Holocentric chromosome
Holocentric chromosomes are chromosomes that possess multiple kinetochores along their length rather than the single centromere typical of other chromosomes.[1] They were first described in cytogenetic experiments in 1935. Since this first observation, the term holocentric chromosome has referred to chromosomes that: i) lack the primary constriction corresponding to the centromere observed in monocentric chromosomes;[2] and ii) possess multiple kinetochores dispersed along the entire chromosomal axis, such that microtubules bind to the chromosome along its entire length and move broadside to the pole from the metaphase plate.[3] Holocentric chromosomes are also termed holokinetic, because, during cell division, the sister chromatids move apart in parallel and do not form the classical V-shaped figures typical of monocentric chromosomes.[4][5][6]
Holocentric chromosomes have evolved several times during both animal and plant evolution, and are currently reported in about eight hundred diverse species, including plants, insects, arachnids, and nematodes.[7][8] As a consequence of their diffuse kinetochores, holocentric chromosomes may stabilize chromosomal fragments created by accidental double-strand breaks, preventing loss of the fragments and favouring karyotype rearrangements.[9][10] However, holocentric chromosomes may also present limitations to crossing over, causing a restriction of the number of chiasma in bivalents,[11] and may cause a restructuring of meiotic divisions resulting in an "inverted" meiosis.[12]
Evolution
Holocentric chromosomes were described for the first time by Franz Schrader in 1935, who identified chromosomes with a diffuse kinetochore (or with a diffuse kinetochore activity), making these chromosomes able to bind to microtubules along their entire length rather than at a single primary constriction, as is typical of chromosomes with centromeres. In recent decades, several studies determined that this behaviour during mitosis can be observed in holocentric/holokinetic chromosomes but also in polykinetic chromosomes, which contain numerous (but discrete) microtubule-binding sites; even so, the term “holocentric/holokinetic” is still commonly used to refer to both processes.[1][5][7]
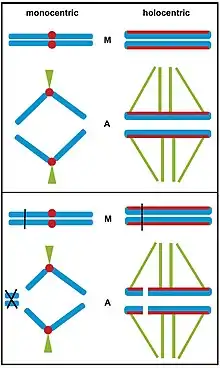
Bottom box: If a chromosomal breakage occurs in a monocentric chromosome, acentric chromosome fragments cannot be attached to microtubules during metaphase (M) and they are subsequently lost during anaphase (A). On the contrary, chromosomal breakage of a holocentric chromosome results in chromosomal fragments that retain kinetic activity due to the chromosome-wide centromere extension and can be properly inherited.
Before molecular methods became available, the presence of holocentric chromosomes was evaluated mostly using cytology and, considering that many species are difficult to study cytologially, it can be surmised that the true presence of holocentrism may be underestimated. In addition, there are several taxa whose chromosomes are still uncharacterized, though their phylogenetic position suggests that they should have holocentric chromosomes.[7][13] The presence of holocentric chromosomes has been up till now assessed in about 800 species, including insects, plants, arachnids, and nematodes,[1][5][7] suggesting that, in general, holocentric chromosomes probably originated by convergent evolution from ancestors possessing monocentric chromosomes. Interesting exceptions are represented by insects belonging to the Oligoneoptera and Neoptera, whose monocentric chromosomes probably evolved from an holocentric ancestor in two different and independent events.[7] Evidence of convergent evolution suggests that holocentrism is adaptive, but the specific conditions under which holocentrism provides a selective advantage seem to be diverse for different taxa.[7][14] Indeed, in phytophagous insects (such as aphids and lepidopterans), holocentrism could be an evolved defense against the production by plants of compounds able to induce chromosomal breakages (clastogens), whereas in other cases, holocentrism may constitute a defense against DNA damage resulting from desiccation and/or other chromosome-breaking factors.[14]
Despite these differences, holocentric chromosomes present intrinsic benefits since chromosomal mutations, such as fissions and fusions, are potentially neutral in holocentric chromosomes with respect to monocentric ones. However, the hypothesis of holocentrism as an anticlastogenic adaptation needs more systematic testing, including both controlled laboratory experiments and field studies across clastogenic gradients and large-scale phylogenetic analyses.[8] At the same time, Nagaki et al.[15] proposed that holocentrism can be easily acquired during plant and animal evolution by a slight difference in the kinetochore origin. In particular, they hypothesized that if the direction of kinetochore origin turns by 90° and occurs along the chromosome axes up to the telomeric regions, it is possible to “generate” holocentric chromosomes without any further step.
