Kayenta Formation
The Kayenta Formation is a geological formation in the Glen Canyon Group that is spread across the Colorado Plateau province of the United States, including northern Arizona, northwest Colorado, Nevada, and Utah. Traditionally has been suggested as Sinemurian-Pliensbachian, but more recent dating of detrital zircons has yielded a depositional age of 183.7 ± 2.7 Ma, thus a Pliensbachian-Toarcian age is more likely.[2] A previous depth work recovered a solid "Carixian" (Lower-Middle Pliensbachian) age from measurements done in the Tenney Canyon.[3] More recent works have provided varied datations for the layers, with samples from Colorado and Arizona suggesting 197.0±1.5-195.2±5.5 Ma (Middle Sinemurian), while the topmost section is likely Toarcian or close in age, maybe even recovering terrestrial deposits coeval with the Toarcian Oceanic Anoxic Event.[4] This last age asignation also correlated the Toarcian Vulcanism on the west Cordilleran Magmatic Arc, as the number of grains from this event correlate with the silt content in the sandstones of the upper layers.[4]
| Kayenta Formation | |
|---|---|
| Stratigraphic range: Sinemurian-Toarcian ~ | |
 Kayenta Formation, Capitol Reef National Park, Utah. | |
| Type | Geological formation |
| Unit of | Glen Canyon Group |
| Underlies | Navajo Sandstone |
| Overlies | Wingate Sandstone |
| Thickness | 100 to 120 metres (330 to 390 ft) |
| Lithology | |
| Primary | Sandstone |
| Other | Siltstone, Limestone |
| Location | |
| Coordinates | 37.8°N 110.6°W |
| Country | United States |
| Extent | northern Arizona, northwest Colorado, Nevada, and Utah[1] |
| Type section | |
| Named for | Kayenta, Arizona |
.jpg.webp)
This rock formation is particularly prominent in southeastern Utah, where it is seen in the main attractions of a number of national parks and monuments. These include Zion National Park, Capitol Reef National Park, the San Rafael Swell, and Canyonlands National Park.
The Kayenta Formation frequently appears as a thinner dark broken layer below Navajo Sandstone and above Wingate Sandstone (all three formations are in the same group). Together, these three formations can result in immense vertical cliffs of 2,000 feet (610 m) or more. Kayenta layers are typically red to brown in color, forming broken ledges.
Kayenta Formation in Utah
Southeast Utah
_Navajo_Sandstone_(Lower_Jurassic)_(Kolob_Canyons%252C_Zion_National_Park%252C_Utah%252C_USA)_(8425013742).jpg.webp)

In most sections that include all three geologic formations of the Glen Canyon group the Kayenta is easily recognized. Even at a distance it appears as a dark-red, maroon, or lavender band of thin-bedded material between two thick, massive, cross bedded strata of buff, tan, or light-red color. Its position is also generally marked by a topographic break. Its weak beds form a bench or platform developed by stripping the Navajo sandstone back from the face of the Wingate cliffs. The Kayenta is made up of beds of sandstone, shale, and limestone, all lenticular, uneven at their tops, and discontinuous within short distances. They suggest deposits made by shifting streams of fluctuating volume. The sandstone beds, from less than 1-inch (25 mm) to more than 10 feet (3.0 m) thick, are composed of relatively coarse, well-rounded quartz grains cemented by lime and iron. The thicker beds are indefinitely cross bedded. The shales are essentially fine-grained, very thin sandstones that include lime concretions and balls of consolidated mud. The limestone appears as solid gray-blue beds, a few inches to a few feet thick, and as lenses of limestone conglomerate. Most of the limestone lenses are less than 25 feet (7.6 m) long, but two were traced for nearly 500 feet (150 m) and one for 1,650 feet (500 m).
Viewed as a whole, the Kayenta is readily distinguished from the geologic formations above and below it. It is unlike them in composition, color, manner of bedding, and sedimentary history. Obviously the conditions of sedimentation changed in passing from the Wingate Sandstone formation to the Kayenta and from the Kayenta to the Navajo sandstone, but the nature and regional significance of the changes have not been determined. In some measured sections the transition from Wingate to Kayenta is gradual; the material in the basal Kayenta, beds seems to have been derived from the Wingate immediately below and redeposited with only the discordance characteristic of fluviatile sediments. But in many sections the contact between the two formations is unconformable; the basal Kayenta consists of conglomerate and lenticular sandstone that fills depressions eroded in the underlying beds. In Moqui Canyon near Red Cone Spring nearly 10 feet (3.0 m) of Kayenta limestone conglomerate rests in a long meandering valley cut in Wingate. Likewise, the contact between the Kayenta and the Navajo in places seems to be gradational, but generally a thin jumbled mass of sandstone and shales, chunks of shale and limestone, mud balls, and concretions of lime and iron, lies at the base of the fine-grained, cross bedded Navajo. Mud cracks, a few ripple marks, and incipient drainage channels were observed in the topmost bed of the Kayenta on Red Rock Plateau; and in west Glen Canyon, wide sand-filled cracks appear at the horizon. These features indicate that, in places at least, the Wingate and Kayenta were exposed to erosion before their overlying geologic formations were deposited, are it may be that the range in thickness of the Kayenta thus in part (is) accounted for.
