Extracellular matrix
In biology, the extracellular matrix (ECM), also called intercellular matrix,[1][2] is a network consisting of extracellular macromolecules and minerals, such as collagen, enzymes, glycoproteins and hydroxyapatite that provide structural and biochemical support to surrounding cells.[3][4][5] Because multicellularity evolved independently in different multicellular lineages, the composition of ECM varies between multicellular structures; however, cell adhesion, cell-to-cell communication and differentiation are common functions of the ECM.[6]
| Extracellular matrix | |
|---|---|
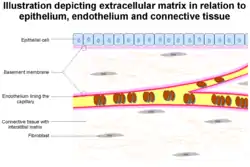 Illustration depicting extracellular matrix (basement membrane and interstitial matrix) in relation to epithelium, endothelium and connective tissue | |
| Details | |
| Identifiers | |
| Latin | Matrix extracellularis |
| Acronym(s) | ECM |
| MeSH | D005109 |
| TH | H2.00.03.0.02001 |
| Anatomical terms of microanatomy | |
The animal extracellular matrix includes the interstitial matrix and the basement membrane.[7] Interstitial matrix is present between various animal cells (i.e., in the intercellular spaces). Gels of polysaccharides and fibrous proteins fill the interstitial space and act as a compression buffer against the stress placed on the ECM.[8] Basement membranes are sheet-like depositions of ECM on which various epithelial cells rest. Each type of connective tissue in animals has a type of ECM: collagen fibers and bone mineral comprise the ECM of bone tissue; reticular fibers and ground substance comprise the ECM of loose connective tissue; and blood plasma is the ECM of blood.
The plant ECM includes cell wall components, like cellulose, in addition to more complex signaling molecules.[9] Some single-celled organisms adopt multicellular biofilms in which the cells are embedded in an ECM composed primarily of extracellular polymeric substances (EPS).[10]
Structure
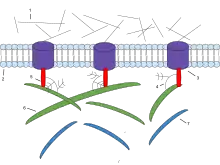
Components of the ECM are produced intracellularly by resident cells and secreted into the ECM via exocytosis.[11] Once secreted, they then aggregate with the existing matrix. The ECM is composed of an interlocking mesh of fibrous proteins and glycosaminoglycans (GAGs).
Proteoglycans
Glycosaminoglycans (GAGs) are carbohydrate polymers and mostly attached to extracellular matrix proteins to form proteoglycans (hyaluronic acid is a notable exception; see below). Proteoglycans have a net negative charge that attracts positively charged sodium ions (Na+), which attracts water molecules via osmosis, keeping the ECM and resident cells hydrated. Proteoglycans may also help to trap and store growth factors within the ECM.
Described below are the different types of proteoglycan found within the extracellular matrix.
Heparan sulfate
Heparan sulfate (HS) is a linear polysaccharide found in all animal tissues. It occurs as a proteoglycan (PG) in which two or three HS chains are attached in close proximity to cell surface or ECM proteins.[12][13] It is in this form that HS binds to a variety of protein ligands and regulates a wide variety of biological activities, including developmental processes, angiogenesis, blood coagulation, and tumour metastasis.
In the extracellular matrix, especially basement membranes, the multi-domain proteins perlecan, agrin, and collagen XVIII are the main proteins to which heparan sulfate is attached.
Chondroitin sulfate
Chondroitin sulfates contribute to the tensile strength of cartilage, tendons, ligaments, and walls of the aorta. They have also been known to affect neuroplasticity.[14]
Keratan sulfate
Keratan sulfates have a variable sulfate content and, unlike many other GAGs, do not contain uronic acid. They are present in the cornea, cartilage, bones, and the horns of animals.
