Hybodontiformes
Hybodontiformes, commonly called hybodonts, are an extinct group of shark-like chondrichthyans, which existed from the late Devonian to the Late Cretaceous. They form the group of Elasmobranchii closest to neoselachians, the clade of modern sharks and rays. Hybodonts were named and are distinguished based on their conical tooth shape. They are also noted for the presence of a spine on each of their two dorsal fins. They were abundant in marine and freshwater environments during the late Paleozoic and early Mesozoic, but were rare in open marine environments by the end of the Jurassic, having been largely replaced by modern sharks, though they were still common in freshwater and marginal marine habitats. They survived until the end of the Cretaceous, before going extinct.
| Hybodontiformes Temporal range: Late Devonian to Late Cretaceous | |
|---|---|
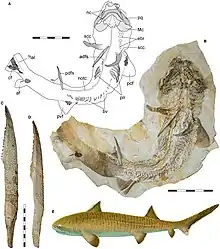 | |
| Fossil and life restoration of Asteracanthus, from the Late Jurassic of Europe | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Eugnathostomata |
| Class: | Chondrichthyes |
| Subclass: | Elasmobranchii |
| Order: | †Hybodontiformes Patterson, 1966 |
| Families | |
| |
Etymology
The term hybodont comes from the Greek word ὕβος or ὑβός meaning hump or hump-backed and ὀδούς, ὀδοντ meaning tooth. This name was given based on their conical compressed teeth.
Taxonomic history
Hybodonts were first described in the nineteenth century based on isolated fossil teeth (Agassiz, 1837). Hybodonts were first separated from living sharks by Zittel (1911).[1] Hybodontiformes are a type of elasmobranch, and the sister group to Neoselachii. Hybodontiformes are classified in the Elasmobranchii with Xenacanthiformes, Ctenacanthiformes and Neoselachii. Hybodonts are divided into a number of families, including Hybodontidae, but the higher level taxonomy of hybodonts, especially Mesozoic taxa, is poorly resolved.[2]
Morphology and teeth
.jpg.webp)
Hybodonts reached a maximum size of 2–3 metres (6.6–9.8 ft) in length, and had a relatively robust bodyform. Due to their cartilaginous skeletons usually disintegrating upon death like other chondrichthyans, hybodonts are generally described and identified based on size and shape of teeth and fin spine fossils, which are more likely to be preserved.[2] Rare complete skeletons are known from areas of exceptional preservation.[3][2][4]
Hybodonts are recognized as having teeth with a prominent cusp which is higher than lateral cusplets.[5] Hybodont teeth are often preserved as incomplete fossils because the base of the tooth is not well attached to the crown.[5] Hybodonts were initially divided into two groups based on their tooth shape.[6] One group had teeth with acuminate cusps that lacked a pulp cavity; these are called osteodont teeth. The other group had a different cusp arrangement and had a pulp cavity, these are called orthodont teeth.[7] For example, the hybodont species Heterophychodus steinmanni have osteodont teeth with vascular canals of dentine which are arranged vertically parallel to each other, also called ‘tubular dentine’.[8] The crowns of these osteodont teeth are covered with a single layer of enameloid. Hybodont teeth served a variety of functions depending on the species, including grinding, crushing (durophagy), tearing, clutching, and even cutting.[2]
Hybodonts are characterized by having two dorsal fins each preceded by a fin spine with a specific shape. The fin spines shape is used to distinguish hybodonts from other shark groups and different hybodont species.[9] The fin spines are elongate and gently curved toward the back of the animal.[9] Male hybodonts had either one or two pairs of cephalic spines on their heads.[10] Hybodonts had a fully heterocercal tail fin, where the upper lobe of the fin was much larger than the lower one due to the spine extending into it.[11]
 Teeth of Durnovariaodus, a member of the family Hybodontidae
Teeth of Durnovariaodus, a member of the family Hybodontidae Teeth of Strophodus a specialised durophagous hybodont belonging to the family Acrodontidae
Teeth of Strophodus a specialised durophagous hybodont belonging to the family Acrodontidae Jaws of the hybodontid Asteracanthus, showing the arrangement of the teeth in jaws
