Highly branched isoprenoid
Highly branched isoprenoids (HBIs) are long-chain alkenes produced by a small number of marine diatoms.[1][2][3] There are a variety of highly branched isoprenoid structures, but C25 Highly branched isoprenoids containing 1 to 3 double bonds are the most common in the sedimentary record. Highly branched isoprenoids have been used as environmental proxies for sea ice conditions in the Arctic and Antarctic throughout the Holocene, and more recently, are being used to reconstruct more ancient ice records.[4]
Background
Highly branched isoprenoids are a type of lipid produced by marine diatoms. Highly branched isoprenoids are biomarkers, and their presence/absence in sedimentary and ice records can be used as a direct proxy for the presence of sea ice. Generally, the highly branched isoprenoids that are used as sea ice proxies are 25 carbon molecules containing 1 - 3 double bonds. The longest carbon chains in the C25 highly branched isoprenoids used for sea ice reconstructions are 15 carbons, but these molecules are highly branched and have shorter carbon chains attached to the primary carbon chain. There are 3 C25 highly branched isoprenoids used as ice proxies: a C25 monoene (HBI I), a C25 diene (HBI II), and a C25 triene (HBI III). Highly branched isoprenoid I and II are unique in that they are primarily produced by sympagic diatoms. Sympagic diatoms live in channels at the base of sea ice, making them a highly accurate proxy for sea ice. During the spring, highly branched isoprenoids are produced by diatoms in the sea ice.[5] In the summer, the ice melts, releasing the highly branched isoprenoids into the water column, where it sinks and is then deposited in the sediments.[5] Highly branched isoprenoids I and II are generally absent from regions which experience no sea ice cover, supporting their use as a proxy for seasonal sea ice. Highly branched isoprenoid III is produced by pelagic algae, or algae that thrives in the open ocean.[6] Highly branched isoprenoid III can be used as a biomarker for seasonal sea ice in the open ocean.[5]
Highly branched isoprenoids were first discovered in 1976 by Patrick Gearing in sediments in the Gulf of Mexico off the coast of Florida in a survey of hydrocarbons in shelf sediments.[7] Following this initial identification, highly branched isoprenoids were identified in a variety of marine environments, such as in the Puget Sound, Antarctica, Spain, and Peru.[1] C25 Highly branched isoprenoids were first identified in marine diatoms by John Volkman in 1994, when he isolated seven different C25 highly branched isoprenoids from the marine diatom Haslea ostrearia.[1] This provided initial evidence that highly branched isoprenoids are produced by marine diatoms.
Currently, the precise biological functions of highly branched isoprenoids are not well-understood. Highly branched isoprenoids are a type of isoprenoid lipid, which have a variety of vital biological functions. Isoprenoids are important in regulating gene expression, making up cell membranes, and are important in electron transport and photosynthesis.[8]
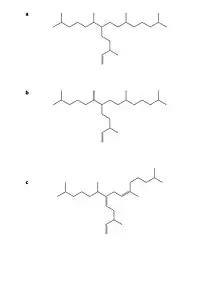
Highly branched isoprenoid I
| Names | |
|---|---|
| IUPAC name
2,6,10,14-tetramethyl-7-(3-methylpent-4-enyl)pentadecane | |
| Identifiers | |
3D model (JSmol) |
|
| ChemSpider | |
PubChem CID |
|
| |
| |
| Properties | |
| C25H50 | |
| Molar mass | 350.675 g·mol−1 |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
Infobox references | |
Highly branched isoprenoid (HBI I), a C25 monoene, is also known as IP25 (ice proxy with 25 carbon atoms). IP25 serves as a biomarker for ice conditions in the Arctic.[4] This highly branched isoprenoid is characterized by a single double bond and was first identified in marine diatoms by Thomas Brown in 2014.[3] A variety of diatoms have been shown to produce IP25, with the majority of highly branched isoprenoids produced by the Arctic diatoms Haslea crucigeroides, Haslea spicula, Haslea kjellmanii, and Pleurosigma stuxbergii var. rhomboids.[3] Despite the fact that these species do not comprise much of the sympagic diatom communities globally, they are common in the Arctic.[3] To date, IP25 has been identified in over 500 Arctic samples.[5]