Hyaluronic acid
Hyaluronic acid (or "hyaluronan") is a polysaccharide consisting of alternating residues of D-glucuronic acid and N-acetylglucosamine, and unlike other GAGs, is not found as a proteoglycan. Hyaluronic acid in the extracellular space confers upon tissues the ability to resist compression by providing a counteracting turgor (swelling) force by absorbing significant amounts of water. Hyaluronic acid is thus found in abundance in the ECM of load-bearing joints. It is also a chief component of the interstitial gel. Hyaluronic acid is found on the inner surface of the cell membrane and is translocated out of the cell during biosynthesis.[15]
Hyaluronic acid acts as an environmental cue that regulates cell behavior during embryonic development, healing processes, inflammation, and tumor development. It interacts with a specific transmembrane receptor, CD44.[16]
Collagen
Collagens are the most abundant protein in the ECM. In fact, collagen is the most abundant protein in the human body[17][18] and accounts for 90% of bone matrix protein content.[19] Collagens are present in the ECM as fibrillar proteins and give structural support to resident cells. Collagen is exocytosed in precursor form (procollagen), which is then cleaved by procollagen proteases to allow extracellular assembly. Disorders such as Ehlers Danlos Syndrome, osteogenesis imperfecta, and epidermolysis bullosa are linked with genetic defects in collagen-encoding genes.[11] The collagen can be divided into several families according to the types of structure they form:
- Fibrillar (Type I, II, III, V, XI)
- Facit (Type IX, XII, XIV)
- Short chain (Type VIII, X)
- Basement membrane (Type IV)
- Other (Type VI, VII, XIII)
Elastin
Elastins, in contrast to collagens, give elasticity to tissues, allowing them to stretch when needed and then return to their original state. This is useful in blood vessels, the lungs, in skin, and the ligamentum nuchae, and these tissues contain high amounts of elastins. Elastins are synthesized by fibroblasts and smooth muscle cells. Elastins are highly insoluble, and tropoelastins are secreted inside a chaperone molecule, which releases the precursor molecule upon contact with a fiber of mature elastin. Tropoelastins are then deaminated to become incorporated into the elastin strand. Disorders such as cutis laxa and Williams syndrome are associated with deficient or absent elastin fibers in the ECM.[11]
Extracellular vesicles
In 2016, Huleihel et al., reported the presence of DNA, RNA, and Matrix-bound nanovesicles (MBVs) within ECM bioscaffolds.[20] MBVs shape and size were found to be consistent with previously described exosomes. MBVs cargo includes different protein molecules, lipids, DNA, fragments, and miRNAs. Similar to ECM bioscaffolds, MBVs can modify the activation state of macrophages and alter different cellular properties such as; proliferation, migration and cell cycle. MBVs are now believed to be an integral and functional key component of ECM bioscaffolds.
Cell adhesion proteins
Fibronectin
Fibronectins are glycoproteins that connect cells with collagen fibers in the ECM, allowing cells to move through the ECM. Fibronectins bind collagen and cell-surface integrins, causing a reorganization of the cell's cytoskeleton to facilitate cell movement. Fibronectins are secreted by cells in an unfolded, inactive form. Binding to integrins unfolds fibronectin molecules, allowing them to form dimers so that they can function properly. Fibronectins also help at the site of tissue injury by binding to platelets during blood clotting and facilitating cell movement to the affected area during wound healing.[11]
Laminin
Laminins are proteins found in the basal laminae of virtually all animals. Rather than forming collagen-like fibers, laminins form networks of web-like structures that resist tensile forces in the basal lamina. They also assist in cell adhesion. Laminins bind other ECM components such as collagens and nidogens.[11]
Development
There are many cell types that contribute to the development of the various types of extracellular matrix found in the plethora of tissue types. The local components of ECM determine the properties of the connective tissue.
Fibroblasts are the most common cell type in connective tissue ECM, in which they synthesize, maintain, and provide a structural framework; fibroblasts secrete the precursor components of the ECM, including the ground substance. Chondrocytes are found in cartilage and produce the cartilaginous matrix. Osteoblasts are responsible for bone formation.