Jaws of the hybodontid Asteracanthus, showing the arrangement of the teeth in jaws.jpg.webp)
Habitat

Hybodont fossils are found in depositional environments ranging from marine to fluvial (river deposits).[12] Many hybodonts are thought to have been euryhaline, able to tolerate a wide range of salinities.[13] Hybodonts inhabited freshwater environments at least since the Permian. Based on isotopic analysis, some species of hybodonts are likely to have permanently lived in freshwater environments,[14][15] while others may have migrated between marine and freshwater environments.[16]
Evolutionary history
The earliest hybodont remains are from the latest Devonian (Famennian) of Iran, belonging to the genus Roongodus,[17] as well as remains assigned to Lissodus of the same age from Belgium.[18] Although the first fossils of hybodonts are from the latest Devonian, they likely branched off from neoselachians (modern sharks) during the early Devonian.[19] Carboniferous hybodonts include both durophagous and non-durophagous forms, while durophagous forms were dominant during the Permian period.[18] By the Permian period, hybodonts had a global distribution.[18][20][21] The Permian-Triassic extinction event only had a limited effect on hybodont diversity.[22] Maximum hybodont diversity is observed during the Triassic. During the Triassic and Early Jurassic, hybodontiforms were the dominant elasmobranchs in both marine and non-marine environments.[12] A shift in hybodonts was seen during the Middle Jurassic, a transition between the distinctly different assemblages seen in the Triassic – Early Jurassic and the Late Jurassic – Cretaceous.[12] As neoselachians (group of modern sharks) diversified further during the Late Jurassic, hybodontiforms became less prevalent in open marine conditions but remained diverse in fluvial and restricted settings during the Cretaceous.[12] By the Cretaceous, hybodontiforms were primarily (though not solely) restricted to freshwater settings.[8] The end-Cretaceous extinction of hybodont sharks may have been caused more by competition with other sharks than by the meteorite impact and volcanic eruptions cited to be the main cause of this extinction event.[23] Most other sharks were not significantly affected by the end-Cretaceous extinction, also suggesting that competition led to the demise of hybodonts.
Ecology
Hybodus is thought to have been an active predator of fast moving prey.[23] Hybodonts have a wide variety of tooth shapes. This variety suggests that they took advantage of multiple food sources.[5] It is thought that some hybodonts which had wider, flatter, teeth specialized in crushing hard-shelled prey (durophagy), evidence for this includes well-developed wear facets on teeth from Lissodus.[12] Species described from Thailand have a range of teeth shapes, suggesting multiple feeding habits. Bulbous teeth were used for crushing hard shelled bottom-dwelling prey.[24] Others were opportunistic feeders and were species that had a diet of large soft-bodied prey.[8] Little is known about the reproductive habits of hybodonts. One study found abundant fossil teeth and eggs sacks in freshwater lake deposits from the Triassic of Kyrgyzstan.[25] The site was interpreted as an ancient shark nursery based on the abundance of eggs and juvenile fossils and the limited number of adult specimens.[25]
One genus of hybodont, Onychoselache of the lower Carboniferous of Scotland, is suggested to have been capable of amphibious locomotion, similar to modern orectolobiform sharks such as bamboo and epaulette sharks, due to its well-developed pectoral fins.[26]
References
- Zittel, K. von, 1911, Grunzuege der Palaontologie, 2 ed. II. Abt. Vertebrata, vii + 598 pp. R. Oldenburg Verlag, Muchen, Berlin.
- Stumpf, Sebastian; López‐Romero, Faviel A.; Kindlimann, René; Lacombat, Frederic; Pohl, Burkhard; Kriwet, Jürgen (August 2021). Cavin, Lionel (ed.). "A unique hybodontiform skeleton provides novel insights into Mesozoic chondrichthyan life". Papers in Palaeontology. 7 (3): 1479–1505. doi:10.1002/spp2.1350. ISSN 2056-2799. S2CID 234204226.