Structure
A detailed molecular analysis of the structure of holocentric chromosomes is currently available only for the nematode Caenorhabditis elegans,[16][17] whereas the presence of true holokinetic nature has also been confirmed in other taxa by the evidence that experimentally induced chromosome fragments continue to attach to the spindle and segregate correctly.[3] For most of the species, data about holocentrism are related to the analysis of the behaviour of chromosomes during anaphase migration since holocentric sister chromatids migrate in parallel to the spindle poles, in contrast to monocentric ones in which pulling forces are exerted on a single chromosomal point and chromosome arms trail behind. As a consequence, chromatids of holocentric chromosomes move apart in parallel and do not form the classical V-shaped figures typical of monocentric ones.[4] Moreover, if a holocentric chromosome is fragmented (for instance by X-ray irradiation), each fragment retains centromere activity and can segregate properly to the poles.
In different organisms
Arthropods
Among arthropods, the presence of holocentric chromosome has been reported in different species belonging to insects (Odonata, Zoraptera, Dermaptera, Psocoptera, Phthiraptera, Thysanoptera, Hemiptera, Trichoptera and Lepidoptera), scorpions (Buthoidea), mites and ticks of the superorder Acariformes and genus Rhipicephalus (Ixodidae), spiders (Dysderidae and Segestridae),[7][14] millipedes[18] and centipedes.[18] Despite this widespread occurrence, most of the currently available data on holocentrism is related to aphid and lepidopteran species.[7][5] In aphids, holocentric chromosomes have been deeply studied and their ability to stabilize chromosomal fragments has been associated to their phytophagous life style. Indeed, several plants produce chemicals able to induce DNA damage to pest insects. Nicotine, for instance, is a naturally occurring alkaloid found primarily in members of the solanaceous plant family (including Nicotiana tabacum) that can cause replication fork stress resulting in various forms of DNA damage, including chromosomal fragmentations.[19][20] Similar effects have been also reported by other plant-produced molecules, such as caffeine and ethanol.[19][20] In view of their ability to favour the inheritance of chromosomal fragments, holocentrism has been associated to recurrent changes in the karyotypes of some aphid species and in particular in the peach potato aphid Myzus persicae, where both inter- and intra-individual rearranged karyotypes have been also observed.[21][22] Aphids also possess a constitutive expression of the telomerase coding gene so that they can initiate a de novo synthesis of telomere sequences at internal breakpoints, resulting in the stabilization of chromosomal fragments.[23][24] Among non-polyploid animals, Lepidoptera exhibit the highest variance in chromosome number between species within a genus and notable levels of interspecific and intraspecific karyotype variability.[12][25][26] Lepidoptera indeed tolerate chromosomal variations in view of their holokinetic chromosomes, which facilitate the successful inheritance of novel fission or fusion fragments. As a consequence, Lepidoptera can avoid the deleterious consequences of large-scale chromosomal fission and fusion.[12][25][26] Nevertheless, they can sometimes tolerate heterozygosity for multiple rearrangements in hybrids between population with differences in their karyotype, raising questions about additional mechanisms that rescue fertility in chromosomal hybrids. In Lepidoptera, therefore, chromosome evolution is believed to play a role in reinforcing speciation.[12] Comparing the genomes of lepidopteran species it has been also possible to analyse the effect of holocentrism in terms of rate of fixed chromosomal rearrangements. This approach evidenced in Lepidoptera two chromosome breaks per megabase of DNA per Million of years: a rate that is much higher than what observed in Drosophila and it is a direct consequence of the holocentric nature of the lepidopteran genomes.[27][28] At a structural level, insect holocentric chromosomes have not been studied in details, but it is interesting to underline the absence of homologues of CENP-C and CENP-A, previously considered essential for kinetochore functioning in eukaryotes.[29]
Nematodes
The best known group of holocentric species can be found in the Secernentea class of the nematodes, which includes C. elegans.[16][17] other nematodes are usually described as holocentric because of their phylogenetic relationship to C. elegans, but real karyotypic evidences are scarce or controversial.[30][31][32] Nematode development is typically characterized by fixed lineages and a single inappropriate cell death, therefore, it has been suggested that holocentrism could avoid the disastrous consequences of unrepaired chromosome breakage events.[33] The availability of several molecular and genomic resources allowed a detailed characterization of C. elegans holocentric chromosomes and in particular the structure of the kinetochore has been molecularly dissected.[34][35] Current data suggest that C. elegans kinetochores form paired lines or plates on opposite faces of condensed mitotic chromosomes,[35] where each line represents the diffuse kinetochore of a single chromatid. transmission electron microscopy of C. elegans chromosomes revealed that the kinetochore has a trilaminar structure very similar to that observed in monocentric chromosomes[35][36] More than 30 different proteins have been identified as components of the C. elegans kinetochore and half of them was already known as functioning in the kinetochores of monocentric chromosomes. Among these, highly studied proteins include homologues of CENP-C and CENP-A, which are highly conserved structural component of the kinetochore in eukaryotes.