Physiology
Stiffness and elasticity
The ECM can exist in varying degrees of stiffness and elasticity, from soft brain tissues to hard bone tissues. The elasticity of the ECM can differ by several orders of magnitude. This property is primarily dependent on collagen and elastin concentrations,[4] and it has recently been shown to play an influential role in regulating numerous cell functions.
Cells can sense the mechanical properties of their environment by applying forces and measuring the resulting backlash.[21] This plays an important role because it helps regulate many important cellular processes including cellular contraction,[22] cell migration,[23] cell proliferation,[24] differentiation[25] and cell death (apoptosis).[26] Inhibition of nonmuscle myosin II blocks most of these effects,[25][23][22] indicating that they are indeed tied to sensing the mechanical properties of the ECM, which has become a new focus in research during the past decade.
Effect on gene expression
Differing mechanical properties in ECM exert effects on both cell behaviour and gene expression.[27] Although the mechanism by which this is done has not been thoroughly explained, adhesion complexes and the actin-myosin cytoskeleton, whose contractile forces are transmitted through transcellular structures are thought to play key roles in the yet to be discovered molecular pathways.[22]
Effect on differentiation
ECM elasticity can direct cellular differentiation, the process by which a cell changes from one cell type to another. In particular, naive mesenchymal stem cells (MSCs) have been shown to specify lineage and commit to phenotypes with extreme sensitivity to tissue-level elasticity. MSCs placed on soft matrices that mimic brain differentiate into neuron-like cells, showing similar shape, RNAi profiles, cytoskeletal markers, and transcription factor levels. Similarly stiffer matrices that mimic muscle are myogenic, and matrices with stiffnesses that mimic collagenous bone are osteogenic.[25]
Durotaxis
Stiffness and elasticity also guide cell migration, this process is called durotaxis. The term was coined by Lo CM and colleagues when they discovered the tendency of single cells to migrate up rigidity gradients (towards more stiff substrates)[23] and has been extensively studied since. The molecular mechanisms behind durotaxis are thought to exist primarily in the focal adhesion, a large protein complex that acts as the primary site of contact between the cell and the ECM.[28] This complex contains many proteins that are essential to durotaxis including structural anchoring proteins (integrins) and signaling proteins (adhesion kinase (FAK), talin, vinculin, paxillin, α-actinin, GTPases etc.) which cause changes in cell shape and actomyosin contractility.[29] These changes are thought to cause cytoskeletal rearrangements in order to facilitate directional migration.
Function
Due to its diverse nature and composition, the ECM can serve many functions, such as providing support, segregating tissues from one another, and regulating intercellular communication. The extracellular matrix regulates a cell's dynamic behavior. In addition, it sequesters a wide range of cellular growth factors and acts as a local store for them.[7] Changes in physiological conditions can trigger protease activities that cause local release of such stores. This allows the rapid and local growth factor-mediated activation of cellular functions without de novo synthesis.
Formation of the extracellular matrix is essential for processes like growth, wound healing, and fibrosis. An understanding of ECM structure and composition also helps in comprehending the complex dynamics of tumor invasion and metastasis in cancer biology as metastasis often involves the destruction of extracellular matrix by enzymes such as serine proteases, threonine proteases, and matrix metalloproteinases.[7][30]
The stiffness and elasticity of the ECM has important implications in cell migration, gene expression,[31] and differentiation.[25] Cells actively sense ECM rigidity and migrate preferentially towards stiffer surfaces in a phenomenon called durotaxis.[23] They also detect elasticity and adjust their gene expression accordingly which has increasingly become a subject of research because of its impact on differentiation and cancer progression.[32]
In the brain, where hyaluronan is the main ECM component, the matrix display both structural and signaling properties. High-molecular weight hyaluronan acts as a diffusional barrier that can modulate diffusion in the extracellular space locally. Upon matrix degradation, hyaluronan fragments are released to the extracellular space, where they function as pro-inflammatory molecules, orchestrating the response of immune cells such as microglia.[33]
Cell adhesion
Many cells bind to components of the extracellular matrix. Cell adhesion can occur in two ways; by focal adhesions, connecting the ECM to actin filaments of the cell, and hemidesmosomes, connecting the ECM to intermediate filaments such as keratin. This cell-to-ECM adhesion is regulated by specific cell-surface cellular adhesion molecules (CAM) known as integrins. Integrins are cell-surface proteins that bind cells to ECM structures, such as fibronectin and laminin, and also to integrin proteins on the surface of other cells.
