Endosperm
The endosperm is a tissue produced inside the seeds of most of the flowering plants following double fertilization. It is triploid (meaning three chromosome sets per nucleus) in most species,[1] which may be auxin-driven.[2] It surrounds the embryo and provides nutrition in the form of starch, though it can also contain oils and protein. This can make endosperm a source of nutrition in animal diet. For example, wheat endosperm is ground into flour for bread (the rest of the grain is included as well in whole wheat flour), while barley endosperm is the main source of sugars for beer production. Other examples of endosperm that forms the bulk of the edible portion are coconut "meat" and coconut "water",[3] and corn. Some plants, such as orchids, lack endosperm in their seeds.
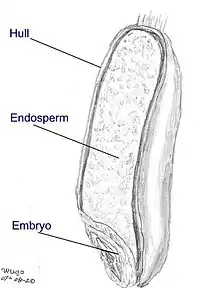
Origin of endosperm
Ancestral flowering plants have seeds with small embryos and abundant endosperm. The evolutionary development of flowering plants trends towards plants with mature seeds with little or no endosperm. In more derived flowering plants the embryo occupies most of the seed and the endosperm is non-developed or consumed before the seed matures.[4][5]
Double fertilization
Endosperm is formed when the two sperm nuclei inside a pollen grain reach the interior of a female gametophyte (sometimes called the embryo sac). One sperm nucleus fertilizes the egg cell, forming a zygote, while the other sperm nucleus usually fuses with the binucleate central cell, forming a primary endosperm cell (its nucleus is often called the triple fusion nucleus). That cell created in the process of double fertilization develops into the endosperm. Because it is formed by a separate fertilization, the endosperm constitutes an organism separate from the growing embryo.
About 70% of angiosperm species have endosperm cells that are polyploid.[6] These are typically triploid (containing three sets of chromosomes), but can vary widely from diploid (2n) to 15n.[7]
One flowering plant, Nuphar polysepala, has diploid endosperm, resulting from the fusion of a pollen nucleus with one, rather than two, maternal nuclei. The same is supposed for some other basal angiosperms.[8] It is believed that early in the development of angiosperm lineages, there was a duplication in this mode of reproduction, producing seven-celled/eight-nucleate female gametophytes, and triploid endosperms with a 2:1 maternal to paternal genome ratio.[9]
Double fertilisation is a characteristic feature of angiosperms.
Endosperm formation
There are three types of Endosperm development:
Nuclear endosperm formation – where repeated free-nuclear divisions take place; if a cell wall is formed it will form after free-nuclear divisions. Commonly referred to as liquid endosperm. Coconut water is an example of this.
Cellular endosperm formation – where a cell-wall formation is coincident with nuclear divisions. Coconut meat is cellular endosperm. Acoraceae has cellular endosperm development while other monocots are helobial.
Helobial endosperm formation – Where a cell wall is laid down between the first two nuclei, after which one half develops endosperm along the cellular pattern and the other half along the nuclear pattern.
Evolutionary origins
The evolutionary origins of double fertilization and endosperm are unclear, attracting researcher attention for over a century. There are the two major hypotheses:[7]
- The double fertilization initially used to produce two identical, independent embryos ("twins"). Later these embryos acquired different roles, one growing into the mature organism, and another merely supporting it. Thus, the early endosperm was probably diploid, like the embryo. Some gymnosperms, such as Ephedra, may produce twin embryos by double fertilization. Either of these two embryos is capable of filling in the seed, but normally only one develops further (the other eventually aborts). Also, most basal angiosperms still contain the four-cell embryo sac and produce diploid endosperms.
- Endosperm is the evolutionary remnant of the actual gametophyte, similar to the complex multicellular gametophytes found in gymnosperms. In this case, acquisition of the additional nucleus from the sperm cell is a later evolutionary step. This nucleus may provide the parental (not only maternal) organism with some control over endosperm development. Becoming triploid or polyploid are later evolutionary steps of this "primary gametophyte". Nonflowering seed plants (conifers, cycads, Ginkgo, Ephedra) form a large homozygous female gametophyte to nourish the embryo within a seed.[10]
The triploid transition - and the production of antipodal cells - may have occurred due to a shift in gametophyte development which produced a new interaction with an auxin-dependent mechanism originating in the earliest angiosperms.[2]
The role of endosperm in seed development
In some groups (e.g. grains of the family Poaceae), the endosperm persists to the mature seed stage as a storage tissue, in which case the seeds are called "albuminous" or "endospermous", and in others it is absorbed during embryo development (e.g., most members of the family Fabaceae, including the common bean, Phaseolus vulgaris), in which case the seeds are called "exalbuminous" or "cotyledonous" and the function of storage tissue is performed by enlarged cotyledons ("seed leaves"). In certain species (e.g. corn, Zea mays); the storage function is distributed between both endosperm and the embryo. Some mature endosperm tissue stores fats (e.g. castor bean, Ricinus communis) and others (including grains, such as wheat and corn) store mainly starches.