Southwest Utah
The red and mauve Kayenta siltstones and sandstones that form the slopes at base of the Navajo Sandstone cliffs record the record of low to moderate energy streams. Poole (1997) has shown that the streams still flowed toward the east depositing from 150 to 210 m (500 to about 700 feet) of sediment here. The sedimentary structures showing the channel and flood plain deposits of streams are well exposed on switchbacks below the tunnel in Pine Creek Canyon.
In the southeastern part of Zion National Park a stratum of cross bedded sandstone is found roughly halfway between the top and bottom of the Kayenta Formation. It is a "tongue" of sandstone that merges with the Navajo formation east of Kanab, and it shows that desert conditions occurred briefly in this area during Kayenta time. This tongue is the ledge that shades the lower portion of the Emerald Pool Trail, and it is properly called Navajo, not Kayenta.
Fossil mudcracks attest to occasional seasonal climate, and thin limestones and fossilized trails of aquatic snails or worms mark the existence of ponds and lakes. The most interesting fossils, however, are the dinosaur tracks that are relatively common in Kayenta mudstone.
These vary in size, but all seem to be the tracks of three-toed reptiles that walked upright, leaving their tracks in the muds on the flood plains. Unfortunately, so far no bone materials have been found in Washington County that would enable more specific identification.
Apparently during Kayenta time Zion was situated in a climatic belt like that of Senegal with rainy summers and dry winters at the southern edge of a great desert. The influence of the desert was about to predominate, however, as North America drifted northward into the arid desert belt.
Glen Canyon
The Kayenta Formation is approximately 400 feet (120 m) thick and consists of a fine-grained sandstone interbedded with layers of siltstone. The alternation of these units generally produces a series of ledges and slopes between the cliffs of the Navajo and Moenave formation. Dinosaur tracks are fairly common in the siltstone, and fresh water mussels and snails occur but are rare. The Kayenta Formation is colored pale red and adds to the splendor of the Vermilion Cliffs. It accumulated as deposits of rivers.
Fossils
Color key
|
Notes Uncertain or tentative taxa are in small text; |
Invertebrates
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
|
Base of the "typical facies" |
Valves |
A freshwater (lacustrine or fluvial) ostracod, type member of the family Darwinulidae. Kayenta specimens have a distinctive columnar calcitic layer. The Kayenta fauna is similar to the Sinemurian faunas of the Toutunhe Formation of Xinjiang, China. |
||
|
Liratina[5] |
|
|
Base of the "typical facies" |
Single shell |
A freshwater (lacustrine or fluvial) snail, incertae sedis inside Mesogastropoda. Differs considerably from the species from the Upper Jurassic Morrison Formation, Liratina jurassicum |
|
|
|
Base of the "typical facies" |
Shells |
A freshwater (lacustrine or fluvial) snail, member of the family Lymnaeidae. |
||
|
Scabriculocypris[5] |
Scabriculocypris n. sp |
|
Base of the "typical facies" |
Valves |
A freshwater (lacustrine or fluvial) ostracod, incertae sedis inside Cypridacea. Differs from all other described species of the genus in being more elongate and from most in being spinose. |
|
|
Unio[7] |
|
|
Base of the "typical facies" |
Shells |
A freshwater (lacustrine or fluvial) Bivalve, member of the family Unionidae. |
|
|
|
Base of the "typical facies" |
Shells |
A freshwater (lacustrine or fluvial) snail, member of the family Valvatidae. |
||
Fishes
The "Kayenta Fish Fauna" is the last one recovered from the Glen Canyon Group sequence and it is delimited mostly to the silty facies of the Lower-Middle Part of the formation.[8] This Fauna is rather scarce and delimited to several concrete locations with proper lacustrine or fluvial deposition, and are also scarce due to preservation bias.[8] Another aspect that can explain the lack of fish fossils found is the use of different research techniques than used on the Chinle Formation.[8]
Chondrichthyes
| Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Hybodontoidea indeterminate |
|
Base of the "typical facies" |
UCMP 136104, 136105 + ten uncatalogued specimens, teeth |
A freshwater (lacustrine or fluvial) non-neoselachian shark, incertae sedis inside Hybodontoidea. The remains of sharks are rather rare on the formation and limited to several locations with typical lacustrine or fluvial floodplain deposition. |
||
|
Toarcibatidae (= "Archaeobatidae")[10] |
Toarcibatidae indeterminate |
Gold Spring Quarry 1 |
Base of the "typical facies" |
Isolated Tooth |
A freshwater (lacustrine or fluvial) toarcibatid. Related originally with Micropristis or Libanopristis, and stated to be reworked from younger Cretaceous deposits, was found due to its asymmetrical cusp to fit within the definition of Toarcibatis, being more likely to be native of the formation.[10] |