Fibronectins bind to ECM macromolecules and facilitate their binding to transmembrane integrins. The attachment of fibronectin to the extracellular domain initiates intracellular signalling pathways as well as association with the cellular cytoskeleton via a set of adaptor molecules such as actin.[8]
Clinical significance
Extracellular matrix has been found to cause regrowth and healing of tissue. Although the mechanism of action by which extracellular matrix promotes constructive remodeling of tissue is still unknown, researchers now believe that Matrix-bound nanovesicles (MBVs) are a key player in the healing process.[20][34] In human fetuses, for example, the extracellular matrix works with stem cells to grow and regrow all parts of the human body, and fetuses can regrow anything that gets damaged in the womb. Scientists have long believed that the matrix stops functioning after full development. It has been used in the past to help horses heal torn ligaments, but it is being researched further as a device for tissue regeneration in humans.[35]
In terms of injury repair and tissue engineering, the extracellular matrix serves two main purposes. First, it prevents the immune system from triggering from the injury and responding with inflammation and scar tissue. Next, it facilitates the surrounding cells to repair the tissue instead of forming scar tissue.[35]
For medical applications, the required ECM is usually extracted from pig bladders, an easily accessible and relatively unused source. It is currently being used regularly to treat ulcers by closing the hole in the tissue that lines the stomach, but further research is currently being done by many universities as well as the U.S. Government for wounded soldier applications. As of early 2007, testing was being carried out on a military base in Texas. Scientists are using a powdered form on Iraq War veterans whose hands were damaged in the war.[36]
Not all ECM devices come from the bladder. Extracellular matrix coming from pig small intestine submucosa are being used to repair "atrial septal defects" (ASD), "patent foramen ovale" (PFO) and inguinal hernia. After one year, 95% of the collagen ECM in these patches is replaced by the normal soft tissue of the heart.[37]
Extracellular matrix proteins are commonly used in cell culture systems to maintain stem and precursor cells in an undifferentiated state during cell culture and function to induce differentiation of epithelial, endothelial and smooth muscle cells in vitro. Extracellular matrix proteins can also be used to support 3D cell culture in vitro for modelling tumor development.[38]
A class of biomaterials derived from processing human or animal tissues to retain portions of the extracellular matrix are called ECM Biomaterial.
In plants
Plant cells are tessellated to form tissues. The cell wall is the relatively rigid structure surrounding the plant cell. The cell wall provides lateral strength to resist osmotic turgor pressure, but it is flexible enough to allow cell growth when needed; it also serves as a medium for intercellular communication. The cell wall comprises multiple laminate layers of cellulose microfibrils embedded in a matrix of glycoproteins, including hemicellulose, pectin, and extensin. The components of the glycoprotein matrix help cell walls of adjacent plant cells to bind to each other. The selective permeability of the cell wall is chiefly governed by pectins in the glycoprotein matrix. Plasmodesmata (singular: plasmodesma) are pores that traverse the cell walls of adjacent plant cells. These channels are tightly regulated and selectively allow molecules of specific sizes to pass between cells.[15]
In Pluriformea and Filozoa
The extracellular matrix functionality of animals (Metazoa) developed in the common ancestor of the Pluriformea and Filozoa, after the Ichthyosporea diverged.[39]
History
The importance of the extracellular matrix has long been recognized (Lewis, 1922), but the usage of the term is more recent (Gospodarowicz et al., 1979).[40][41][42][43]
References
- "Matrix - Definition and Examples - Biology Online Dictionary". 24 December 2021.
- "Body Tissues | SEER Training". training.seer.cancer.gov. Retrieved 12 January 2023.
