus.[11]
The following cladogram shows the position of Cymbospondylus within the Ichthyosauria after Sander et al., (2021):[12][21]
| Ichthyosauromorpha |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Palaeobiology
Paleoecology
Massare & Callaway (1990) propose that many Triassic ichthyosaurs including Cymbospondylus may have been ambush predators. They argue that the long neck and torso would create drag in water while the laterally-flattened tail lacking the lunate fluke of later ichthyosaur taxa was more suited for an undulating swimming style. In their research they suggest that the elongated flexible bodies of early ichthyosaurs were built to support an undulating swimming style while the powerful tail would provide bursts of speed, both of which they cite as being possible adaptations to ambush prey. Massare & Callaway put this in contrast with Jurassic taxa, known for their compact, dolphin-like bodies adapted for more continuous swimming favorable to pursuit predators.[61] A strikingly similar bauplan was later obtained by two other large bodied marine amniote groups, mosasaurs and archaeocete whales.
Direct evidence for its diet exists for the medium-sized Cymbospondylus buchseri from Switzerland, which was found with its stomach contents exclusively consisting of hooks belonging to soft-bodied coleoid cephalopods. However, this does not exclude the possibility that C. buchseri could have taken larger prey, as its last meal may not reflect its typical diet accurately. Bindellini and colleagues suggest that C. buchseri may have employed a more forceful feeding strategy with a slower feeding cycle and a higher biteforce, supported by the animal's robust rostrum. In the Besano Formation, Cymbospondylus would have coexisted with two other smaller ichthyosaurs, the more gracile skulled Besanosaurus and small mixosaurs. Whether or not C. buchseri would have gone after large vertebrate prey, all three taxa display clear adaptations for different hunting strategies and prey preferences, however the details of their ecologies are not yet fully understood.[60]
For C. youngorum a generalist diet of squid and fish is inferred based on the blunt and conical teeth in combination with the elongated rostrum. However, as with C. buchseri, Sander et al. entertain the possibility that C. youngorum could have fed on large-bodied vertebrates as well, including the other Cymbospondylus species of the region.[12]
Cymbospondylus was especially diverse in the Fossil Hill Member of the Favret and Prida formations of Nevada, which preserves a pelagic environment from the middle to late Anisian (ca. 246 - 242 million years ago). The Fossil Hill Member specifically preserves some of the greatest ichthyosaur diversity recorded, with eight distinct taxa present. This includes four species of Cymbospondylus, the early macropredatory Thalattoarchon, the small Phalarodon fraasi and P. callawayi as well as the poorly understood Omphalosaurus. Other animals present in the environment include a single sauropterygian, Augustasaurus hagdorni, halobiid bivalves but only few remains of fish. However this may be due to preservation bias. Despite this abundance of amniote fossils, analysis of the preserved ecosystem indicates that the Fossil Hill Member represents a stable food web largely dominated by said amniotes, occupying niches held by fish in modern marine ecosystems. Furthermore, the analysis conducted by Eva Maria Griebeler in the study of Sander and colleagues indicates the presence of an energy surplus, which may indicate the presence of an as of yet unknown animal in the Fossil Hill fauna or that this energy surplus may have been exploited by animals that evolved later. The ecosystem could have supported another giant marine reptile, assuming it were be a bulk feeder specialised on small prey that's lower in the food chain. Such an animal could have for example been a filter feeder, a niche not yet occupied by any animal in the fauna of the Fossil Hill Member.[12]
Bindellini and colleagues notes that shastasaurid diversity may have profited from the extinction of Cymbospondylus, such as the Carnian of China, known to have supported three ecologically different shastasaurids but no examples of cymbospondylids, which had gone extinct by that time.[60]
Reproduction
The holotype of C. duelferi preserves three small strings of articulated vertebrae located within the trunk region of the specimen.[11] These vertebrae, which are only a third the size of the adult specimen, have been interpreted to represent the remains of three fetuses, with one specimen specifically facing towards the rear end of the putative mother. Following this interpretation, Cymbospondylus would have given live birth to a minimum of three offspring.
Notes
- Leidy gives a slightly different description of the holotype in his 1868 paper, stating that it had only four dorsal vertebrae, due to the preservation of the fossil block.[1]
- This specimen is one of the several additional C. petrinus fossils described by Meriam in 1908.[20]
- Sander and colleagues estimate animal size based on the 95 % prediction interval in C. youngorum. Two additional estimates were also made by the team. The prediction interval less than 95 % gives a length of 12.5 m (41 ft) for a weight of 14.7 metric tons (16.2 short tons), while that which is greater than 95 % gives a length of 25 m (82 ft) for a weight of 135.8 metric tons (149.7 short tons).[12] As estimates do not come close to 95 %, these measurements are not considered to be the probable maximum sizes of C. youngorum.[21]
References
- Joseph Leidy (1868). "Notice of some reptilian remains from Nevada". Proceedings of the Philadelphia Academy of Sciences. 20: 177–178.