|
Actinopterygii
| Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Lophionotus[11] |
Lophionotus kanabensis |
|
Base of the "typical facies" |
|
A freshwater (lacustrine or fluvial) semionotid semionotiform. |
|
|
Palaeoniscidae indeterminate |
|
Base of the "typical facies" |
|
A freshwater (lacustrine or fluvial) palaeoniscid palaeonisciform. |
||
|
Semionotidae indeterminate |
|
Base of the "typical facies" |
|
A freshwater (lacustrine or fluvial) semionotid semionotiform, probably related to the genus Semionotus. Semionotiformes are the only properly identified bony fishes from the formation, including a large specimen exposed at the Dan O’Laurie Museum.[8] |
 Semionotus is probably related to the Kayenta Seminotiformes | |
Sarcopterygii
| Genus / Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Ceratodus stewarti |
|
Middle “Silty Facies” |
|
A freshwater (lacustrine or fluvial) ceratodontid dipnomorph (lungfish). |
| |
|
Coelacanthidae indeterminate |
|
Base of the "typical facies" |
Uncertain Remains |
A freshwater (lacustrine or fluvial) coelacanthid Coelacanthiform. Coelacanths are quoted from this zone, but their remains have not been studied. |
||
|
Potamoceratodus guentheri |
|
Middle “Silty Facies” |
|
A freshwater (lacustrine or fluvial) ceratodontid dipnomorph (lungfish). Was described as C. felchi, know from the Upper Jurassic Morrison Formation. Other dipnoan specimens have been cited but never described.[14] |
||

Amphibia
| Genus / Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Anura Indeterminate |
Gold Spring Quarry 1 |
Silty Facies Member |
MCZ 9019, distal humerus; MCZ 9020-24, ilia |
An early frog, incertae sedis relationships |
||
|
Eocaecilia micropodia |
Gold Spring Quarry 1 |
Silty Facies Member |
MNA V8066 (type); MNA V8053, 8054, 8055, 8057, 8058, 8059, 8060, 8062, 8063, 8064, 8065, 8066, 8067, 8068, 8069, 8070, 8071, 8072, 8073, 9346; MCZ 9011A, 9015, 9095, 9152, 9156. 9158, 9163, 9166, 9167, 9169A, 9171A, 9173, 9231A, 9233, 9235, 9237, 9238A, 9241, 9242 |
A genus whose relationships are controversial, being considered one of the earliest gymnophionans as a close relative of caecilians |
| |
|
Gold Spring Quarry 1 |
Silty Facies Member |
|
Incertae sedis relationships |
||
|
Prosalirus bitis |
Gold Spring Quarry 1 |
Silty Facies Member |
MNA V 8725 (type), MCZ 9324 A, MCZ 9323 A |
An early frog, probably related to Notobatrachidae |
| |
|
Urodela indeterminate |
Gold Spring Quarry 1 |
Silty Facies Member |
MCZ 9017, 9018, atlas vertebrae |
A possible stem-salamander, incertae sedis inside Urodela. The oldest record of an urodelan from North America |
||


Rhynchocephalia
| Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Navajosphenodon sani |
|
Silty Facies |
|
An Advanced Sphenodont, member of Sphenodontinae. The skeleton of N. sani shows a large number of similarities with the modern tuatara S. punctatus, clustering them closely together in the morphospace of sphenodontians and early lepidosaurs. |
| |
|
Rhynchocephalia Indeterminate |
Airhead West |
Silty Facies |
Uncertain Fragments |
Rhynchocephalians of uncertain assignment |
||
|
Sphenodontia Indeterminate |
Gold Spring Quarry 1 |
Silty Facies |
MCZ 9036 through 9040, jaw fragments |
An indeterminate sphenodont |
||
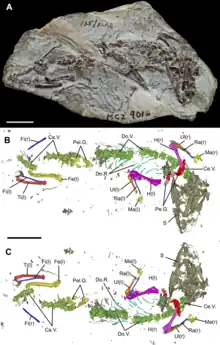
Testudinatans
| Genus / taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Cryptodira indeterminate |
Red Knob |
Silty Facies |
Uncertain fragments |
Cryptodirans of uncertain assignment |
||
|
|
Silty Facies Member |
|
| ||
|
Testudinata indeterminate |
|
Silty Facies |
Uncertain fragments |
Testudinatans of uncertain assignment |
||

Crocodylomorphs
| Genus / Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Calsoyasuchus valliceps |
Adeii Eechii Cliffs, Navajo Nation |
Silty Facies |
TMM 43631-1 (holotype), partial skull |
A terrestrial member of the Hsisosuchidae. Alternatively can be a relative of Thalattosuchia |
||
|
|
Silty Facies |
|
Indeterminate crocodylomorphs. Includes a new taxon with skull similar to Orthosuchus stormbergi. |
||
|
Eopneumatosuchus colberti |
Blue layer, Silty Facies |
Silty Facies |
|
An early terrestrial or semiterrestrial protosuchid crocodylomorph |
||
|
Kayentasuchus walkeri |
|
Silty Facies |
|
An early terrestrial or semiterrestrial crocodylomorph |
||
|
|
Silty Facies |
|
Early terrestrial or semi-terrestrial herbivorous Crocodylomorphs. Includes two taxa similar to the Cretaceous Edentosuchus tienshanensis, one that has been referred to informally as "Gomphosuchus" (including UCMP 97638 and UCMP 125871) and another unnamed taxon (including UCMP 130082).[25][26] Previously considered one taxon (the 'Kayenta form') in older literature. |
||
Dinosauria
Indeterminate ornithischian remains located in Arizona, USA.[27] Ornithischian tracks located in Arizona, USA.[27] Indeterminate theropod remains located in Arizona, US.[27] Theropod tracks located in Arizona and Utah, US.[27] Possible theropod tracks located in Arizona, Colorado, and Utah, US.[27]