[36][37] Contrarily to what generally observed in monocentric chromosomes, in holocentric ones the preferential localization of centromeres within heterochromatic areas is missing together with the presence of specific DNA sequences that in C. elegans are not required for the assembly of a functional kinetochore.[36][37] In this regard, it has been observed that holocentric chromosomes of nematodes are unique because they have a large number of satellites scattered throughout their genome, whereas no scattered satellites are found in the monocentric chromosomes of the nematode trichinella spiralis.[38] Interestingly, these satellite DNAs are not conserved in their sequences among species suggesting that highly repetitive DNAs may facilitate the formation of kinetochores in view of their repetitiveness rather than for their specific sequence.[38] The absence of a localized centromere prompted several studies to identify proteins that are involved in the sister chromatid cohesion assessing that it is accomplished by a separate complex of conserved proteins, termed cohesin, that is composed of the core subunits Scc3, Smc1, Smc3 and Scc1. Interestingly, they play the same function in organisms with monocentric chromosomes with an exception related to the subunit Scc1, whose gene in addition to the Scc1 orthologue present three additional paralogous genes.[36][37]
Plants
In plants, holocentric chromosomes have been found in zygnematophycean algae,[39] in the genera Myristica (Myristicaceae), Chionographis (Melanthiaceae), Cuscuta (Convolvulaceae) and Droseraceae,[40][41][42] in the species Trithuria submersa (Hydatellaceae), Prionium serratum (Thurniaceae),[43][44] and, among higher plants, in many genera belonging to the families Cyperaceae and Juncaceae, including the snowy woodrush Luzula nivea (Juncaceae), the most well-studied holocentric plant.[45][46] In Luzula spp., the centromeric activity is localized simultaneously at several evenly spaced sites along each chromosome and chromosomes can be fragmented naturally or by irradiation into smaller (but viable) chromosomes.[47][48] The presence of rearranged karyotypes does not affect fitness, as assessed by studies reporting that Luzula hybrids with parents possessing smaller and larger chromosomes showed the smaller chromosomes aligned and paired with the larger ones.[47] Similarly, in plants belonging to the genus Carex, differentiation of the karyotype has been demonstrated to correlate with genetic divergence within species,[49] among populations within species,[50] and within populations,[51] suggesting that, as previously reported in the Lepidoptera,[12] holocentric chromosome rearrangements contribute to genetic differentiation at different evolutionary scales in Carex evolution and speciation. In plants it has also been suggested that the diffuse kinetochore of holocentric chromosomes may suppress the meiotic drive of centromeric repeats and its negative consequences.[46] In particular, the expansions (or contractions) of centromeric repeats may lead to a larger (or smaller) kinetochore, which attracts more (or fewer) microtubules during meiosis.[17][46] This hypothesis, which correlates the presence of holocentric chromosomes with centromere drive suppression, is very intriguing but it only explains the evolution of chromosomal holocentrism in meiosis and not in mitosis, a fact which is not trivial considering that some species with holocentric chromosomes may present a restriction of kinetochore activity during meiosis.[17][52] Similarly to what was previously reported for C. elegans, in L. elegans centromeres are not made by centromere-associated retrotransposons nor centromere-associated satellite DNAs, but cenH3 proteins seem to be associated with a centromere-specific chromatin folding rather than with specific centromeric DNA sequences.[53] Conservation of elements between mono- and holocentric chromosomes is not limited to centromeric proteins, but it is also extended to epigenetic marks. Indeed, the cell cycle-dependent phosphorylation of serine 10 or serine 28 of H3 (that is typically enriched in pericentric regions of monocentric plant chromosomes) occurs uniformly along the Luzula chromosomes.[53] As previously described in aphids, L. elegans possesses a rapid and efficient de novo telomere formation based on a telomerase-mediated healing process that is active immediately after chromosomal damage by irradiation of chromosomes.[54] Newly formed telomere repeats were cytologially detectable 21 days after irradiation in about 50% of cases, with a complete healing of telomeres after three months favouring the fragment stabilization and karyotype fixation.[54]
Meiosis
In the late 19th century, van Beneden (1883) and Boveri (1890) described meiosis for the first time through a careful observation of germ cell formation in the nematode Ascaris. These observations, together with several further analyses, evidenced that canonical meiosis consists of a first division (called reductional division) that involves the segregation of chromosomal homologs resulting in the reduction of chromosome number, followed by a second division (called equational division) that involves the segregation of sister chromatids. A general rule for meiosis is therefore: first homologs, then sisters.
