Killer-cell immunoglobulin-like receptor
Killer-cell immunoglobulin-like receptors (KIRs), are a family of type I transmembrane glycoproteins expressed on the plasma membrane of natural killer (NK) cells and a minority of T cells.[1][2] At least 15 genes and 2 pseudogenes encoding KIR map in a 150-kb region of the leukocyte receptor complex (LRC) on human chromosome 19q13.4.[3]
| Killer-cell immunoglobulin-like receptor | |
|---|---|
| Identifiers | |
| Symbol | KIR |
| Membranome | 18 |
They regulate the killing function of these cells by interacting with major histocompatibility (MHC) class I molecules, which are expressed on all nucleated cell types. KIR receptors can distinguish between MHC I allelic variants, which allows them to detect virally infected cells or transformed cells. KIRs are paired receptors with both activating and inhibitory functions; most KIRs are inhibitory: their recognition of MHC molecules suppresses the cytotoxic activity of their NK cell.[4]
A limited number of KIRs are activating: their recognition of MHC molecules activates the cytotoxic activity of their cell.[5] Initial expression of KIRs on NK cells is stochastic, but NK cells undergo an educational process as they mature that alters the KIR expression to maximize the balance between effective defense and self-tolerance. KIR's role in killing unhealthy self-cells and not killing healthy self-cells, involves them in protection against and propensity to viral infection, autoimmune disease, and cancer.[2][6] KIR molecules are polymorphic: their gene sequences differ greatly across individuals. They are also polygenic so that it is rare for two unrelated individuals to possess the same KIR genotype.[7]
Unlike T lymphocytes, resting NK cells use preformed lytic granules to kill target cells, implying a rapid cytolytic effect that requires a finely regulated control mechanism. The ability to spare normal tissues, but not transformed cells, is termed the "missing self" hypothesis.[8][9] This phenomenon is determined by MHC class I–specific inhibitory receptors that functionally dominate the triggering potentials induced by activating receptors [10][11] Thus, NK cells use a complex array of inhibitory or activating receptor/ligand interactions, the balance of which regulates NK cell function and cytolytic activity.[10][12][13][14][15][16] Receptors displaying this function evolved during phylogenesis following the rapid evolution of genes coding for MHC class I molecules. Thus, in primates and a few other species, evolved MHC class I–inhibitory receptors belong to the KIR immunoglobulin superfamily,[17][18][19] while in rodents and other species the same function is under the control of type II integral transmembrane glycoproteins, structurally characterized as disulfide-linked homodimers belonging to the Ly49 protein family.[20]
Function
Role in natural killer cells
Natural killer (NK) cells are a type of lymphocyte cell involved in the innate immune system's response to viral infection and tumor transformation of host cells.[19][6] Like T cells, NK cells have many qualities characteristic of the adaptive immune system, including the production of “memory” cells that persist following encounter with antigens and the ability to create a secondary recall response.[6] Unlike T cells, NK cell receptors are germline encoded, and therefore do not require somatic gene rearrangements.[6] Because NK cells target self cells, they have an intricate mechanism by which they differentiate self and non-self cells in order to minimize the destruction of healthy cells and maximize the destruction of unhealthy cells.[19]
Natural killer cell cytolysis of target cells and cytokine production is controlled by a balance of inhibitory and activating signals, which are facilitated by NK cell receptors.[19][21][22] NK cell inhibitory receptors are part of either the immunoglobulin-like (IgSF) superfamily or the C-type lectin-like receptor (CTLR) superfamily.[19][6] Members of the IgSF family include the human killer cell immunoglobulin-like receptor (KIR) and the Immunoglobulin-like transcripts (ILT).[6] CTLR inhibitory receptors include the CD94/NKG2A and the murine Ly49, which is probably analogous to the human KIR.[22][4]
Role in T cells
KIR and CD94 (CTLR) receptors are expressed by 5% of peripheral blood T cells.[11][22]
Nomenclature and classification
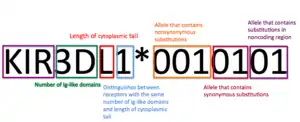