- Merriam 1908, p. 123.
- P. Martin Sander (1989). "The large ichthyosaur Cymbospondylus buchseri, sp. nov., from the Middle Triassic of Monte San Giorgio (Switzerland), with a survey of the genus in Europe". Journal of Vertebrate Paleontology. 9 (2): 163–173. doi:10.1080/02724634.1989.10011750. JSTOR 4523251. S2CID 128403865.
- Nadia Fröbisch; P. Martin Sander; Olivier Rieppel (2006). "A new species of Cymbospondylus (Diapsida, Ichthyosauria) from the Middle Triassic of Nevada and a re-evaluation of the skull osteology of the genus". Zoological Journal of the Linnean Society. 147 (4): 515–538. doi:10.1111/j.1096-3642.2006.00225.x. S2CID 84720049.
- "Cymbospondylus". Paleofile.
- John C. Merriam (1902). "Triassic Ichthyopterygia from California and Nevada". University of California Publications. Bulletin of the Department of Geology. 3: 63–108.
- Merriam 1908, p. 5-6.
- Merriam 1908, p. 65-70.
- Merriam 1908, p. 104-122.
- Michael W. Maisch; Andreas T. Matzke (2004). "Observations on Triassic ichthyosaurs. Part XIII: New data on the cranial osteology of Cymbospondylus petrinus (LEIDY, 1868) from the Middle Triassic Prida Formation of Nevada". Neues Jahrbuch für Geologie und Paläontologie - Monatshefte. 2004 (6): 370–384. doi:10.1127/njgpm/2004/2004/370. S2CID 221243435.
- Nicole Klein; Lars Schmitz; Tanja Wintrich; P. Martin Sander (2020). "A new cymbospondylid ichthyosaur (Ichthyosauria) from the Middle Triassic (Anisian) of the Augusta Mountains, Nevada, USA". Journal of Systematic Palaeontology. 18 (14): 1167–1191. doi:10.1080/14772019.2020.1748132. S2CID 219078178.
- P. Martin Sander; Eva Maria Griebeler; Nicole Klein; Jorge Velez Juarbe; Tanja Wintrich; Liam J. Revell; Lars Schmitz (2021). "Early giant reveals faster evolution of large body size in ichthyosaurs than in cetaceans". Science. 374 (6575): eabf5787. doi:10.1126/science.abf5787. PMID 34941418. S2CID 245444783.
- Merriam 1908, p. 123-124.
- McGowan & Motani 2003, p. 65-66.
- Andrzej S. Wolniewicz (2017). The Anatomy, Taxonomy and Systematics of Middle Triassic–Early Jurassic Ichthyosaurs (Reptilia: Ichthyopterygia) and the Phylogeny of Ichthyopterygia (PhD). University of Oxford.
- International Commission on Zoological Nomenclature (2012). "Article 8. What constitutes published work". International Code of Zoological Nomenclature (4th ed.). Retrieved 16 July 2021.
- Mike Taylor (8 June 2010). "Notes on Early Mesozoic Theropods and the future of zoological nomenclature". Sauropod Vertebra Picture of the Week. Archived from the original on 9 March 2021.
- Bernhard Peyer (1944). "Die Reptilien vom Monte San Giorgio". Neujahrsblau der Naturforschcnden Gesellschaft in Zürich (in German). 146: 1–95.
- Emil Kuhn-Schnyder (1964). "Die Wirbellierfauna der Tessiner Kalkalpen". Geologische Rundschau (in German). 53: 393–412. doi:10.1007/BF02040759. S2CID 129726404.
- Merriam 1908, p. 105-110.
- P. Martin Sander; Eva Maria Griebeler; Lars Schmitz (2021). "Supplementary Materials for Early giant reveals faster evolution of large body size in ichthyosaurs than in cetaceans" (PDF). Science.
- Nadia B. Fröbisch; Jörg Fröbisch; P. Martin Sander; Lars Schmitz; Olivier Rieppel (2013). "Macropredatory ichthyosaur from the Middle Triassic and the origin of modern trophic networks". Proceedings of the National Academy of Sciences of the United States of America. 110 (4): 1393–1397. Bibcode:2013PNAS..110.1393F. doi:10.1073/pnas.1216750110. PMC 3557033. PMID 23297200.
- Alexandra Houssaye; Yasuhisa Nakajima; P. Martin Sander (2018). "Structural, functional, and physiological signals in ichthyosaur vertebral centrum microanatomy and histology". Geodiversitas. 40 (2): 161–170. doi:10.5252/geodiversitas2018v40a7. S2CID 56134834.