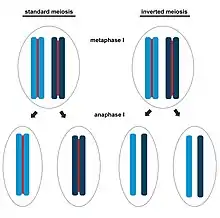
However, the understanding of the reductional division in meiosis of Ascaris spp. has been obtained by studying the holocentric chromosomes which, in many other taxa, follow a reverse order of meiotic division.[12] Indeed, as reported in several nematodes, in insects belonging to the Hemiptera and Lepidoptera,[55][56] in mites,[57] and in some flowering plants,[8] species with holocentric chromosomes generally present an inverted meiotic sequence, in which segregation of homologs is postponed until the second meiotic division.
Furthermore, in most cases of inverted meiosis the absence of a canonical kinetochore structure has been observed, together with a restriction of the kinetic activity to the chromosomal ends.[12][55][56] These changes are related to the peculiar cohesion occurring in tetrads of the holocentric homologous chromosomes during meiosis that impose obstacles to the releases of chromosomes involved in multiple crossing over events.[55][56][57] In the holocentric chromosomes of C. elegans female meiosis,[58] this problem is circumvented by restricting crossing over to form only a single chiasma per bivalent and triggering the redistribution of kinetochore proteins along the bivalent axis, forming meiosis-specific cup-like structures that uniformly coat each half-bivalent but are excluded from the mid-bivalent region.[58] During anaphase I, C. elegans homologous chromosomes are segregated to the poles by microtubule-pushing from the mid-bivalent regions towards the poles.[58]
In contrast to C. elegans, other organisms with holocentric chromosomes, including both plants and insects,[12][55][56] circumvent this problem by segregating sister chromatids during meiosis I, leading to the term inverted meiosis, in which the order of reductional and equational division is inverted with respect to canonical meiosis. In this case therefore the separation of homologous chromosomes follows rather than precedes the segregation of sister chromatids. However, in order to have a successful inverted meiosis, it is necessary that a bipolar orientation of sister kinetochores occurs, together with their attachment to microtubules from opposite spindle poles in meiosis I. This allows the segregation of sister chromatids to opposite poles in anaphase I (equational division), but requires a mechanism to align and pair homologous chromosomes during the second meiotic division.[55][56][57] Interestingly, the presence of inverted meiosis can also facilitate proper chromosome segregation in hybrids from parental species with differences in their karyotypes or derived by populations with rearranged karyotypes, allowing rescue of the fertility and viability of hybrids and promoting a fast karyotype evolution and possibly chromosomal speciation, as reported in the Lepidoptera.[12]
References
- Schrader F (1935). "Notes an the Mitotic Behavior of Long Chromosomes". Cytologia. 6 (4): 422–430. doi:10.1508/cytologia.6.422. ISSN 0011-4545.
- Mandrioli M, Manicardi GC (2003). "Analysis of insect holocentric chromosomes by atomic force microscopy". Hereditas. 138 (2): 129–32. doi:10.1034/j.1601-5223.2003.01661.x. hdl:11380/5054. PMID 12921164.
- Hughes-Schrader S; Schrader F (January 1961). "The kinetochore of the Hemiptera". Chromosoma. 12 (1): 327–50. doi:10.1007/bf00328928. PMID 13716663. S2CID 5169204.