Highly branched isoprenoid II
| Names | |
|---|---|
| IUPAC name
2,10,14-trimethyl-6-methylidene-7-(3-methylpent-4-enyl)pentadecane | |
| Identifiers | |
3D model (JSmol) |
|
| ChemSpider | |
PubChem CID |
|
| |
| |
| Properties | |
| C25H48 | |
| Molar mass | 348.659 g·mol−1 |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
Infobox references | |
Highly branched isoprenoid II (HBI II), a C25 diene, is also known as IPSO25 (ice proxy for the Southern Ocean with 25 carbon atoms). IPSO25 is a biomarker proxy for paleo ice in the Southern Ocean. IPSO25 has also been found to co-occur with IP25 in the Arctic.[3] HBI II contains two double bonds and was first identified in marine diatoms by Simon Belt in 2016.[9] The primary source of IPSO25 the sympagic diatom Berkeleya adeliensis, which lives within platelet ice.[9] The IPSO25 proxy is a less-developed biomarker than IP25, and its Arctic sources are unclear. IPSO25 has also been identified in the diatom Haslea ostrearia and in sediments in non-polar locations, indicating that more work is needed to fully understand and develop IPSO25 as a paleo ice proxy.[10][11]
Highly branched isoprenoid III
| Names | |
|---|---|
| IUPAC name
(Z,9E)-2,6,10,14-tetramethyl-9-(3-methylpent-4-enylidene)pentadec-6-ene | |
| Identifiers | |
3D model (JSmol) |
|
| ChemSpider | |
PubChem CID |
|
| |
| |
| Properties | |
| C25H46 | |
| Molar mass | 346.643 g·mol−1 |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
Infobox references | |
Highly branched isoprenoid III (HBI III), a C25 triene, is a biomarker useful for the analysis of the marginal ice zone (MIZ), a zone between the open ocean and sea ice. Highly branched isoprenoid III is primarily produced by pelagic algae of the genus Rhizosolenia, particularly Rhizsolenia setigera, Rhizosolenia herbetata f. semispina, and Rhizosolenia polydactyla.[6] Its source was determined by Simon Belt in 2017, who isolated highly branched isoprenoid III from phytoplankton samples from western Svalbard and the South Atlantic.[6] The production of highly branched isoprenoid III appears to be enhanced in the MIZ, however why this occurs is currently not well-understood.[5] The absence of highly branched isoprenoid III in sediments is typically attributed to sea ice cover in the region, given that ice cover would not allow for pelagic algae production.[5]
Preservation
Many biomarkers possess functional groups or are unsaturated, causing them to undergo diagenesis when emplaced in sediments. Highly branched isoprenoids are observed to be well-preserved in the sedimentary record.[5] Highly branched isoprenoids primarily form in the seasonal sea ice during the spring sympagic diatom blooms. When these ice sheets subsequently melt, the highly branched isoprenoids formed in the sea ice are released into the sea water. They then fall through the water column and are emplaced into the sediments, where they can be preserved. The sediment conditions impacts how well the highly branched isoprenoids are preserved. Generally, the stability of highly branched isoprenoids is dependent on their degree of unsaturation.[12] C25 trienes are more likely to undergo degradation compared to C25 monenes and dienes, and degradation is enhanced by increasing temperature and exposure to light, which results in oxidation.[12] The mono-unsaturated C25 IP25 is the least reactive highly branched isoprenoid, and thus is the most resistant to degradation and is best preserved in sediments.[12]
Highly branched isoprenoids, particularly highly branched isoprenoid I, are hypothesized to have long-term stability. While most studies to date have focused on studying highly branched isoprenoids in sediments from the Holocene, highly branched isoprenoids have been detected, measured, and analyzed in 12 million year old sediments from the late Miocene.[13] More work is necessary to determine whether highly branched isoprenoids can be preserved beyond 12 million years, but it is likely that their time to degradation is dependent on the local sediment conditions.