KIR receptors are named based on the number of their extracellular Ig-like domains (2D or 3D) and by the length of their cytoplasmic tail (long (L), short (S), or pseudogene (P)).[11][22] The number following the L, S, or P in the case of a pseudogene, differentiates KIR receptors with the same number of extracellular domains and length of cytoplasmic tail.[3][22] Finally, the asterisk after this nomenclature indicates allelic variants.[4][22]
Single substitutions, insertions, or deletions in the genetic material that encodes KIR receptors changes the site of termination for the gene, causing the cytoplasmic tail to be long or short, depending on the site of the stop codon.[11][3] These single nucleotide alterations in the nucleotide sequence fundamentally alter KIR function. With the exception of KIR2DL4, which has both activating and inhibitory capabilities, KIR receptors with long cytoplasmic tails are inhibitory and those with short tails are activating.[19][22]
Receptor Types
Inhibitory Receptors
Inhibitory receptors recognize self-MHC class I molecules on target self cells, causing the activation of signaling pathways that stop the cytolytic function of NK cells.[23] Self-MHC class I molecules are always expressed under normal circumstance.[19] According to the missing-self hypothesis, inhibitory KIR receptors recognize the downregulation of MHC class I molecules in virally-infected or transformed self cells, leading these receptors to stop sending the inhibition signal, which then leads to the lysis of these unhealthy cells.[19][22] Because natural killer cells target virally infected host cells and tumor cells, inhibitory KIR receptors are important in facilitating self-tolerance.[5]
KIR inhibitory receptors signal through their immunoreceptor tyrosine-based inhibitory motif (ITIM) in their cytoplasmic domain. When inhibitory KIR receptors bind to a ligand, their ITIMs are tyrosine phosphorylated and protein tyrosine phosphatases, including SHP-1, are recruited. Inhibition occurs early in the activation signaling pathway, likely through the interference of the pathway by these phosphatases.[19][22]
Activating Receptors
Activating receptors recognize ligands that indicate host cell aberration, including induced-self antigens (which are markers of infected self cells and include MICA, MICB, and ULBP, all of which are related to MHC class 1 molecules), altered-self antigens (MHC class I antigens laden with foreign peptide), and/or non-self (pathogen encoded molecules).[19][22] The binding of activating KIR receptors to these molecules causes the activation of signaling pathways that cause NK cells to lyse virally infected or transformed cells.[22]
Activating receptors do not have the immunoreceptor tyrosine-base inhibition motif (ITIM) characteristic of inhibitory receptors, and instead contain a positively charged lysine or arginine residue in their transmembrane domain (with the exception of KIR2L4) that helps to bind DAP12, an adaptor molecule containing a negatively charged residue as well as immunoreceptor tyrosine-based activation motifs (ITAM).[22][12] Activating KIR receptors include KIR2DS, KIR2DL, and KIR3DS.[22]
Much less is known about activating receptors compared to inhibitory receptors. A significant proportion of the human population lacks activating KIR receptors on the surface of their NK cells as a result of truncated variants of KIR2DS4 and 2DL4, which are not expressed on the cell surface, in individuals who are heterozygous for the KIR group A haplotype.[19] This suggests that a lack of activating KIR receptors is not incredibly detrimental, likely because there are other families of activating NK cell surface receptors that bind MHC class I molecules that are probably expressed in individuals with this phenotype. Because little is known about the function of activating KIR receptors, however, it is possible that there is an important function of activating KIR receptors of which we are not yet aware.[19]
Activating receptors have lower affinity for their ligands than do inhibitory receptors.[22] Although the purpose of this difference in affinity is unknown, it is possible that the cytolysis of target cells occurs preferentially under conditions in which the expression of stimulating MHC class I molecules on target cells is high, which may occur during viral infection.[22] This difference, which is also present in Ly49, the murine homolog to KIR, tips the balance towards self-tolerance.[16]
Expression