- Wrensch DL, Kethley JB, Norton RA (1994). "Cytogenetics of Holokinetic Chromosomes and Inverted Meiosis: Keys to the Evolutionary Success of Mites, with Generalizations on Eukaryotes". Mites. Springer US. pp. 282–343. doi:10.1007/978-1-4615-2389-5_11. ISBN 978-1-4613-6012-4.
- White MJ (1973). Animal cytology and evolution (3d ed.). Cambridge [England]: University Press. ISBN 0-521-07071-6. OCLC 674359.
- Mandrioli M, Manicardi GC (August 2012). "Unlocking holocentric chromosomes: new perspectives from comparative and functional genomics?". Current Genomics. 13 (5): 343–9. doi:10.2174/138920212801619250. PMC 3401891. PMID 23372420.
- Melters DP, Paliulis LV, Korf IF, Chan SW (July 2012). "Holocentric chromosomes: convergent evolution, meiotic adaptations, and genomic analysis". Chromosome Research. 20 (5): 579–93. doi:10.1007/s10577-012-9292-1. PMID 22766638. S2CID 3351527.
- Benavente R (May 1982). "Holocentric chromosomes of arachnids: Presence of kinetochore plates during meiotic divisions". Genetica. 59 (1): 23–27. doi:10.1007/bf00130811. ISSN 0016-6707. S2CID 21290751.
- Monti V, Lombardo G, Loxdale HD, Manicardi GC, Mandrioli M (March 2012). "Continuous occurrence of intra-individual chromosome rearrangements in the peach potato aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae)". Genetica. 140 (1–3): 93–103. doi:10.1007/s10709-012-9661-x. hdl:11380/739277. PMID 22644285. S2CID 15715405.
- Manicardi GC, Nardelli A, Mandrioli M (2015-08-01). "Fast chromosomal evolution and karyotype instability: recurrent chromosomal rearrangements in the peach potato aphidMyzus persicae(Hemiptera: Aphididae)". Biological Journal of the Linnean Society. 116 (3): 519–529. doi:10.1111/bij.12621. ISSN 0024-4066.
- Nokkala S, Kuznetsova VG, Maryanska-Nadachowska A, Nokkala C (2004). "Holocentric chromosomes in meiosis. I. Restriction of the number of chiasmata in bivalents". Chromosome Research. 12 (7): 733–9. doi:10.1023/b:chro.0000045797.74375.70. PMID 15505408. S2CID 22226624.
- Lukhtanov VA, Dincă V, Friberg M, Šíchová J, Olofsson M, Vila R, et al. (October 2018). "Versatility of multivalent orientation, inverted meiosis, and rescued fitness in holocentric chromosomal hybrids". Proceedings of the National Academy of Sciences of the United States of America. 115 (41): E9610–E9619. doi:10.1073/pnas.1802610115. PMC 6187165. PMID 30266792.
- Bureš P, Zedek F, Marková M (2012-08-23). "Holocentric Chromosomes". Plant Genome Diversity. Vol. 2. Springer Vienna. pp. 187–208. doi:10.1007/978-3-7091-1160-4_12. ISBN 978-3-7091-1159-8.
- Escudero M, Márquez-Corro JI, Hipp AL (2016-09-01). "The Phylogenetic Origins and Evolutionary History of Holocentric Chromosomes". Systematic Botany. 41 (3): 580–585. doi:10.1600/036364416x692442. ISSN 0363-6445. S2CID 89161837.
- Nagaki K, Kashihara K, Murata M (July 2005). "Visualization of diffuse centromeres with centromere-specific histone H3 in the holocentric plant Luzula nivea". The Plant Cell. 17 (7): 1886–93. doi:10.1105/tpc.105.032961. PMC 1167539. PMID 15937225.
- Howe M, McDonald KL, Albertson DG, Meyer BJ (June 2001). "HIM-10 is required for kinetochore structure and function on Caenorhabditis elegans holocentric chromosomes". The Journal of Cell Biology. 153 (6): 1227–38. doi:10.1083/jcb.153.6.1227. PMC 2192032. PMID 11402066.
- Zedek F, Bureš P (2012-01-23). "Evidence for centromere drive in the holocentric chromosomes of Caenorhabditis". PLOS ONE. 7 (1): e30496. Bibcode:2012PLoSO...730496Z. doi:10.1371/journal.pone.0030496. PMC 3264583. PMID 22291967.