Measurement techniques
GC/MS
Highly branched isoprenoids and other organic materials can be extracted from sediments for analysis. Coupled gas chromatography and mass spectrometry can be utilized to analyze the organic materials present in the sediments. Peaks can be identified using a gas chromatograph and a mass spectrum, which provide information about the retention time and mass-to-charge ratio of the organic compounds present in the molecule. This allows for the identification of highly branched isoprenoids within a material. To identify highly branched isoprenoids in sediments, typically selective ion monitoring (SIM) is utilized.[3] SIM gathers data at masses of interest within an expected retention time window, enabling the identification and quantification of compounds with high sensitivity. Highly branched isoprenoid I, highly branched isoprenoid II, and highly branched isoprenoid III are identified and quantified via SIM using the characteristic mass spectrum peaks at m/z = 350, 348, and 346, respectively.[5]
Isotope ratios
Carbon isotopic measurements can be utilized to confirm the sea ice origin of highly branched isoprenoids. These carbon isotope measurements are obtained using an Isotope Ratio Mass Spectrometer. Highly branched isoprenoids with a sea ice origin are enriched in δ13C in both sea ice and in sediments.[5] The measurement of δ13C values enables a diagnostic determination of whether the highly branched isoprenoid came from sea ice.
Highly branched isoprenoid I
Highly branched isoprenoid I has been observed to be enriched in 13C. δ13C values range from -16.9 to -22.7‰ in sea ice and -16.3 to -23.2‰ in sediments.[15][16] This enrichment has been hypothesized to be in part a result of the fact that the marine diatoms that produce highly branched isoprenoid I live under CO2-limited conditions.[15] Temperature and diatom growth rate may also play a role in observed isotopic composition of highly branched isoprenoid I, but more work is needed to fully understand the drivers of the observed δ13C values.[15]
Highly branched isoprenoid II
Highly branched isoprenoid II has a characteristic carbon isotope composition, with measured δ13C values ranging from -5.7 to -8.5‰.[17] This value is indicative of an enrichment in 13C. This enrichment has also been observed for highly branched isoprenoid II in sediments and in waters near melting sea ice.[17][18] This enrichment likely results from the CO2-limited conditions under which the diatom producers of highly branched isoprenoid II grow.[17] This distinctive isotopic composition provides strong evidence for the sea ice origin of highly branched isoprenoid II, making it a good proxy for sea ice around Antarctica.
Highly branched isoprenoid III
Highly branched isoprenoid III is depleted in δ13C, with values ranging from -35 to -40‰.[17] Highly branched isoprenoid III is depleted in δ13C because it is not produced under CO2-limited conditions, so the depletion is a result of biological fractionation.[17]
Case study: Use of IP25 to reconstruct ice records
Currently, more than 60 paleo sea ice records that have been constructed based on IP25 over the Holocene, the Mid-Pleistocene Transition, the Pliocene/Pleistocene boundary, and the late Miocene.[19] The presence of IP25 in sediments is a direct proxy for the presence of seasonal sea ice cover. One study utilized the concentration of IP25 in sediments to reconstruct sea-ice records in the western North Pacific and Bering Sea over the past 18,000 years.[20] Researchers from Germany obtained sediment samples from the North Pacific Ocean and Bering Sea during a Sonne cruise in 2009.[20] IP25 was identified in the samples using GC/MS, and the sediments were dated using a chronostratigraphic approach, X-ray fluorescence, and radiocarbon dating of planktonic foraminifera.[20] The researchers found that variations in the concentration of the sea-ice proxy IP25 were consistent with known temperature variations based on other evidence, such as δ18O values and biogenic opal data.[20] Generally, during cold intervals, the concentration of IP25 in sediments was elevated, indicating more extensive sea ice cover.[20] More specifically, between 18,000 and 15,000 years ago, IP25 concentrations were relatively high, but decreased between 14,700 and 12,900 years ago during the Bølling/Allerød warming period.[20] At 12,500 years ago, a significant increase in IP25 concentration was detected in the sediments, consistent with the start of the Younger Dryas period, which marked a return to glacial conditions.[20] IP25 concentrations decreased to approximately 0 11,500 years ago, marking the end of the Younger Dryas. For the entire Holocene, IP25 concentrations have remained low, which is accordant with the lack of extensive ice cover throughout this period.[20] This reconstruction is consistent with other paleoclimate proxies and known climate variations, demonstrating the ability of the IP25 proxy to reconstruct paleo ice records.