| Genus / Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Unnamed, informally known as the "Shake-N-Bake" coelophysid |
|
Silty Facies |
|
A coelophysid neotheropod. |
||
|
Silty Facies |
|
A coelophysid neotheropod. Referred to as Syntarsus by Weishampel et al.[27] Formerly known as Megapnosaurus. |
| ||
| D. wetherilli |
|
Silty Facies |
|
An advanced neotheropod, type member of the family Dilophosauridae. Dilophosaurus is the main identified dinosaur from the formation, being both the most known and studied. It was among the largest theropods present locally, and very likely an active hunter, rather than a fisher.[32] |
| |
|
Heterodontosauridae indeterminate |
Gold Springs |
Silty Facies |
MCZ 9092, Partial Skull & Associated Remains (complete upper and lower dentitions, many other portions of the skull, vertebrae from all portions of the axial column, and portions of fore and hind girdles and limbs)[34] |
A heterodontosaurid of uncertain placement. Its presence in North America suggest early dispersal of the family. |
||
|
Kayentavenator elysiae[35] |
Willow Springs |
Silty Facies |
UCMP V128659, six proximal caudal centra, three centra, two partial neural arches, fragmentary ilium, proximal pubes, pubic shaft fragments, incomplete femora, proximal tibiae, proximal fibula, fragments |
A neotheropod of uncertain relationships, probably a coelophysoid. Originally referred to M. kayentakatae by Rowe. |
| |
|
|
Silty Facies |
|
A uncertain placement large ornithischian and teeth from diverse type of genera. The femur was assigned to Dilophosaurus wetherilli.[36] The femur resembles that of the early neornithischian Lesothosaurus.[37] |
||
|
Sarahsaurus aurifontanalis[27] |
|
Silty Facies |
|
A sauropodomorph, a member of the family Massospondylidae. Originally thought to be Massospondylus[27] |
| |
|
S. sp. (S. "arizonensis") |
Valley of the Buttes |
Silty Facies |
UCMP 130056, scutes |
A controversial thyreophoran, resembles the osteoderms of S. harrisonii. |
||
|
Scutellosaurus lawleri |
|
Silty Facies |
|
A basal thyreophoran, the most abundant dinosaur of the formation |
| |
|
Theropoda indeterminate |
|
Silty Facies |
|
Incertae sedis within Theropoda, probably Neotheropoda |
||




Pterosauria
| Genus / Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Pterosauria? Indeterminate |
|
Silty Facies |
|
Possible pterosaur teeth |
||
|
Rhamphinion jenkinsi[43] |
|
Silty Facies, Ward Mesa |
|
A pterosaur, considered a member of the family Dimorphodontidae. Was originally classified as a "rhamphorhynchoid", represents the only major pterosaur identified from the formation and one of the oldest from North America. |
||
Synapsida
| Genus / Taxon | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
|
Silty Facies |
|
A relatively large and common tritylodont |
||
|
Dinnetherium nezorum |
|
Silty Facies |
MNA V3221; MCZ 20870-20877 |
A mammaliaform, member of the family Megazostrodontidae |
||
|
Haramiyidae? indet. |
|
Silty Facies |
MCZ 20879 |
Incertae sedis, a possible haramiyid |
||
|
|
Silty Facies |
|
A Large tritylodont, with a suggested semiaquatic mode of life. A specimen has been recovered with several associated perinates.[47] |
| |
|
Morganucodon sp. |
|
Silty Facies |
MCZ 20878 |
A mammaliaform, member of the family Morganucodontidae |
| |
|
|
Silty Facies |
|
A tritylodont, also present on coeval deposits from Asia and Europe. |
| |
|
Tritylodontidae indeterminate |
|
Silty Facies |
Teeth |
Indeterminate tritylodontid remains |
||


Ichnofossils
| Genus | Species | Location | Material | Type | Origin | Notes | Images |
|---|---|---|---|---|---|---|---|
|
|
Footprints |
Moving Tracks |
|
Ornithischian Footprints of the ichnofamily Moyenisauropodidae. |
||
|
|
Tracks |
Moving Tracks |
|
Pseudosuchia Footprints of the ichnofamily Batrachopodidae. |
||
|
|
Tracks |
Moving Trails |
|
Dinosaur Traces left while swimming |
||
|
|
Footprints |
Moving Tracks |
|
Theropod Footprints of the ichnofamily Grallatoridae. |
||
|
|
Footprints |
Moving Tracks |
|
Possible Dinosaur Footprints, non assigned to any concrete ichnogenus |
 Example of Indeterminate Dinosaur Footprint from the Kayenta Formation | |
|
|
Footprints |
Moving Tracks |
|
Theropod Footprints of the ichnofamily Grallatoridae. This type of tracks match with Dilophosaurus pes |
||
|
|
Footprints |
Moving Tracks |
|
Theropod Footprints of the ichnofamily Grallatoridae. Likely from smaller local theropods |
||
|
|
Footprints |
Moving Tracks |
|
Theropod Footprints of the ichnofamily Grallatoridae. |
||
|
|
Trackways |
Moving Tracks |
|
Saltwater/Blackish-linked tracks with resemblance with extant Xiphosuran traces |
||
|
|
Footprints |
Moving Tracks |
|
Ornithischian Footprints of the ichnofamily Moyenisauropodidae. |
||
|
|
Footprints |
Moving Tracks |
|
Theropod Footprints of the ichnofamily Otozoidae. Includes tracks referable to bipedal Sauropodomorphs |
||
|
|
Cylindrical burrows |
Pascichnia |
|