Activating and inhibitory KIR receptors are expressed on NK cells in patchy, variegated combinations, leading to distinct NK cells.[22] The IgSF and CTLR superfamily inhibitory receptors expressed on the surface of NK cells are each expressed on a subset of NK cells in such a way that not all classes of inhibitory NK cell receptors are expressed on each NK cell, but there is some overlap.[22] This creates unique repertories of NK cells, increasing the specificity with which NK cells recognize virally-infected and transformed self-cells.[22] Expression of KIR receptors is determined primarily by genetic factors, but recent studies have found that epigenetic mechanisms also play a role in KIR receptor expression.[22] Activating and inhibitory KIR receptors that recognize the same class I MHC molecule are mostly not expressed by the same NK cell.[22] This pattern of expression is beneficial in that target cells that lack inhibitory MHC molecules but express activating MHC molecules are extremely sensitive to cytolysis.[22]
Although initial expression of inhibitory and activating receptors on NK cells appears to be stochastic, there is an education process based on MHC class I alleles expressed by the host that determines the final repertoire of NK receptor expression.[22][11] This process of education is not well understood.[22] Different receptor genes are expressed primarily independently of other receptor genes, which substantiates the idea that initial expression of receptors is stochastic.[22] Receptors are not expressed entirely independently of each other, however, which supports the idea that there is an education process that reduces the amount of randomness associated with receptor expression. Further, once an NK receptor gene is activated in a cell, its expression is maintained for many cell generations.[11][22] It appears that some proportion of NK cells are developmentally immature and therefore lack inhibitory receptors, making them hyporesponsive to target cells.[22] In the human fetal liver, KIR and CD49 receptors are already expressed by NK cells, indicating that at least some KIR receptors are present in fetal NK cells, although more studies are needed to substantiate this idea.[22] Although the induction of NK receptor expression is not fully understood, one study found that human progenitor cells cultured in vitro with cytokines developed into NK cells, and many of these cells expressed CD94/NKG2A receptors, a CTLR receptor.[22] Moreover, there was little to no KIR receptor expression in these cells, so additional signals are clearly required for KIR induction.[22]
The balance between effective defense and self-tolerance is important to the functioning of NK cells. It is thought that NK cell self-tolerance is regulated by the educational process of receptor expression described above, although the exact mechanism is not known.[22] The “at least one” hypothesis is an attractive, though not yet fully substantiated, hypothesis that tries to explain the way in which self-tolerance is regulated in the education process. This hypothesis posits that the NK cell repertoire is regulated so that at least one inhibitory receptor (either of the IgSF or CTLR superfamily) is present on every NK cell, which would ensure self-tolerance.[22] Effective defense requires an opposing pattern of receptor expression. The co-expression of many MHC-specific receptors by NK cells is disfavored, likely because cells that co-express receptors are less able to attack virally infected or transformed cells that have down-regulated or lost one MHC molecule compared to NK cells that co-express receptors to a lesser degree.[22] Minimization of co-expression, therefore, is important for mounting an effective defense by maximizing the sensitivity of response.[22]
Structure
Gene Structure
The KIR gene cluster has approximately 150 kb and is located in the leukocyte receptor complex (LRC) on human chromosome 19q13.4.[1][23][11] KIR genes have 9 exons, which are strongly correlated with KIR receptor protein domains (leader, D0, D1, and D2, stem, transmembrane, and cytosolic domains).[19] Furthermore, the promoter regions of the KIR genes share greater than 90% sequence identity, which indicates that there is similar transcriptional regulation of KIR genes.[19]