References
- Volkman, John K.; Barrett, Stephanie M.; Dunstan, Graeme A. (March 1994). "C25 and C30 highly branched isoprenoid alkenes in laboratory cultures of two marine diatoms". Organic Geochemistry. 21 (3–4): 407–414. doi:10.1016/0146-6380(94)90202-X.
- Belt, Simon T; Allard, W.Guy; Massé, Guillaume; Robert, Jean-Michel; Rowland, Steven J (November 2000). "Highly branched isoprenoids (HBIs): identification of the most common and abundant sedimentary isomers". Geochimica et Cosmochimica Acta. 64 (22): 3839–3851. Bibcode:2000GeCoA..64.3839B. doi:10.1016/S0016-7037(00)00464-6.
- Brown, T. A.; Belt, S. T.; Tatarek, A.; Mundy, C. J. (September 2014). "Source identification of the Arctic sea ice proxy IP25". Nature Communications. 5 (1): 4197. Bibcode:2014NatCo...5.4197B. doi:10.1038/ncomms5197. ISSN 2041-1723. PMID 24939562.
- Belt, Simon T.; Massé, Guillaume; Rowland, Steven J.; Poulin, Michel; Michel, Christine; LeBlanc, Bernard (January 2007). "A novel chemical fossil of palaeo sea ice: IP25". Organic Geochemistry. 38 (1): 16–27. doi:10.1016/j.orggeochem.2006.09.013.
- Belt, Simon T. (November 2018). "Source-specific biomarkers as proxies for Arctic and Antarctic sea ice". Organic Geochemistry. 125: 277–298. doi:10.1016/j.orggeochem.2018.10.002. hdl:10026.1/12842. S2CID 133860206.
- Belt, Simon T.; Brown, Thomas A.; Smik, Lukas; Tatarek, Agnieszka; Wiktor, Józef; Stowasser, Gabriele; Assmy, Philipp; Allen, Claire S.; Husum, Katrine (August 2017). "Identification of C25 highly branched isoprenoid (HBI) alkenes in diatoms of the genus Rhizosolenia in polar and sub-polar marine phytoplankton". Organic Geochemistry. 110: 65–72. doi:10.1016/j.orggeochem.2017.05.007. hdl:10026.1/9642.
- Gearing, Patrick; Gearing, Juanita Newman; Lytle, Thomas F.; Lytle, Julia Sever (1976-09-01). "Hydrocarbons in 60 northeast Gulf of Mexico shelf sediments: a preliminary survey". Geochimica et Cosmochimica Acta. 40 (9): 1005–1017. Bibcode:1976GeCoA..40.1005G. doi:10.1016/0016-7037(76)90043-0.
- Holstein, Sarah A.; Hohl, Raymond J. (April 2004). "Isoprenoids: Remarkable diversity of form and function". Lipids. 39 (4): 293–309. doi:10.1007/s11745-004-1233-3. ISSN 0024-4201. PMID 15357017. S2CID 4061743.
- Belt, S. T.; Smik, L.; Brown, T. A.; Kim, J.-H.; Rowland, S. J.; Allen, C. S.; Gal, J.-K.; Shin, K.-H.; Lee, J. I.; Taylor, K. W. R. (November 2016). "Source identification and distribution reveals the potential of the geochemical Antarctic sea ice proxy IPSO25". Nature Communications. 7 (1): 12655. Bibcode:2016NatCo...712655B. doi:10.1038/ncomms12655. ISSN 2041-1723. PMC 5013606. PMID 27573030.
- Johns, L.; Wraige, E.J.; Belt, S.T.; Lewis, C.A.; Massé, G.; Robert, J.-M.; Rowland, S.J. (November 1999). "Identification of a C 25 highly branched isoprenoid (HBI) diene in Antarctic sediments, Antarctic sea-ice diatoms and cultured diatoms". Organic Geochemistry. 30 (11): 1471–1475. doi:10.1016/S0146-6380(99)00112-6.