Burrow-like ichnofossils. It is referred to vermiform deposit-feeders. It is controversial, since is considered a strictly a junior synonym of Palaeophycus.[63] |
 Example of Planolites fossil | |
|
|
Cylindrical to subcylindrical Burrows |
Domichnia |
|
Burrow-like ichnofossils. Ichnofossils done by organisms advancing along the bottom surface. Very narrow, vertical or subvertical, slightly winding unlined shafts filled with mud. Interpreted as dwelling structures of vermiform animals, more concretely the Domichnion of a suspension-feeding Worm or Phoronidan, with certain Skolithos representing entrance shafts to more complicated burrows. |
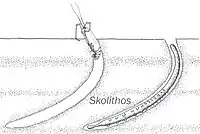 Skolithos ichnofosil reconstruction, with possible fauna associated | |
|
Synapsidipedia[49] |
|
|
Tracks |
Moving Tracks |
|
Possible Synapsid Footprints, non assigned to any concrete ichnogenus |
|
|
|
Unlined meniscate burrows |
Fodinichnia |
|
Saltwater/Blackish burrow-like ichnofossils. Taenidium is a meniscate backfill structure, usually considered to be produced by an animal progressing axially through the sediment and depositing alternating packets of differently constituted sediment behind it as it moves forward. |
||
|
|
Footprints |
Moving Tracks |
|
Possible Theropod Footprints, non assigned to any concrete ichnogenus |
 Example of Indeterminate Theropod Footprint from the Kayenta Formation | |
|
|
Trails |
Moving Trails |
|
Fish-swimming fossil trail left as a fossil impression on a substrate |
||
Plants
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Clathropteris[13] |
|
|
|
Affinities with Dipteridaceae inside Polypodiales. |
 Example of Clathropteris meniscioides specimen |
|
|
|
Affinities with Cycadidae inside Cycadopsida. |
 Example of extant cycad, Encephalartos longifolius | |
|
|
|
Affinities with Williamsoniaceae inside Bennettitales. |
||
|
|
|
|||
|
|
|
A member of Williamsoniaceae inside Bennettitales. It has been interpreted as a cycad in the family Cycadaceae or a Bennettitalean plant, and also a late surviving member of Noeggerathiales. |
 Example of Zamites mandelslohi specimen | |
See also
- Kayenta, Arizona
- List of dinosaur-bearing rock formations
- List of fossil sites (with link directory)
Footnotes
- Weishampel, David B; et al. (2004). "Dinosaur distribution (Early Jurassic, North America)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. pp. 530–532. ISBN 0-520-24209-2.
- Marsh, A.D.; Rowe, T.; Simonetti, A.; Stockli, D.; Stockli, L. (2014). "The age of the Kayenta Formation of northeastern Arizona: overcoming the challenges of dating fossil bone". J. Vertebr. Paleontol. Prog. Abst. 34 (2): 178. Retrieved 19 November 2021.
- Steiner, M.; Tanner, L.H. (2014). "Magnetostratigraphy and paleopoles of the Kayenta Formation and the Tenney Canyon Tongue" (PDF). Volumina Jurassica. 12: 31–38. Retrieved 7 March 2022.
- Marsh, Adam Douglas (2018). "Contextualizing the evolution of theropod dinosaurs in western North America using U-Pb geochronology of the Chinle Formation and Kayenta Formation on the Colorado Plateau". doi:10.26153/tsw/41876.
{{cite journal}}: Cite journal requires|journal=(help) - Kietzke, K.K.; Lucas, S.G. (1995). "Ostracoda and Gastropoda from the Kayenta Formation (Lower Jurassic) of Arizona, U.S.A". Journal of Arizona–Nevada Academy of Science. 28 (1–2): 23–32. JSTOR 40024298. Retrieved 19 November 2021.
- Lewis, G. E.; Irwin, J. H.; Wilson, R. F. (1961). "Age of the Glen Canyon Group (Triassic and Jurassic) on the Colorado Plateau". Geological Society of America Bulletin. 72 (9): 1437–1440. Bibcode:1961GSAB...72.1437L. doi:10.1130/0016-7606(1961)72[1437:AOTGCG]2.0.CO;2. Retrieved 2 January 2022.
- Harshbarger, J.W.; Repenning, C.A.; Irwin, J.H. (1957). "Stratigraphy of the uppermost Triassic and the Jurassic rocks of the Navajo Country" (PDF). United States Geological Survey Professional Paper. 291 (1): 1–74. Retrieved 2 January 2022.
- Milner, A. R.; Kirkland, J. I.; Birthisel, T. A. (2006). "The geographic distribution and biostratigraphy of Late Triassic–Early Jurassic freshwater fish faunas of the southwestern United States". New Mexico Museum of Natural History and Science Bulletin. 37 (1): 522–529. Retrieved 19 November 2021.
- Curtis, K.; Padian, K. (1999). "An Early Jurassic microvertebrate fauna from the Kayenta Formation of northeastern Arizona: microfaunal change across the Triassic‐Jurassic boundary". PaleoBios. 19 (1): 19–37.
- Delsate, D.; Candoni, L. (2001). "Description de nouveaux morphotypes dentaires de Batomorphii toarciens (Jurassique inférieur) du Bassin de Paris: Archaeobatidae nov. fam". Bulletin-Société des Naturalistes Luxembourgeois. 102 (1): 131–143. Retrieved 19 November 2021.
- Milner, A. R. C.; Birthisel, T. A.; Kirkland, J. I.; Breithaupt, B.H; Matthews, N. A.; Lockley, M. G.; Santucci, V. L.; Gibson, S. Z; DeBlieux, D. D.; Hurlbut, M.; Harris, J. D.; Olsen, P. E. (2011). "Tracking Early jurassic dinosaurs across southwestern Utah and the Triassic-Jurassic transition". Nevada State Museum Paleontological Papers. 1 (1): 1–107.