The human killer cell immunoglobulin-like receptors superfamily (which share 35-50% sequence identity and the same fold as KIR) includes immunoglobulin-like transcripts (ILT, also known as leukocyte immunoglobulin-like receptors (LIRs)), leukocyte-associated Ig-like receptors (LAIR), paired Ig-like receptors (PIR), and gp49.[3] Moreover, it has been reported that between 12 and 17 KIR receptors have been identified.[3][22][11] There was a single ancestral gene from which all extant KIR receptor genes arose via duplications, recombinations, and mutations, and all KIR receptors share more than 90% sequence identity.[19]
Genes
Protein Structure
NK cell receptors bind directly to the MHC class I molecules on the surface of target cells.[22] Human killer cell immunoglobulin-like receptors recognize the α1 and α2 domains of class I human leukocyte antigens (HLA-A, -B, and –C), which are the human versions of MHCs.[22][11] Position 44 in the D1 domain of KIR receptors and position 80 in HLA-C are important for the specificity of KIR-HLA binding.[11]
Diversity
Allelic Diversity
All but two KIR genes (KIR2DP1 and KIR3DL3) have multiple alleles, with KIR3DL2 and KIR3DL1 having the most variations (12 and 11, respectively).[5] In total, as of 2012 there were 614 known KIR nucleotide sequences encoding 321 distinct KIR proteins.[22] Further, inhibitory receptors are more polymorphic than activating receptors.[22] The great majority (69%) of substitutions in the KIR DNA sequence are nonsynonymous, and 31% are synonymous.[5] The ratio of nonsynonymous to synonymous substitutions (dN/dS) is greater than one for every KIR and every KIR domain, indicating that positive selection is occurring.[5] Further, the 5` exons, which encode the leader peptide and the Ig-like domains, have a larger proportion of nonsynonymous substitutions than do the 3` exons, which encode the stem, transmembrane region, and the cytoplasmic tail.[5] This indicates that stronger selection is occurring on the 5` exons, which encodes the extracellular part of the KIR that binds to the MHC.[5] There is, therefore, evidence of strong selection on the KIR ligand binding sites, which is consistent with the high specificity of the KIR ligand binding site, as well as the rapid evolution of class I MHC molecules and viruses.[5][22]
Genotype and haplotype diversity
Human genomes differ in their amount of KIR genes, in their proportion of inhibitory versus activating genes, and in their allelic variations of each gene.[9][7] As a result of these polygenic and polymorphic variations, less than 2% of unrelated individuals have the same KIR genotype, and ethnic populations have broadly different KIR genotype frequencies. This incredible diversity likely reflects the pressure from rapidly evolving viruses.[22] 30 distinct haplotypes have been classified, all of which can be broadly characterized by group A and group B haplotypes.[22] The Group A haplotype has a fixed set of genes, which are KIR3DL3, 2L3, 2DP1, 2DL1, 3DP1, 2DL4, 3DL1, 2DS4, and 3DL2.[19][22] Group B haplotypes encompass all other haplotypes, and therefore have a variable set of genes, including several genes absent from group A, including KIR2DS1, 2DS2, 2DS3, 2DS5, 2DL2, 2DL5, and 3DS1.[19][22] Because group B has both gene and allelic diversity (compared to just allelic diversity in group A), group B is even more diverse than group A.[19] Four KIR genes (2DL4, 3DL2, 3DL3, AND 3DP1) are present in nearly all KIR haplotypes and as a result are known as framework genes. Inheritance of maternal and paternal haplotypes results in further diversity of individual KIR genotype.[22]
Group A only has one activating KIR receptor, whereas Group B contains many activating KIR receptors, and as a result group B haplotype carriers have a stronger response to virally infected and transformed cells.[22] As a result of the huge migrations peoples indigenous to India, Australia, and the Americas made from Africa, activating KIR receptors became advantageous to these populations, and as a result these populations acquired activating KIR receptors.[22]
A study of the genotypes of 989 individuals representing eight distinct populations found 111 distinct KIR genotypes. Individuals with the most frequent genotype, which comprised 27% of the individuals studied, are homozygous for the group A haplotype.[9] The remaining 110 KIR genotypes found in this study are either group A and group B heterozygotes or group B homozygotes (who are indistinguishable from heterozygotes by genotype alone). 41% (46) of the genotypes identified were found in only one individual, and 90% of individuals had the same 40 genotypes.[4] Clearly, there is extensive diversity in human KIR genotypes, which allows for rapid evolution in response to rapidly evolving viruses.