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK (February 2016). "Extracellular matrix structure". Advanced Drug Delivery Reviews. 97: 4–27. doi:10.1016/j.addr.2015.11.001. PMID 26562801.
- Bonnans C, Chou J, Werb Z (December 2014). "Remodelling the extracellular matrix in development and disease". Nature Reviews. Molecular Cell Biology. 15 (12): 786–801. doi:10.1038/nrm3904. PMC 4316204. PMID 25415508.
- Michel G, Tonon T, Scornet D, Cock JM, Kloareg B (October 2010). "The cell wall polysaccharide metabolism of the brown alga Ectocarpus siliculosus. Insights into the evolution of extracellular matrix polysaccharides in Eukaryotes". The New Phytologist. 188 (1): 82–97. doi:10.1111/j.1469-8137.2010.03374.x. PMID 20618907.

- Abedin M, King N (December 2010). "Diverse evolutionary paths to cell adhesion". Trends in Cell Biology. 20 (12): 734–42. doi:10.1016/j.tcb.2010.08.002. PMC 2991404. PMID 20817460.
- Kumar; Abbas; Fausto (2005). Robbins and Cotran: Pathologic Basis of Disease (7th ed.). Philadelphia: Elsevier. ISBN 978-0-7216-0187-8.
- Alberts B, Bray D, Hopin K, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2004). "Tissues and Cancer". Essential cell biology. New York and London: Garland Science. ISBN 978-0-8153-3481-1.
- Brownlee, Colin (October 2002). "Role of the extracellular matrix in cell-cell signalling: paracrine paradigms". Current Opinion in Plant Biology. 5 (5): 396–401. doi:10.1016/S1369-5266(02)00286-8. PMID 12183177.
- Kostakioti M, Hadjifrangiskou M, Hultgren SJ (April 2013). "Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era". Cold Spring Harbor Perspectives in Medicine. 3 (4): a010306. doi:10.1101/cshperspect.a010306. PMC 3683961. PMID 23545571.
- Plopper G (2007). The extracellular matrix and cell adhesion, in Cells (eds Lewin B, Cassimeris L, Lingappa V, Plopper G). Sudbury, MA: Jones and Bartlett. ISBN 978-0-7637-3905-8.
- Gallagher JT, Lyon M (2000). "Molecular structure of Heparan Sulfate and interactions with growth factors and morphogens". In Iozzo RV (ed.). Proteoglycans: structure, biology and molecular interactions. Marcel Dekker Inc. New York, New York. pp. 27–59. ISBN 9780824703349.
- Iozzo RV (1998). "Matrix proteoglycans: from molecular design to cellular function". Annual Review of Biochemistry. 67 (1): 609–52. doi:10.1146/annurev.biochem.67.1.609. PMID 9759499. S2CID 14638091.

- Hensch, Takao K. (2005). "Critical Period Mechanisms in Developing Visual Cortex". Neural Development. Current Topics in Developmental Biology. Vol. 69. pp. 215–237. doi:10.1016/S0070-2153(05)69008-4. ISBN 978-0-12-153169-0. PMID 16243601.
- Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J (2008). "Integrating Cells Into Tissues". Molecular Cell Biology (5th ed.). New York: WH Freeman and Company. pp. 197–234.
- Peach RJ, Hollenbaugh D, Stamenkovic I, Aruffo A (July 1993). "Identification of hyaluronic acid binding sites in the extracellular domain of CD44". The Journal of Cell Biology. 122 (1): 257–64. doi:10.1083/jcb.122.1.257. PMC 2119597. PMID 8314845.
- Di Lullo GA, Sweeney SM, Korkko J, Ala-Kokko L, San Antonio JD (February 2002). "Mapping the ligand-binding sites and disease-associated mutations on the most abundant protein in the human, type I collagen". The Journal of Biological Chemistry. 277 (6): 4223–31. doi:10.1074/jbc.M110709200. PMID 11704682.