- Gay, R. J.; Milner, A. R. (2015). "The first report of an archosaur from the Kayenta Formation of Washington County, Utah" (PDF). PeerJ PrePrints. 1048 (1): 1–21. Retrieved 19 November 2021.
- Milner, A. R.; Gay, R. J.; Irmis, R.; Overkamp, F.; Santella, M. (2017). "New southwestern Utah paleontological locality from the Lower Jurassic Kayenta Formation reveals a diverse vertebrate fauna based on teeth and tracks [abs.]". Journal of Vertebrate Paleontology. 37 (1): 164.
- Milner, A. R.; Kirkland, J. I. (2006). "Preliminary review of the early Jurassic (Hettangian) freshwater Lake Dixie fish fauna in the Whitmore Point Member, Moenave Formation in southwest Utah". New Mexico Museum of Natural History and Science Bulletin. 37 (1–2): 510–521. CiteSeerX 10.1.1.537.1492.
- Frederickson, Joseph A; Cifelli, Richard L. (2017). "New Cretaceous lungfishes (Dipnoi, Ceratodontidae) from western North America". Journal of Paleontology. 91 (1): 146–161. doi:10.1017/jpa.2016.131. S2CID 131962612.
- Jenkins, F. A.; Walsh, D. M. (1993). "An Early Jurassic caecilian with limbs". Nature. 365 (2–4): 246–250. Bibcode:1993Natur.365..246J. doi:10.1038/365246a0. S2CID 4342438. Retrieved 19 November 2021.
- Shubin, N. H.; Jenkins, F. A. (1995). "An early Jurassic jumping frog". Nature. 377 (6544): 49–52. Bibcode:1995Natur.377...49S. doi:10.1038/377049a0. S2CID 4308225. Retrieved 19 November 2021.
- Simões, Tiago R.; Kinney-Broderick, Grace; E. Pierce, Stephanie (2022). "An exceptionally preserved Sphenodon-like sphenodontian reveals deep time conservation of the tuatara skeleton and ontogeny". Communications Biology. 5 (1): 195–208. doi:10.1038/s42003-022-03144-y. PMC 8894340. PMID 35241764. S2CID 247227560.
 Text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - Padian, K. (1984). "Pterosaur remains from the Kayenta Formation (Early Jurassic) of Arizona". Palaeontology. 27 (2): 407–413. Retrieved 19 November 2021.
- Gafney, E.; Hutchinson, H.; Jenkins, F.; Meeker, L. (1987). "Modern turtle origins; the oldest known cryptodire". Science. 237 (2–5): 289–291. Bibcode:1987Sci...237..289G. doi:10.1126/science.237.4812.289. PMID 17772056. S2CID 36112523. Retrieved 19 November 2021.
- Tykoski, Ronald S; Rowe, Timothy B.; Ketcham, Richard A; Colbert, Matthew W. (2002). "Calsoyasuchus valliceps, a new crocodyliform from the Early Jurassic Kayenta Formation of Arizona" (PDF). Journal of Vertebrate Paleontology. 22 (3): 593–611. doi:10.1671/0272-4634(2002)022[0593:CVANCF]2.0.CO;2. S2CID 85969174. Retrieved 19 November 2021.
- Crompton, A. W.; Smith, K. K. (1980). Jacobs, L. (ed.). "A new genus and species from the Kayenta Formation (Late Triassic?) of Northern Arizona". Aspects of Vertebrate History. Museum of Northern Arizona Press. 34 (1): 193–217. Retrieved 19 November 2021.
- Clark, J. M.; Sues, Hans-Dieter (2002). "Two new species of basal crocodylomorphs and the status of the Sphenosuchia". Zoological Journal of the Linnean Society. 136 (1–6): 77−96. doi:10.1046/j.1096-3642.2002.00026.x. S2CID 83511047.
- Ősi, Attila (2014). "The evolution of jaw mechanism and dental function in heterodont crocodyliforms". Historical Biology. 26 (3): 279–414. doi:10.1080/08912963.2013.777533. S2CID 85707749. Retrieved 24 November 2021.
- Ruebenstahl, A. A.; Klein, M. D.; Yi, H.; Xu, X.; Clark, J. M. (2022). "Anatomy and relationships of the early diverging Crocodylomorphs Junggarsuchus sloani and Dibothrosuchus elaphros". The Anatomical Record. 305 (Special Issue): 2463–2556. doi:10.1002/ar.24949. PMC 9541040. PMID 35699105. S2CID 249645515.
- Melstrom, K. M.; Irmis, R. B. (2019). "Repeated evolution of herbivorous crocodyliforms during the age of dinosaurs". Current Biology. 29 (14): 2389–95. doi:10.1016/j.cub.2019.05.076. PMID 31257139. S2CID 195699188.
- Weishampel, David B; et al. (2004). "Dinosaur distribution (Early Jurassic, North America)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. pp. 530–32. ISBN 0-520-24209-2.
- Tykoski, R.S. (1997). "A new ceratosaurid theropod from the Early Jurassic Kayenta Formation of Northern Arizona". Journal of Vertebrate Paleontology. 17 (3): 81–82.