- Lene Liebe Delsett; Nicholas D. Pyenson (2021). "Early and fast rise of Mesozoic ocean giants". Science. 374 (6575): 1554–1555. Bibcode:2021Sci...374.1554D. doi:10.1126/science.abm3751. PMID 34941421. S2CID 245456946.
- Merriam 1908, p. 104.
- John W. Hulke (1873). "Memorandum on some fossil vertebrate remains collected by the Swedish expeditions to Spitzbergen in 1864 and 1868". Bihang Till Kongliga Svenska Vetenskaps-Akademiens Handlingar. 1 (9): 1–11.
- Nikolai N. Yakowlew (1902). "Neue funde von Trias-Sauriern auf Spitzbergen". Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg (in German). 40: 179–202.
- Merriam 1908, p. 149-150.
- Carl Wiman (1910). "Ichthyosaurier aus der Trias Spitzbergens" (PDF). Bulletin of the Geological Institutions of the University of Uppsala (in German). 10: 124–148.
- McGowan & Motani 2003, p. 128.
- Merriam 1908, p. 124-127.
- McGowan & Motani 2003, p. 125.
- Merriam 1908, p. 150-152.
- John C. Merriam (1911). "Notes on the relationships of the marine saurian fauna described from the Triassic of Spitzbergen by Wiman". University of California Publications. Bulletin of the Department of Geology. 6 (13): 317–327.
- Elizabeth L. Nicholls; Donald B. Brinkman; Jack M. Callaway (1999). "New Material of Phalarodon (Reptilia: Ichthyosauria) from the Triassic of British Columbia and its Bearing on the Interrelationships of Mixosaurs". Palaeontographica Abteilung A. 252 (1): 1–22. doi:10.1127/pala/252/1998/1. S2CID 246923904.
- Lars Schmitz (2005). "The taxonomic status of Mixosaurus nordenskioeldii (Ichthyosauria)". Journal of Vertebrate Paleontology. 25 (4): 983–985. doi:10.1671/0272-4634(2005)025[0983:TTSOMN]2.0.CO;2. JSTOR 4524525.
- Friedrich von Huene (1916). "Beiträge zur Kenntnis der Ichthyosaurier im deutschen Muschelkalk". Palaeontographica (in German). 68: 1–68.
- Von F. Broili (1916). "Einige Bemerkungen über die Mixosauridae". Anatomischer Anzeiger (in German). 49: 474–494.
- Jack M. Callaway; Judith A. Massare (1989). "Geographic and stratigraphic distribution of the Triassic Ichthyosauria (Reptilia; Diapsida)". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 178: 37–58.
- McGowan & Motani 2003, p. 129.
- Chun Li; Hai-Lu You (2002). "Cymbospondylus from the Upper Triassic of Guizhou, China" (PDF). Vertebrata PalAsiatica (in Chinese and English). 40: 9–16.
- Qing-Hua Shang; Chun Li (2009). "On the occurrence of the ichthyosaur Shastasaurus in the Guanling biota (Late Triassic), Guizhou, China" (PDF). Vertebrata PalAsiatica (in Chinese and English). 47 (3): 178–193.
- Michael W. Maisch (2010). "Phylogeny, systematics, and origin of the Ichthyosauria – the state of the art" (PDF). Palaeodiversity. 3: 151–214.
- P. Martin Sander; Xiaohong Chen; Long Cheng; Xiaofeng Wang (2011). "Short-Snouted Toothless Ichthyosaur from China Suggests Late Triassic Diversification of Suction Feeding Ichthyosaurs". PLOS ONE. 6 (5): e19480. Bibcode:2011PLoSO...619480S. doi:10.1371/journal.pone.0019480. PMC 3100301. PMID 21625429.
- Qing-Hua Shang; Chun Li (2013). "On the sexual dimorphism of Shastasaurus tangae (Reptilia: Ichthyosauria) from the Triassic Guanling Biota, China" (PDF). Vertebrata PalAsiatica (in Chinese and English). 51 (4): 253–264.
- Cheng Ji; Da-Yong Jiang; Ryosuke Motani; Olivier Rieppel; Wei-Cheng Hao; Zuo-Yu Sun (2016). "Phylogeny of the Ichthyopterygia incorporating recent discoveries from South China". Journal of Vertebrate Paleontology. 36 (1): e1025956. Bibcode:2016JVPal..36E5956J. doi:10.1080/02724634.2015.1025956. S2CID 85621052.
- Benjamin C. Moon (2019). "A new phylogeny of ichthyosaurs (Reptilia: Diapsida)" (PDF). Journal of Systematic Palaeontology. 17 (2): 129–155. doi:10.1080/14772019.2017.1394922. hdl:1983/463e9f78-10b7-4262-9643-0454b4aa7763. S2CID 90912678.