The dust-like seeds of orchids have no endosperm. Orchid seedlings are mycoheterotrophic in their early development. In some other species, such as coffee, the endosperm also does not develop.[11] Instead, the nucellus produces a nutritive tissue termed "perisperm". The endosperm of some species is responsible for seed dormancy.[12] Endosperm tissue also mediates the transfer of nutrients from the mother plant to the embryo, it acts as a location for gene imprinting, and is responsible for aborting seeds produced from genetically mismatched parents.[6] In angiosperms, the endosperm contain hormones such as cytokinins, which regulate cellular differentiation and embryonic organ formation.[13]
Cereal grains
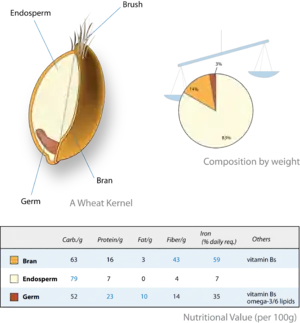
Cereal crops are grown for their edible fruit (grains or caryopses), which are primarily endosperm. In the caryopsis, the thin fruit wall is fused to the seed coat. Therefore, the nutritious part of the grain is the seed and its endosperm. In some cases (e.g. wheat, rice) the endosperm is selectively retained in food processing (commonly called white flour), and the embryo (germ) and seed coat (bran) removed. The processed grain has a lower quality of nutrition. Endosperm thus has an important role within the human diet worldwide.
The aleurone is the outer layer of endosperm cells, present in all small grains and retained in many dicots with transient endosperm. The cereal aleurone functions for both storage and digestion. During germination, it secretes the amylase enzyme that breaks down endosperm starch into sugars to nourish the growing seedling.[14][15]
See also
References
- Stewart-Cox JA, Britton NF, Mogie M (August 2004). "Endosperm triploidy has a selective advantage during ongoing parental conflict by imprinting". Proceedings. Biological Sciences. 271 (1549): 1737–43. doi:10.1098/rspb.2004.2783. PMC 1691787. PMID 15306295.
- Friedman, W. E. (2009-06-25). "Auxin at the Evo-Devo Intersection". Science. American Association for the Advancement of Science. 324 (5935): 1652–1653. Bibcode:2009Sci...324.1652F. doi:10.1126/science.1176526. ISSN 0036-8075. PMID 19556491. S2CID 206521265.
- "Edible Palm Fruits". Wayne's Word: An Online Textbook of Natural History. Archived from the original on 2 September 2017. Retrieved 14 July 2010.
- "The Seed Biology Place - Seed Dormancy". Seedbiology.de. Retrieved 2014-02-05.
- Friedman WE (1998), "The evolution of double fertilization and endosperm: an "historical" perspective", Sexual Plant Reproduction, 11: 6, doi:10.1007/s004970050114, S2CID 19785565
- Olsen O (2007). Endosperm: Developmental and Molecular Biology. ISBN 9783540712350.
- Baroux C, Spillane C, Grossniklaus U (August 2002). "Evolutionary origins of the endosperm in flowering plants". Genome Biology. 3 (9): reviews1026. doi:10.1186/gb-2002-3-9-reviews1026. PMC 139410. PMID 12225592.
- Williams JH, Friedman WE (January 2002). "Identification of diploid endosperm in an early angiosperm lineage". Nature. 415 (6871): 522–6. Bibcode:2002Natur.415..522W. doi:10.1038/415522a. PMID 11823859. S2CID 4396197.
- Friedman WE, Williams JH (February 2003). "Modularity of the angiosperm female gametophyte and its bearing on the early evolution of endosperm in flowering plants". Evolution; International Journal of Organic Evolution. 57 (2): 216–30. doi:10.1111/j.0014-3820.2003.tb00257.x. PMID 12683519. S2CID 24303275.
- Friedman WE (April 1995). "Organismal duplication, inclusive fitness theory, and altruism: understanding the evolution of endosperm and the angiosperm reproductive syndrome". Proceedings of the National Academy of Sciences of the United States of America. 92 (9): 3913–7. Bibcode:1995PNAS...92.3913F. doi:10.1073/pnas.92.9.3913. PMC 42072. PMID 11607532.
- Houk WG (1938). "Endosperm and Perisperm of Coffee with Notes on the Morphology of the Ovule and Seed Development". American Journal of Botany. 25 (1): 56–61. doi:10.2307/2436631. JSTOR 2436631.
- Basra AS (1994). Mechanisms of plant growth and improved productivity: modern approaches. New York: M. Dekker. ISBN 978-0-8247-9192-6.
- Pearson LC (1995). The diversity and evolution of plants. Boca Raton: CRC Press. p. 547. ISBN 978-0-8493-2483-3.
- Becraft PW, Yi G (March 2011). "Regulation of aleurone development in cereal grains". Journal of Experimental Botany. 62 (5): 1669–75. doi:10.1093/jxb/erq372. PMID 21109580.
- Becraft PW, Gutierrez-Marcos J (2012). "Endosperm development: dynamic processes and cellular innovations underlying sibling altruism". Wiley Interdisciplinary Reviews: Developmental Biology. 1 (4): 579–93. doi:10.1002/wdev.31. PMID 23801534. S2CID 5752973.
External links
- Beach, Chandler B., ed. (1914). . . Chicago: F. E. Compton and Co.
- Endosperm: the pivot of the sexual conflict in flowering plants at Earthling Nature
| Subdisciplines | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant groups | |||||||||||
| Plant anatomy |
| ||||||||||
| Plant physiology Materials | |||||||||||
| Plant growth and habit | |||||||||||
| Reproduction | |||||||||||
| Plant taxonomy | |||||||||||
| Practice | |||||||||||
| |||||||||||