- Rowe, T. (1989). "A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta Formation of Arizona" (PDF). Journal of Vertebrate Paleontology. 9 (2): 125–36. doi:10.1080/02724634.1989.10011748. Retrieved 19 November 2021.
- Bristowe, A.; M.A. Raath (2004). "A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus.(USA)". Palaeontologica Africana. 40 (40): 31–41.
- Welles, S.P. (1954). "New Jurassic dinosaur from the Kayenta Formation of Arizona". Bulletin of the Geological Society of America. 65 (1): 591–98. Bibcode:1954GSAB...65..591W. doi:10.1130/0016-7606(1954)65[591:NJDFTK]2.0.CO;2. Retrieved 19 November 2021.
- Marsh, Adam D.; Rowe, Timothy B. (7 July 2020). "A comprehensive anatomical and phylogenetic evaluation of Dilophosaurus wetherilli (Dinosauria, Theropoda) with descriptions of new specimens from the Kayenta Formation of northern Arizona". Journal of Paleontology. 94 (S78): 1–103. doi:10.1017/jpa.2020.14. ISSN 0022-3360. S2CID 220601744.
- Attridge, J.; Crompton, A.W.; Jenkins, F.A .Jr (1985). "The southern African Liassic prosauropod Massospondylus discovered in North America". J Vertebr Paleontol. 5 (1): 128–32. doi:10.1080/02724634.1985.10011850. Retrieved 19 November 2021.
- Sereno, Paul C. (2012-10-03). "Taxonomy, morphology, masticatory function and phylogeny of heterodontosaurid dinosaurs". ZooKeys (223): 1–225. doi:10.3897/zookeys.223.2840. ISSN 1313-2989. PMC 3491919. PMID 23166462.
- Gay, Robert J. (2010). "Kayentavenator elysiae, a new Tetanuran from the Early Jurassic of Arizona". Notes on Early Jurassic Theropods. 1 (1): 23–36.
- Breeden, Benjamin T.; Raven, Thomas J.; Butler, Richard J.; Rowe, Timothy B.; Maidment, Susannah C. R. (2021). "The anatomy and palaeobiology of the early armoured dinosaur Scutellosaurus lawleri (Ornithischia: Thyreophora) from the Kayenta Formation (Lower Jurassic) of Arizona". Royal Society Open Science. 8 (7): 2016–76. Bibcode:2021RSOS....801676B. doi:10.1098/rsos.201676. PMC 8292774. PMID 34295511.
- Breeden, Benjamin T. (2018). "Fragmentary specimens provide evidence for hidden taxonomic diversity of ornithischian dinosaurs within the Lower Jurassic Kayenta Formation (northeastern Arizona, USA)". SVP 2018. 1 (1): 96.
- Marsh, Adam D.; Rowe, Timothy B. (2018-10-10). "Anatomy and systematics of the sauropodomorph Sarahsaurus aurifontanalis from the Early Jurassic Kayenta Formation". PLOS ONE. 13 (10): e0204007. Bibcode:2018PLoSO..1304007M. doi:10.1371/journal.pone.0204007. ISSN 1932-6203. PMC 6179219. PMID 30304035.
- Padian, K. (1989). "Presence of the dinosaur Scelidosaurus indicates Jurassic age for the Kayenta Formation (Glen Canyon Group, northern Arizona)". Geology. 17 (5): 438–41. Bibcode:1989Geo....17..438P. doi:10.1130/0091-7613(1989)017<0438:POTDSI>2.3.CO;2. Retrieved 19 November 2021.
- Norman, David B (2020-01-27). "Scelidosaurus harrisonii from the Early Jurassic of Dorset, England: the dermal skeleton". Zoological Journal of the Linnean Society. 190 (1): 1–53. doi:10.1093/zoolinnean/zlz085. ISSN 0024-4082.
- Breeden, B. T.; Rowe, T. B. (2020). "New specimens of Scutellosaurus lawleri Colbert,1981, from the Lower Jurassic Kayenta Formation in Arizona elucidate the early evolution of thyreophoran dinosaurs". Journal of Vertebrate Paleontology. 40 (4): e1791894. doi:10.1080/02724634.2020.1791894. S2CID 224961326. Retrieved 19 November 2021.
- Rosenbaum, J. N.; Padian, Kevin (2000). "New material of the basal thyreophoran Scutellosaurus lawleri from the Kayenta Formation (Lower Jurassic) of Arizona". PaleoBios. 20 (1): 13–23. Retrieved 19 November 2021.
- Wellnhofer, Peter (1991). Summary of Lower Jurassic Pterosaurs. The Illustrated Encyclopedia of Pterosaurs (1 ed.). London: Salamander Books Limited. p. 79. ISBN 0-86101-566-5.
- Hans-Dieter, Sues (1986). "Dinnebitodon amarali, a new tritylodontid (Synapsida) from the Lower Jurassic of western North America". Journal of Paleontology. 60 (3): 758–62. doi:10.1017/S0022336000022277. S2CID 131801988. Retrieved 19 November 2021.
- Jenkins, F. A.; Crompton, A. W.; Downs, W. R. (1983). "Mesozoic mammals from Arizona: new evidence on mammalian evolution". Science. 222 (4629): 1233–35. Bibcode:1983Sci...222.1233J. doi:10.1126/science.222.4629.1233. PMID 17806725. S2CID 35607107. Retrieved 19 November 2021.