Role in disease
Genotypes that are inhibitory KIR receptor dominant are likely susceptible to infection and reproductive disorders but protective against autoimmune diseases, whereas activating KIR receptor dominant genotypes are likely susceptible to autoimmunity but protective against viral infection and cancer.[19][22] The relationship between inhibitory vs stimulatory KIR genotype dominance, however, is more complicated than this because diseases are so diverse and have so many different causes, and immune activation or de-activation may not be protective or harmful at every stage of disease.[19] KIR2DS2 or 2DS1, which are activating receptors, are strongly correlated with most autoimmune diseases, which is logical because activating receptors induce signaling pathways that lead to cytolysis of target cells.[19][22] Another activating receptor, KIR3DS1, is protective to hepatitis-C virus infection, is associated with slowing down of AIDs progression, and is associated with cervical cancer, which is associated with a distinct strain of HPV.[19][22] It is likely that KIR3DS1 is associated with cervical cancer despite its stimulatory nature because cervical tumors generally associate with localized inflammation.[19]
As a drug target
1-7F9 is a human monoclonal antibody that binds to KIR2DL1/2L3.[24] Very similar Lirilumab is intended for the treatment of cancers e.g. leukemia.[25][26]
See also
- NK-92, a natural killer cell line that does not express KIR
References
- Yawata M, Yawata N, Abi-Rached L, Parham P (2002). "Variation within the human killer cell immunoglobulin-like receptor (KIR) gene family". Critical Reviews in Immunology. 22 (5–6): 463–82. PMID 12803322.
- Bashirova AA, Martin MP, McVicar DW, Carrington M (2006). "The killer immunoglobulin-like receptor gene cluster: tuning the genome for defense". Annual Review of Genomics and Human Genetics. 7: 277–300. doi:10.1146/annurev.genom.7.080505.115726. PMID 16824023.
- Wende H, Colonna M, Ziegler A, Volz A (1999). "Organization of the leukocyte receptor cluster (LRC) on human chromosome 19q13.4". Mammalian Genome. 10 (2): 154–160. doi:10.1007/s003359900961. PMID 9922396. S2CID 25092393.
- Raulet DH, Vance RE, McMahon CW (2001). "Regulation of the natural killer cell receptor repertoire". Annual Review of Immunology. 19: 291–330. doi:10.1146/annurev.immunol.19.1.291. PMID 11244039.
- Vilches C, Parham P (2002). "KIR: diverse, rapidly evolving receptors of innate and adaptive immunity". Annual Review of Immunology. 20: 217–51. doi:10.1146/annurev.immunol.20.092501.134942. PMID 11861603.
- Rajalingam R (2012). "Overview of the killer cell Immunoglobulin-like receptor system". Immunogenetics. Methods in Molecular Biology. Vol. 882. pp. 3914–414. doi:10.1007/978-1-61779-842-9_23. ISBN 978-1-61779-841-2. PMID 22665247.
- Uhrberg M (January 2005). "The KIR gene family: life in the fast lane of evolution". European Journal of Immunology. 35 (1): 10–5. doi:10.1002/eji.200425743. PMID 15580655. S2CID 41486412.
- Ljunggren HG, Karre K (1985). "Host resistance directed selectively against H-2 deficient lymphoma variants. Analysis of the mechanism". Journal of Experimental Medicine. 162 (6): 1745–1759. doi:10.1084/jem.162.6.1745. PMC 2187973. PMID 3877776.
- Ljunggren HG, Karre K (1990). "In search of the "missing self". MHC molecules and NK cell recognition". Immunology Today. 11 (7): 237–244. doi:10.1016/0167-5699(90)90097-S. PMID 2201309.
- Moretta A, Bottino C, Vitale M, Pende, D, Biassoni R, Mingari MC, Moretta L (1996). "Receptors for HLA-class I molecules in human Natural Killer cells". Annual Review of Immunology. 14: 619–648. doi:10.1146/annurev.immunol.14.1.619. PMID 8717527.
- Biassoni R, Malnati MS (2018). "Human Natural Killer Receptors, Co-Receptors, and their Ligands". Current Protocols in Immunology. 121 (1): e47. doi:10.1002/cpim.47. PMID 30040219. S2CID 205710349.