- Lane, J. A., and Maisey, J. G., 2009, Pectoral Anatomy of Tribodus limae (Elasmobranchii: Hybodontiformes) from the Lower Cretaceous of Northeastern Brazil: Journal of Vertebrate Paleontology, v. 29, no. 1, p. 25-38.
- Wang, N.-Z.; Zhang, X.; Zhu, M.; Zhao, W.-J. (May 2009). "A new articulated hybodontoid from Late Permian of northwestern China". Acta Zoologica. 90: 159–170. doi:10.1111/j.1463-6395.2008.00382.x.
- Koot, M. B., Cuny, G., Tintori, A., and Twitchett, R. J., 2013, A new diverse shark fauna from the Wordian (Middle Permian) Khuff Formation in the interior Haushi-Huqf area, Sultanate of Oman: Palaeontology, v. 56, no. 2, p. 303-343.
- Agassiz, L., 1833-1844, Recherches sur les poisons fossils. Neuchatel, 5 vols. 1420 pp. with supplement.
- Maisey, J. G., 1982, The anatomy and interrelationships of Mesozoic hybodont sharks: American Museum Novitates, v. 2724.
- Cuny, G., Suteethorn, V., Buffetaut, E., and Philippe, M., 2003, Hybodont sharks from the Mesozoic Khorat Group of Thailand: Mahasarakham University Journal, v. 22.
- Maisey, J. G., 1978, Growth and form of spines in hybodont sharks: Palaeontology, v. 21, no. 3, p. 657-666.
- Rees, Jan (March 2002). "Shark fauna and depositional environment of the earliest Cretaceous Vitabäck Clays at Eriksdal, southern Sweden". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 93 (1): 59–71. doi:10.1017/S0263593300000328. ISSN 1473-7116. S2CID 130984933.
- Kim, Sun H.; Shimada, Kenshu; Rigsby, Cynthia K. (March 2013). "Anatomy and Evolution of Heterocercal Tail in Lamniform Sharks: CAUDAL FINS OF LAMNIFORM SHARKS". The Anatomical Record. 296 (3): 433–442. doi:10.1002/ar.22647. PMID 23381874. S2CID 205411092.
- Rees, J. A. N., and Underwood, C. J., 2008, Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic): Palaeontology, v. 51, no. 1, p. 117-147.
- Batchelor, Trevor John (December 2013). "A new species of Vectiselachos (Chondrichthyes, Selachii) from the Early Cretaceous of southern England". Proceedings of the Geologists' Association. 124 (6): 967–972. Bibcode:2013PrGA..124..967B. doi:10.1016/j.pgeola.2013.05.001.
- Fischer, Jan; Schneider, Jörg W.; Voigt, Silke; Joachimski, Michael M.; Tichomirowa, Marion; Tütken, Thomas; Götze, Jens; Berner, Ulrich (2013-03-29). "Oxygen and strontium isotopes from fossil shark teeth: Environmental and ecological implications for Late Palaeozoic European basins". Chemical Geology. 342: 44–62. Bibcode:2013ChGeo.342...44F. doi:10.1016/j.chemgeo.2013.01.022. ISSN 0009-2541.
- Klug, Stefanie; Tütken, Thomas; Wings, Oliver; Pfretzschner, Hans-Ulrich; Martin, Thomas (2010-09-01). "A Late Jurassic freshwater shark assemblage (Chondrichthyes, Hybodontiformes) from the southern Junggar Basin, Xinjiang, Northwest China". Palaeobiodiversity and Palaeoenvironments. 90 (3): 241–257. doi:10.1007/s12549-010-0032-2. ISSN 1867-1608. S2CID 129236098.
- Leuzinger, L.; Kocsis, L.; Billon-Bruyat, J.-P.; Spezzaferri, S.; Vennemann, T. (2015-12-07). "Stable isotope study of a new chondrichthyan fauna (Kimmeridgian, Porrentruy, Swiss Jura): an unusual freshwater-influenced isotopic composition for the hybodont shark <i>Asteracanthus</i>". Biogeosciences. 12 (23): 6945–6954. Bibcode:2015BGeo...12.6945L. doi:10.5194/bg-12-6945-2015. ISSN 1726-4189. S2CID 129044300.