- Karsenty G, Park RW (1995). "Regulation of type I collagen genes expression". International Reviews of Immunology. 12 (2–4): 177–85. doi:10.3109/08830189509056711. PMID 7650420.
- Kern B, Shen J, Starbuck M, Karsenty G (March 2001). "Cbfa1 contributes to the osteoblast-specific expression of type I collagen genes". The Journal of Biological Chemistry. 276 (10): 7101–7. doi:10.1074/jbc.M006215200. PMID 11106645.
- Huleihel L, Hussey GS, Naranjo JD, Zhang L, Dziki JL, Turner NJ, Stolz DB, Badylak SF (June 2016). "Matrix-bound nanovesicles within ECM bioscaffolds". Science Advances. 2 (6): e1600502. Bibcode:2016SciA....2E0502H. doi:10.1126/sciadv.1600502. PMC 4928894. PMID 27386584.
- Plotnikov SV, Pasapera AM, Sabass B, Waterman CM (December 2012). "Force fluctuations within focal adhesions mediate ECM-rigidity sensing to guide directed cell migration". Cell. 151 (7): 1513–27. doi:10.1016/j.cell.2012.11.034. PMC 3821979. PMID 23260139.
- Discher DE, Janmey P, Wang YL (November 2005). "Tissue cells feel and respond to the stiffness of their substrate". Science. 310 (5751): 1139–43. Bibcode:2005Sci...310.1139D. CiteSeerX 10.1.1.318.690. doi:10.1126/science.1116995. PMID 16293750. S2CID 9036803.
- Lo CM, Wang HB, Dembo M, Wang YL (July 2000). "Cell movement is guided by the rigidity of the substrate". Biophysical Journal. 79 (1): 144–52. Bibcode:2000BpJ....79..144L. doi:10.1016/S0006-3495(00)76279-5. PMC 1300921. PMID 10866943.
- Hadjipanayi E, Mudera V, Brown RA (February 2009). "Close dependence of fibroblast proliferation on collagen scaffold matrix stiffness". Journal of Tissue Engineering and Regenerative Medicine. 3 (2): 77–84. doi:10.1002/term.136. PMID 19051218. S2CID 174311.
- Engler AJ, Sen S, Sweeney HL, Discher DE (August 2006). "Matrix elasticity directs stem cell lineage specification". Cell. 126 (4): 677–89. doi:10.1016/j.cell.2006.06.044. PMID 16923388. S2CID 16109483.
- Wang HB, Dembo M, Wang YL (November 2000). "Substrate flexibility regulates growth and apoptosis of normal but not transformed cells". American Journal of Physiology. Cell Physiology. 279 (5): C1345-50. doi:10.1152/ajpcell.2000.279.5.C1345. PMID 11029281.
- Wahbi, Wafa; Naakka, Erika; Tuomainen, Katja; Suleymanova, Ilida; Arpalahti, Annamari; Miinalainen, Ilkka; Vaananen, Juho; Grenman, Reidar; Monni, Outi; Al-Samadi, Ahmed; Salo, Tuula (February 2020). "The critical effects of matrices on cultured carcinoma cells: Human tumor-derived matrix promotes cell invasive properties". Experimental Cell Research. 389 (1): 111885. doi:10.1016/j.yexcr.2020.111885. hdl:10138/325579. PMID 32017929. S2CID 211035510.
- Allen JL, Cooke ME, Alliston T (September 2012). "ECM stiffness primes the TGFβ pathway to promote chondrocyte differentiation". Molecular Biology of the Cell. 23 (18): 3731–42. doi:10.1091/mbc.E12-03-0172. PMC 3442419. PMID 22833566.
- Kanchanawong P, Shtengel G, Pasapera AM, Ramko EB, Davidson MW, Hess HF, Waterman CM (November 2010). "Nanoscale architecture of integrin-based cell adhesions". Nature. 468 (7323): 580–4. Bibcode:2010Natur.468..580K. doi:10.1038/nature09621. PMC 3046339. PMID 21107430.