- Ogawa K (1953). "Chromosome Studies in the Myriapoda". Japanese Journal of Genetics (遺伝學雑誌). 28 (1): 12–18. doi:10.1266/jjg.28.12.
- Richards RI (June 2001). "Fragile and unstable chromosomes in cancer: causes and consequences". Trends in Genetics. 17 (6): 339–45. doi:10.1016/s0168-9525(01)02303-4. PMID 11377796.
- Freudenreich CH (September 2007). "Chromosome fragility: molecular mechanisms and cellular consequences". Frontiers in Bioscience. 12 (12): 4911–24. doi:10.2741/2437. PMID 17569619.
- Manicardi GC, Mandrioli M, Blackman RL (February 2015). "The cytogenetic architecture of the aphid genome". Biological Reviews of the Cambridge Philosophical Society. 90 (1): 112–25. doi:10.1111/brv.12096. hdl:11380/1062551. PMID 24593177. S2CID 21793030.
- Monti V, Mandrioli M, Rivi M, Manicardi GC (2011-12-16). "The vanishing clone: karyotypic evidence for extensive intraclonal genetic variation in the peach potato aphid, Myzus persicae (Hemiptera: Aphididae)". Biological Journal of the Linnean Society. 105 (2): 350–358. doi:10.1111/j.1095-8312.2011.01812.x. ISSN 0024-4066.
- Monti V, Giusti M, Bizzaro D, Manicardi GC, Mandrioli M (July 2011). "Presence of a functional (TTAGG)(n) telomere-telomerase system in aphids". Chromosome Research. 19 (5): 625–33. doi:10.1007/s10577-011-9222-7. hdl:11380/659235. PMID 21667174. S2CID 25127536.
- Mandrioli M, Borsatti F (2007-11-17). "Analysis of heterochromatic epigenetic markers in the holocentric chromosomes of the aphid Acyrthosiphon pisum". Chromosome Research. 15 (8): 1015–22. doi:10.1007/s10577-007-1176-4. hdl:11380/594885. PMID 18004669. S2CID 20329150.
- Marec F, Tothova A, Sahara K, Traut W (December 2001). "Meiotic pairing of sex chromosome fragments and its relation to atypical transmission of a sex-linked marker in Ephestia kuehniella (Insecta: Lepidoptera)". Heredity. 87 (Pt 6): 659–71. doi:10.1046/j.1365-2540.2001.00958.x. PMID 11903561. S2CID 415409.
- Hill J, Rastas P, Hornett EA, Neethiraj R, Clark N, Morehouse N, et al. (June 2019). "Unprecedented reorganization of holocentric chromosomes provides insights into the enigma of lepidopteran chromosome evolution". Science Advances. 5 (6): eaau3648. Bibcode:2019SciA....5.3648H. doi:10.1126/sciadv.aau3648. PMC 6561736. PMID 31206013.
- d'Alençon E, Sezutsu H, Legeai F, Permal E, Bernard-Samain S, Gimenez S, et al. (April 2010). "Extensive synteny conservation of holocentric chromosomes in Lepidoptera despite high rates of local genome rearrangements". Proceedings of the National Academy of Sciences of the United States of America. 107 (17): 7680–5. Bibcode:2010PNAS..107.7680D. doi:10.1073/pnas.0910413107. PMC 2867904. PMID 20388903.
- Lukhtanov VA, Dincă V, Talavera G, Vila R (April 2011). "Unprecedented within-species chromosome number cline in the Wood White butterfly Leptidea sinapis and its significance for karyotype evolution and speciation". BMC Evolutionary Biology. 11 (1): 109. doi:10.1186/1471-2148-11-109. PMC 3113740. PMID 21507222.
- Drinnenberg IA, deYoung D, Henikoff S, Malik HS (2014-08-18). "Author response: Recurrent loss of CenH3 is associated with independent transitions to holocentricity in insects". eLife. doi:10.7554/elife.03676.020.
- Mutafova T, Dimitrova Y, Komandarev S (1982). "The karyotype of four Trichinella species". Zeitschrift für Parasitenkunde. 67 (1): 115–20. doi:10.1007/bf00929519. PMID 7072318. S2CID 22178390.
- Spakulová M, Králová I, Cutillas C (March 1994). "Studies on the karyotype and gametogenesis in Trichuris muris". Journal of Helminthology. 68 (1): 67–72. doi:10.1017/s0022149x0001350x. PMID 8006389. S2CID 35922875.