- Biassoni R, Cantoni C, Pende D, Sivori S, Parolini S, Vitale M, Bottino C, Moretta A (2001). "Human natural killer cell receptors and co-receptors". Immunological Reviews. 181: 203–214. doi:10.1034/j.1600-065X.2001.1810117.x. PMID 11513142. S2CID 24523974.
- Lanier LL (2005). "NK cell recognition". Annual Review of Immunology. 23: 225–274. doi:10.1146/annurev.immunol.23.021704.115526. PMID 15771571.
- Long EO, Kim HS, Liu D, Peterson ME, Rajagopalan S (2013). "Controlling natural killer cell responses: integration of signals for activation and inhibition". Annual Review of Immunology. 31: 227–258. doi:10.1146/annurev-immunol-020711-075005. PMC 3868343.
- Parham P (2005). "MHC class I molecules and KIRs in human history, health and survival". Nature Reviews Immunology. 5 (3): 201–214. doi:10.1038/nri1570. PMID 15719024. S2CID 31182134.
- Raulet DH, Vance RE (2006). "Self-tolerance of natural killer cells". Nature Reviews Immunology. 6 (7): 520–531. doi:10.1038/nri1863. PMID 16799471. S2CID 8652332.
- Wagtmann N, Biassoni R, Cantoni C, Verdiani S, Malnati M, Vitale M, Bottino C, Moretta L, Moretta A, Long EO (1995). "Molecular clones of the p58 Natural Killer cell receptor reveal immunoglobulin-related molecules with diversity in both the extra- and intracellular domains". Immunity. 2 (5): 439–449. doi:10.1016/1074-7613(95)90025-x. PMID 7749980.
- Colonna M, Samaridis J (1995). "Cloning of immunoglobulin-superfamily members associated with HLA-C and HLA-B recognition by human natural killer cells". Science. 268 (5209): 405–408. Bibcode:1995Sci...268..405C. doi:10.1126/science.7716543. PMID 7716543.
- Andre P, Biassoni R, Colonna M, Cosman D, Lanier LL, Long EO, Lopez-Botet M, Moretta A, Moretta L, Parham P, Trowsdale J, Vivier E, Wagtmann N, Wilson MJ (2001). "New nomenclature for MHC receptors". Nature Immunology. 2 (8): 661. doi:10.1038/90589. PMID 11477395. S2CID 32201997.
- Yokoyama WM, Kim S, French AR (2004). "The dynamic life of natural killer cells". Annual Review of Immunology. 22: 405–429. doi:10.1146/annurev.immunol.22.012703.104711. PMID 15032583.
- Boyington JC, Sun PD (May 2002). "A structural perspective on MHC class I recognition by killer cell immunoglobulin-like receptors". Molecular Immunology. 38 (14): 1007–21. doi:10.1016/s0161-5890(02)00030-5. PMID 11955593.
- Parham P (Feb 2005). "Immunogenetics of killer cells immunoglobulin-like receptors". Molecular Immunology. 42 (4): 459–462. doi:10.1016/j.molimm.2004.07.027. PMID 15607799.
- Radaev S, Sun PD (2003). "Structure and function of natural killer cell surface receptors". Annual Review of Biophysics and Biomolecular Structure. 32: 93–114. doi:10.1146/annurev.biophys.32.110601.142347. PMID 12471063.
- Romagné F, André P, Spee P, Zahn S, Anfossi N, Gauthier L, et al. (September 2009). "Preclinical characterization of 1-7F9, a novel human anti-KIR receptor therapeutic antibody that augments natural killer-mediated killing of tumor cells". Blood. 114 (13): 2667–77. doi:10.1182/blood-2009-02-206532. PMC 2756126. PMID 19553639.
- World Health Organization (2012). "International Nonproprietary Names for Pharmaceutical Substances (INN). Proposed INN: List 107" (PDF). WHO Drug Information. 26 (2).
- Statement On A Nonproprietary Name Adopted By The USAN Council - Lirilumab, American Medical Association.
External links
- http://www.KIRtyping.info
- The KIR Gene Cluster, NIH Bookshelf, PDF
- killer+cell+immunoglobulin-like+receptor at the U.S. National Library of Medicine Medical Subject Headings (MeSH)