- Da-Yong Jiang; Ryosuke Motani; Andrea Tintori; Olivier Rieppel; Cheng Ji; Min Zhou; Xue Wang; Hao Lu; Zhi-Guang Li (2020). "Evidence supporting predation of 4-m marine reptile by Triassic megapredator". iScience. 23 (9): 101347. Bibcode:2020iSci...23j1347J. doi:10.1016/j.isci.2020.101347. PMC 7520894. PMID 32822565.
- Gabriele Bindellini; Andrzej S. Wolniewicz; Feiko Miedema; Torsten M. Scheyer; Cristiano Dal Sasso (2021). "Cranial anatomy of Besanosaurus leptorhynchus Dal Sasso & Pinna, 1996 (Reptilia: Ichthyosauria) from the Middle Triassic Besano Formation of Monte San Giorgio, Italy/Switzerland: taxonomic and palaeobiological implications". PeerJ. 9: e11179. doi:10.7717/peerj.11179. PMC 8106916. PMID 33996277.
- P. Martin Sander (2000). "Ichthyosauria: their diversity, distribution, and phylogeny". Paläontologische Zeitschrift. 74 (1): 1–35. doi:10.1007/BF02987949. S2CID 85352593.
- Ryosuke Motani (2005). "Evolution of Fish-Shaped Reptiles (reptilia: Ichthyopterygia) in Their Physical Environments and Constraints" (PDF). Annual Review of Earth and Planetary Sciences. 33: 395–420. Bibcode:2005AREPS..33..395M. doi:10.1146/annurev.earth.33.092203.122707. S2CID 54742104. Archived from the original (PDF) on 23 December 2018.
- Ryan Marek (2015). "Fossil Focus: Ichthyosaurs". Palaeontology Online. 5: 8. Archived from the original on 18 January 2021. Retrieved 13 June 2020.
- Judith M. Pardo-Pérez; Benjamin P. Kear; Erin E. Maxwell (2020). "Skeletal pathologies track body plan evolution in ichthyosaurs". Scientific Reports. 10 (1): 4206. Bibcode:2020NatSR..10.4206P. doi:10.1038/s41598-020-61070-7. PMC 7060314. PMID 32144303.
- Johan Lindgren; Michael W. Caldwell; Takuya Konishi; Luis M. Chiappe (2010). "Convergent Evolution in Aquatic Tetrapods: Insights from an Exceptional Fossil Mosasaur". PLOS ONE. 5 (8): e11998. Bibcode:2010PLoSO...511998L. doi:10.1371/journal.pone.0011998. PMC 2918493. PMID 20711249.
- McGowan & Motani 2003, p. 66.
- Merriam 1908, p. 105.
- Delsett, L. L.; Pyenson, N. D. (2021). "Early and fast rise of Mesozoic ocean giants". Science. 374 (6575): 1554–1555. doi:10.1126/science.abm3751. PMID 34941421. S2CID 245456946.
- Michael W. Maisch (2010). "Phylogeny, systematics, and origin of the Ichthyosauria – the state of the art" (PDF). Palaeodiversity. 3: 151–214.
- Ryosuke Motani (1999). "Phylogeny of the Ichthyopterygia" (PDF). Journal of Vertebrate Paleontology. 19 (3): 472–495. doi:10.1080/02724634.1999.10011160. JSTOR 4524011. S2CID 84536507. Archived from the original (PDF) on 5 March 2016. Retrieved 19 October 2011.
- Bindellini, G.; Wolniewicz, A.S.; Miedema, F.; Scheyer, T.M.; Dal Sasso, C. (2021). "Cranial anatomy of Besanosaurus leptorhynchus Dal Sasso & Pinna, 1996 (Reptilia: Ichthyosauria) from the Middle Triassic Besano Formation of Monte San Giorgio, Italy/Switzerland: taxonomic and palaeobiological implications". PeerJ. 9: e11179. doi:10.7717/peerj.11179. PMC 8106916. PMID 33996277.
- Massare, J.A.; Callaway, JM (1990). "The affinities and ecology of Triassic ichthyosaurs". Geological Society of America Bulletin. 102 (4): 409–416. doi:10.1130/0016-7606(1990)102<0409:TAAEOT>2.3.CO;2.
Bibliography
- Merriam, John C. (1908). Triassic Ichthyosauria, with special reference to the American forms. Memoirs of the University of California. Vol. 1. Berkeley, California: The University Press. pp. 1–196. OCLC 457714430.
- Maisch, Michael W.; Matzke, Andreas T. (2000). "The Ichthyosauria". Stuttgarter Beiträge zur Naturkunde, Serie B. 298: 1–159.
- McGowan, Christopher; Motani, Ryosuke (2003). Handbook of Paleoherpetology Part 8: Ichthyopterygia. Vol. 1. Munich: Verlag Dr. Friedrich Pfeil. pp. 1–175. ISBN 978-3-899-37007-2. OCLC 469848769.
External links
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||