- Post R (November 2005). "The chromosomes of the Filariae". Filaria Journal. 4 (1): 10. doi:10.1186/1475-2883-4-10. PMC 1282586. PMID 16266430.
- Pimpinelli S, Goday C (September 1989). "Unusual kinetochores and chromatin diminution in Parascaris". Trends in Genetics. 5 (9): 310–5. doi:10.1016/0168-9525(89)90114-5. PMID 2686123.
- Dernburg AF (June 2001). "Here, there, and everywhere: kinetochore function on holocentric chromosomes". The Journal of Cell Biology. 153 (6): F33-8. doi:10.1083/jcb.153.6.f33. PMC 2192025. PMID 11402076.
- Maddox PS, Oegema K, Desai A, Cheeseman IM (2004). ""Holo"er than thou: chromosome segregation and kinetochore function in C. elegans". Chromosome Research. 12 (6): 641–53. doi:10.1023/b:chro.0000036588.42225.2f. PMID 15289669. S2CID 38989489.
- Albertson DG, Thomson JN (October 1982). "The kinetochores of Caenorhabditis elegans". Chromosoma. 86 (3): 409–28. doi:10.1007/bf00292267. PMID 7172865. S2CID 24941882.
- Monen J, Maddox PS, Hyndman F, Oegema K, Desai A (December 2005). "Differential role of CENP-A in the segregation of holocentric C. elegans chromosomes during meiosis and mitosis". Nature Cell Biology. 7 (12): 1248–55. doi:10.1038/ncb1331. PMID 16273096. S2CID 37436777.
- Subirana JA, Messeguer X (2013-04-24). "A satellite explosion in the genome of holocentric nematodes". PLOS ONE. 8 (4): e62221. Bibcode:2013PLoSO...862221S. doi:10.1371/journal.pone.0062221. PMC 3634726. PMID 23638010.
- Godward MB (1966). Chromosomes of the Algae. Edward Arnold. pp. 24–51. ISBN 9780713120585.
- Kolodin P, Cempírková H, Bureš P, Horová L, Veleba A, Francová J, Adamec L, Zedek F (December 2018). "Holocentric chromosomes may be an apomorphy of Droseraceae". Plant Systematics and Evolution. 304 (10): 1289–96. doi:10.1007/s00606-018-1546-8. S2CID 52882155.
- Luceño M, Vanzela AL, Guerra M (1998-03-01). "Cytotaxonomic studies in Brazilian Rhynchospora (Cyperaceae), a genus exhibiting holocentric chromosomes". Canadian Journal of Botany. 76 (3): 440–449. doi:10.1139/b98-013. ISSN 0008-4026.
- Kuta E, Bohanec B, Dubas E, Vizintin L, Przywara L (April 2004). "Chromosome and nuclear DNA study on Luzula - a genus with holokinetic chromosomes". Genome. 47 (2): 246–56. doi:10.1139/g03-121. PMID 15060577.
- Kynast RG, Joseph JA, Pellicer J, Ramsay MM, Rudall PJ (September 2014). "Chromosome behavior at the base of the angiosperm radiation: karyology of Trithuria submersa (Hydatellaceae, Nymphaeales)". American Journal of Botany. 101 (9): 1447–55. doi:10.3732/ajb.1400050. PMID 25253705.
- Zedek F, Veselý P, Horová L, Bureš P (June 2016). "Flow cytometry may allow microscope-independent detection of holocentric chromosomes in plants". Scientific Reports. 6 (1): 27161. Bibcode:2016NatSR...627161Z. doi:10.1038/srep27161. PMC 4891681. PMID 27255216.
- Haizel T, Lim YK, Leitch AR, Moore G (2005). "Molecular analysis of holocentric centromeres of Luzula species". Cytogenetic and Genome Research. 109 (1–3): 134–43. doi:10.1159/000082392. PMID 15753569. S2CID 45200884.
- Zedek F, Bureš P (December 2016). "Absence of positive selection on CenH3 in Luzula suggests that holokinetic chromosomes may suppress centromere drive". Annals of Botany. 118 (7): 1347–1352. doi:10.1093/aob/mcw186. PMC 5155603. PMID 27616209.
- Madej A, Kuta E (2001). "Holokinetic chromosomes of Luzula luzuloides [Juncaceae] in callus culture". Acta Biologica Cracoviensia. Series Botanica (43). ISSN 0001-5296.