- Liotta LA, Tryggvason K, Garbisa S, Hart I, Foltz CM, Shafie S (March 1980). "Metastatic potential correlates with enzymatic degradation of basement membrane collagen". Nature. 284 (5751): 67–8. Bibcode:1980Natur.284...67L. doi:10.1038/284067a0. PMID 6243750. S2CID 4356057.
- Wang JH, Thampatty BP, Lin JS, Im HJ (April 2007). "Mechanoregulation of gene expression in fibroblasts". Gene. 391 (1–2): 1–15. doi:10.1016/j.gene.2007.01.014. PMC 2893340. PMID 17331678.
- Provenzano PP, Inman DR, Eliceiri KW, Keely PJ (December 2009). "Matrix density-induced mechanoregulation of breast cell phenotype, signaling and gene expression through a FAK-ERK linkage". Oncogene. 28 (49): 4326–43. doi:10.1038/onc.2009.299. PMC 2795025. PMID 19826415.
- Soria FN, Paviolo C, Doudnikoff E, Arotcarena ML, Lee A, Danné N, Mandal AK, Gosset P, Dehay B, Groc L, Cognet L, Bezard E (July 2020). "Synucleinopathy alters nanoscale organization and diffusion in the brain extracellular space through hyaluronan remodeling". Nature Communications. 11 (1): 3440. Bibcode:2020NatCo..11.3440S. doi:10.1038/s41467-020-17328-9. PMC 7351768. PMID 32651387.
- "Pitt researchers solve mystery on how regenerative medicine works". EurekAlert!. Retrieved 2017-03-01.
- 'Pixie dust' helps man grow new finger
- HowStuffWorks, Humans Can Regrow Fingers? In 2009, the St. Francis Heart Center announced the use of the extracellular matrix technology in repair surgery. Archived March 10, 2007, at the Wayback Machine
- "First Ever Implantation of Bioabsorbable Biostar Device at DHZB". DHZB NEWS. December 2007. Archived from the original on 2008-12-11. Retrieved 2008-08-05.
The almost transparent collagen matrix consists of medically purified pig intestine, which is broken down by the scavenger cells (macrophages) of the immune system. After about 1 year the collagen has been almost completely (90-95%) replaced by normal body tissue: only the tiny metal framework remains. An entirely absorbable implant is currently under development.
- Kleinman HK, Luckenbill-Edds L, Cannon FW, Sephel GC (October 1987). "Use of extracellular matrix components for cell culture". Analytical Biochemistry. 166 (1): 1–13. doi:10.1016/0003-2697(87)90538-0. PMID 3314585.
- Tikhonenkov, Denis V. (2020). "Insights into the origin of metazoan multicellularity from predatory unicellular relatives of animals". BMC Biology. 18 (39): 39. doi:10.1186/s12915-020-0762-1. PMC 7147346. PMID 32272915.
- Lewis WH (1922). "The adhesive quality of cells". Anat Rec. 23 (7): 387–392. doi:10.1002/ar.1090230708. S2CID 84566330.
- Gospodarowicz D, Vlodovsky I, Greenburg G, Johnson LK (1979). "Cellular shape is determined by the extracellular matrix and is responsible for the control of cellular growth and function". In Sato GH, Ross R (eds.). Hormones and Cell Culture. Coldspring Harbor Laboratory. p. 561. ISBN 9780879691257.
- Mecham R, ed. (2011). The extracellular matrix: an overview. Springer. ISBN 9783642165559.
- Rieger R, Michaelis A, Green MM (2012-12-06). Glossary of Genetics: Classical and Molecular (5th ed.). Berlin: Springer-Verlag. p. 553. ISBN 9783642753336.
Further reading
- Extracellular matrix: review of its roles in acute and chronic wounds
- Usage of Extracellular Matrix from pigs to regrow human extremities
- Sound Medicine - Heart Tissue Regeneration - July 19 interview discussing ECM and its uses in cardiac tissue repair (requires MP3 playback).