- Hairapetian, V. and Ginter, M. 2009. Famennian chondrichthyan remains from the Chahriseh section, central Iran. Acta Geologica Polonica, 59, 173–200.
- Hodnett, J-P., Elliott, D. K., and Olson, T. J. 2013. A new basal hybodont (Chondrichthyes, Hybodontiformes) from the Middle Permian (Roadian) Kaibab Formation, of northern Arizona. New Mexico Museum of Natural History and Science Bulletin, 60:103–108.
- Coates, M. I., and Gess, R. W., 2007, A new reconstruction of Onychoselache Traquairi, comments on early Chondrichthyan pectoral girdles and hybodontiform phylogeny: Palaeontology, v. 50, no. 6, p. 1421-1446.
- Peecook, Brandon R.; Bronson, Allison W.; Otoo, Benjamin K.A.; Sidor, Christian A. (November 2021). "Freshwater fish faunas from two Permian rift valleys of Zambia, novel additions to the ichthyofauna of southern Pangea". Journal of African Earth Sciences. 183: 104325. Bibcode:2021JAfES.18304325P. doi:10.1016/j.jafrearsci.2021.104325.
- Cione, Alberto Luis; Gouiric-Cavalli, Soledad; Mennucci, Jorge Augusto; Cabrera, Daniel Alfredo; Freije, Rubén Hugo (2010). "First vertebrate body remains from the Permian of Argentina (Elasmobranchii and Actinopterygii)". Proceedings of the Geologists' Association. 121 (3): 301–312. Bibcode:2010PrGA..121..301C. doi:10.1016/j.pgeola.2010.04.003.
- Wen, Wen; Zhang, Qiyue; Benton, Michael J.; Kriwet, Jürgen; Hu, Shixue; Huang, Jinyuan; Zhou, Changyong; Cui, Xindong; Ma, Zhixin; Min, Xiao (February 2023). "First occurrence of hybodontid teeth in the Luoping Biota (Middle Triassic, Anisian), emphasizing recovery of the marine ecosystem after the end-Permian mass extinction". Palaeogeography, Palaeoclimatology, Palaeoecology. 617: 111471. doi:10.1016/j.palaeo.2023.111471.
- Maisey, J. G., 2012, What is an ‘elasmobranch’? The impact of palaeontology in understanding elasmobranch phylogeny and evolution: Journal of Fish Biology, v. 80, no. 5, p. 918-951.
- Cappetta, H., Buffetaut, E., Cuny, G., and Suteethorn, V., 2006, A new Elasmobranch assemblage from the Lower Cretaceous of Thailand Palaeontology, v. 49, no. 3, p. 547-555.
- Fischer, J. A. N., Voigt, S., Schneider, J. W., Buchwitz, M., and Voigt, S., 2011, A selachian freshwater fauna from the Triassic of Kyrgyzstan and its implication for Mesozoic shark nurseries: Journal of Vertebrate Paleontology, v. 31, no. 5, p. 937-953.
- Coates, Michael I.; Gess, Robert W. (2007). "A New Reconstruction of Onychoselache Traquairi, Comments on Early Chondrichthyan Pectoral Girdles and Hybodontiform Phylogeny". Palaeontology. 50 (6): 1421–1446. Bibcode:2007Palgy..50.1421C. doi:10.1111/j.1475-4983.2007.00719.x. S2CID 140556654.
External links
- https://web.archive.org/web/20081106004709/http://palaeos.com/Vertebrates/Units/070Chondrichthyes/070.600.html#Hybodontiformes
- http://www.helsinki.fi/~mhaaramo/metazoa/deuterostoma/chordata/chondrichthyes/elasmobranchii/hybodontiformes/hybodontiformes.html
| Forerunners |
|
| |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jawless fish |
| ||||||||||||
| Jawed fish |
| ||||||||||||
| Lists | |||||||||||||
| Related | |||||||||||||
† extinct | |||||||||||||