- Nordenskiöld, Hedda (1961). "Tetrad Analysis and the Course of Meiosis in Three Hybrids of Luzula Campestris". Hereditas. 47 (2): 203–238. doi:10.1111/j.1601-5223.1961.tb01771.x. ISSN 1601-5223.
- Escudero M, Vargas P, Arens P, Ouborg NJ, Luceño M (January 2010). "The east-west-north colonization history of the Mediterranean and Europe by the coastal plant Carex extensa (Cyperaceae)". Molecular Ecology. 19 (2): 352–70. doi:10.1111/j.1365-294X.2009.04449.x. PMID 20002603. S2CID 3577580.
- Hipp AL, Rothrock PE, Roalson EH (2008-12-05). "The Evolution of Chromosome Arrangements in Carex (Cyperaceae)". The Botanical Review. 75 (1): 96–109. doi:10.1007/s12229-008-9022-8. ISSN 0006-8101. S2CID 4489708.
- Escudero M, Weber JA, Hipp AL (August 2013). "Species coherence in the face of karyotype diversification in holocentric organisms: the case of a cytogenetically variable sedge (Carex scoparia, Cyperaceae)". Annals of Botany. 112 (3): 515–26. doi:10.1093/aob/mct119. PMC 3718211. PMID 23723260.
- Wanner G, Schroeder-Reiter E, Ma W, Houben A, Schubert V (December 2015). "The ultrastructure of mono- and holocentric plant centromeres: an immunological investigation by structured illumination microscopy and scanning electron microscopy". Chromosoma. 124 (4): 503–17. doi:10.1007/s00412-015-0521-1. PMID 26048589. S2CID 14145859.
- Cuacos M, H Franklin FC, Heckmann S (2015-10-26). "Atypical centromeres in plants-what they can tell us". Frontiers in Plant Science. 6: 913. doi:10.3389/fpls.2015.00913. PMC 4620154. PMID 26579160.
- Jankowska M, Fuchs J, Klocke E, Fojtová M, Polanská P, Fajkus J, et al. (December 2015). "Holokinetic centromeres and efficient telomere healing enable rapid karyotype evolution". Chromosoma. 124 (4): 519–28. doi:10.1007/s00412-015-0524-y. PMID 26062516. S2CID 2530401.
- Bogdanov YF (May 2016). "[Inverted meiosis and its place in the evolution of sexual reproduction pathways]". Genetika. 52 (5): 541–60. doi:10.1134/s1022795416050033. PMID 29368476. S2CID 18427008.
- Viera A, Page J, Rufas JS (2009). "Inverted meiosis: the true bugs as a model to study". Genome Dynamics. Karger. 5: 137–156. doi:10.1159/000166639. ISBN 978-3-8055-8967-3. PMID 18948713.
- Vanzela AL, Cuadrado A, Guerra M (2003). "Localization of 45S rDNA and telomeric sites on holocentric chromosomes of Rhynchospora tenuis Link (Cyperaceae)". Genetics and Molecular Biology. 26 (2): 199–201. doi:10.1590/S1415-47572003000200014.
- Martinez-Perez E, Schvarzstein M, Barroso C, Lightfoot J, Dernburg AF, Villeneuve AM (October 2008). "Crossovers trigger a remodeling of meiotic chromosome axis composition that is linked to two-step loss of sister chromatid cohesion". Genes & Development. 22 (20): 2886–901. doi:10.1101/gad.1694108. PMC 2569886. PMID 18923085.
![]() This article was adapted from the following source under a CC BY 4.0 license (2020) (reviewer reports):
Mauro Mandrioli; Gian Carlo Manicardi (2020). "Holocentric chromosomes". PLOS Genetics. 16 (7): e1008918. doi:10.1371/JOURNAL.PGEN.1008918. ISSN 1553-7390. PMC 7392213. PMID 32730246. Wikidata Q97932961.
This article was adapted from the following source under a CC BY 4.0 license (2020) (reviewer reports):
Mauro Mandrioli; Gian Carlo Manicardi (2020). "Holocentric chromosomes". PLOS Genetics. 16 (7): e1008918. doi:10.1371/JOURNAL.PGEN.1008918. ISSN 1553-7390. PMC 7392213. PMID 32730246. Wikidata Q97932961.