- Xu, Yunping; Jaffé, Rudolf; Wachnicka, Anna; Gaiser, Evelyn E. (July 2006). "Occurrence of C25 highly branched isoprenoids (HBIs) in Florida Bay: Paleoenvironmental indicators of diatom-derived organic matter inputs". Organic Geochemistry. 37 (7): 847–859. doi:10.1016/j.orggeochem.2006.02.001.
- Cabedo Sanz, Patricia; Smik, Lukas; Belt, Simon T. (July 2016). "On the stability of various highly branched isoprenoid (HBI) lipids in stored sediments and sediment extracts". Organic Geochemistry. 97: 74–77. doi:10.1016/j.orggeochem.2016.04.010. hdl:10026.1/6762.
- Stein, Ruediger; Fahl, Kirsten; Schreck, Michael; Knorr, Gregor; Niessen, Frank; Forwick, Matthias; Gebhardt, Catalina; Jensen, Laura; Kaminski, Michael; Kopf, Achim; Matthiessen, Jens (2016-04-04). "Evidence for ice-free summers in the late Miocene central Arctic Ocean". Nature Communications. 7 (1): 11148. Bibcode:2016NatCo...711148S. doi:10.1038/ncomms11148. ISSN 2041-1723. PMC 4822014. PMID 27041737.
- Belt, Simon T. (2018-11-01). "Source-specific biomarkers as proxies for Arctic and Antarctic sea ice". Organic Geochemistry. 125: 277–298. doi:10.1016/j.orggeochem.2018.10.002. hdl:10026.1/12842. S2CID 133860206.
- Belt, Simon T.; Massé, Guillaume; Vare, Lindsay L.; Rowland, Steven J.; Poulin, Michel; Sicre, Marie-Alexandrine; Sampei, Makoto; Fortier, Louis (December 2008). "Distinctive 13C isotopic signature distinguishes a novel sea ice biomarker in Arctic sediments and sediment traps". Marine Chemistry. 112 (3–4): 158–167. doi:10.1016/j.marchem.2008.09.002.
- Brown, Thomas (2011). "Production and preservation of the Arctic sea ice diatom biomarker IP25".
{{cite journal}}: Cite journal requires|journal=(help) - Massé, Guillaume; Belt, Simon T.; Crosta, Xavier; Schmidt, Sabine; Snape, Ian; Thomas, David N.; Rowland, Steven J. (October 2011). "Highly branched isoprenoids as proxies for variable sea ice conditions in the Southern Ocean". Antarctic Science. 23 (5): 487–498. Bibcode:2011AntSc..23..487M. doi:10.1017/S0954102011000381. ISSN 0954-1020. S2CID 130625079.
- Schmidt, Katrin; Brown, Thomas A.; Belt, Simon T.; Ireland, Louise C.; Taylor, Kyle W. R.; Thorpe, Sally E.; Ward, Peter; Atkinson, Angus (2018-04-04). "Do pelagic grazers benefit from sea ice? Insights from the Antarctic sea ice proxy IPSO<sub>25</sub>". Biogeosciences. 15 (7): 1987–2006. Bibcode:2018BGeo...15.1987S. doi:10.5194/bg-15-1987-2018. ISSN 1726-4189.
- Belt, Simon T. (January 2019). "What do IP25 and related biomarkers really reveal about sea ice change?". Quaternary Science Reviews. 204: 216–219. Bibcode:2019QSRv..204..216B. doi:10.1016/j.quascirev.2018.11.025. hdl:10026.1/13151. S2CID 134185260.
- Méheust, Marie; Stein, Ruediger; Fahl, Kirsten; Max, Lars; Riethdorf, Jan-Rainer (April 2016). "High-resolution IP25-based reconstruction of sea-ice variability in the western North Pacific and Bering Sea during the past 18,000 years". Geo-Marine Letters. 36 (2): 101–111. Bibcode:2016GML....36..101M. doi:10.1007/s00367-015-0432-4. ISSN 0276-0460. S2CID 131249679.