- Hans-Dieter, Sues; Jenkins, F. A. (2006). "The Postcranial Skeleton of Kayentatherium Wellesi from the Lower Jurassic Kayenta Formation of Arizona and the Phylogenetic Significance of Postcranial Features in Tritylodontid Cynodonts". Carrano, Matthew T., Gaudin, T. J., Blob, R. W. And Wible, J. R.: Amniote Paleobiology. Perspectivers on the Evolution of Mammals, Birds, and Reptiles the University of Chicago Press. 1 (1): 114–52.
- Hoffman, E. A.; Rowe, T. B. (2018). "Jurassic stem-mammal perinates and the origin of mammalian reproduction and growth". Nature. 561 (7721): 104–08. Bibcode:2018Natur.561..104H. doi:10.1038/s41586-018-0441-3. PMID 30158701. S2CID 205570021. Retrieved 19 November 2021.
- Palmer, D. (1999). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. p. 193. ISBN 1-84028-152-9.
- Lockley, M. G.; Hunt, A. P. (1995). Dinosaur Tracks and Other Fossil Footprints of the Western United States (2 ed.). USA: Columbia University Press. pp. 15–287. ISBN 9780231079273. Retrieved 2 January 2022.
- Milner, A.R.C.; Birthisel, T. A.; Kirkland, J. I.; Breithaupt, B.H.; Matthews, N. A.; Lockley, M. G.; Santucci, V. L.; Gibson, S. Z.; DeBlieux, D. D.; Hurlbut, M.; Harris, J. D.; Olsen, P. E. (2011). "Tracking Early jurassic dinosaurs across southwestern Utah and the Triassic-Jurassic transition". Nevada State Museum Paleontological Papers. 1 (1): 1–107.
- Miller, W. E.; Britt, B. B.; Stadtman, K. L. (1986). "Theropod and prosauropod trackways from the Moenave Formation of southwestern Utah". In D. D. Gillette (Ed.), First International Symposium on Dinosaur Tracks and Traces, Abstracts with Program. 21 (2): 36–48.
- Lockey, M.; Milner, A.C.; Hamblin, D. (2006). "Dinosaur tracksites from the Kayenta Formation (Lower Jurassic),'. Desert Tortoise site,'Washington County, Utah". New Mexico Museum of Natural History and Science Bulletin. 37 (2): 269–275. Retrieved 2 January 2022.
- Welles, S. P. (1971). "Dinosaur footprints from the Kayenta Formation of northern Arizona". Plateau. 44 (1): 27–38.
- Stokes, W. L.; Bruhn, A. F. (1960). "Dinosaur tracks from Zion National Park and Vicinity, Utah". Utah Academy Proceedings. 37 (2): 75–76.
- Barnes, F. A. (1997). "Canyon Country Dinosaur". Tracks and Trackers. 54 (1): 1–176.
- Hamblin, A. H.; Lockley, M.G.; Milner, A.R.C. (2006). "More reports of theropod dinosaur tracksites from the Kayenta Formation (Lower Jurassic), Washington County, Utah: implications for describing the Springdale megatracksite". New Mexico Museum of Natural History and Science Bulletin. 37 (3): 276–281.
- Hamblin, A. H. (2006). Spectrum Tracksite—also known as the Grapevine Pass Wash Tracksite. California State University: Desert Studies Consortium and LSA Associates.
- Lockley, M. G.; Hunt, A. P.; Meyer, C. A.; Rainforth, E. C.; Schultz, R. J. (1998). "A survey of fossil footprint sites at Glen Canyon National Recreation Area (western USA): a case study in documentation of trace fossil resources at a national preserve". Ichnos. 5 (3): 177–211. doi:10.1080/10420949809386417.
- Hamblin, A. H.; Foster, J. R. (2001). "Ancient animal footprints and traces in the Grand Staircase-Escalante National Monument, south-central Utah". Utah Geological Association Publication. 28 (4): 1–12.
- Lockey, M.G.; Gierlinski, G.D. (2014). "A new Otozoum-dominated tracksite in the Glen Canyon Group (Jurassic) of eastern Utah". New Mexico Museum of Natural History and Science Bulletin. 62 (3): 211–214. Retrieved 3 January 2022.
- Lockley, M.G.; Matthews, N.; Breithaupt, B.H.; Gierlinski, G.; Cart, K.; Hunt-Foster, R. (2018). "Large Dinosaur Tracksites in the Lower Jurassic Kayenta Formation near Moab, Utah: implications for Paleoecology". Bull N M Mus Nat Hist Sci. 79 (4): 441–449. Retrieved 3 January 2022.
- Difley, R.; Ekdale, A. A. (2006). "Trace fossils and paleoenvironments of the Early Jurassic Kayenta Formation, Washington County, Utah" (PDF). Making Tracks Across the Southwest. 6 (2): 49–51. Retrieved 2 January 2022.
- Keighley, D. G.; Pickerill, R. K (1995). "The ichnotaxa Palaeophycus and Planolites_ historical perspectives and recommendations". Ichnos. 3 (4). doi:10.1080/10420949509386400.
References
- Geology of Eastern Iron County, Utah by Herbert E. Gregory Utah Geological and Mineral Survey Bulletin No. 37 (1950)
- The San Juan Country – A Geographic and Reconnaissance of Southeastern Utah by Herbert E. Gregory. U.S. Geological Survey Professional Paper No. 188
- Geology Studies, Vol. 15 Part 5 (1968)
- Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. 861 pp. ISBN 0-520-24209-2.