Last universal common ancestor
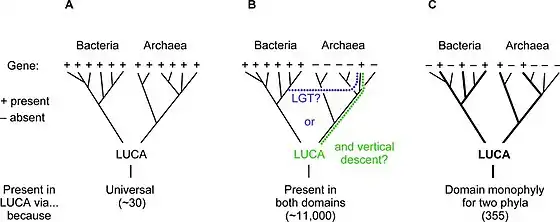
The last universal common ancestor (LUCA) is mostly hypothesized to have been a common ancestral cell[2] from which the three domains of life,[1] the Bacteria, the Archaea, and the Eukarya originated. It is suggested to have been a "cellular organism that had a lipid bilayer and used DNA, RNA, and protein".[3][2] The LUCA has also been defined as "a hypothetical organism ancestral to all three domains".[4] In general, the LUCA is considered the point or stage at which the three domains of life diverged from precursing forms of life (about 3.5 - 3.8 billion years ago). The nature of this point or stage of divergence remains a topic of research.[4]
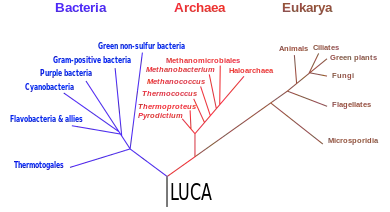
All earlier forms of life precursing this divergence and, of course, all extant terrestrial organisms are generally thought to share common ancestry. On the basis of a formal statistical test,[5] this theory of a universal common ancestry (UCA) is supported versus competing multiple-ancestry hypotheses.
The genesis of viruses, before or after the LUCA as well as the diversity of extant viruses and their hosts are subjects of investigation.[3]
While no specific fossil evidence of the LUCA exists, the detailed biochemical similarity of all current life (the existing three domains) makes it plausible. Its characteristics can be inferred from shared features of modern genomes. These genes describe a complex life form with many co-adapted features, including transcription and translation mechanisms to convert information from DNA to mRNA to proteins. The earliest forms of life probably lived in the high-temperature water of deep sea vents near ocean-floor magma flows around 4 billion years ago.
Historical background

A phylogenetic tree directly portrays the idea of evolution by descent from a single ancestor.[6] An early tree of life was sketched by Jean-Baptiste Lamarck in his Philosophie zoologique in 1809.[7][8] Charles Darwin more famously proposed the theory of universal common descent through an evolutionary process in his book On the Origin of Species in 1859: "Therefore I should infer from analogy that probably all the organic beings which have ever lived on this earth have descended from some one primordial form, into which life was first breathed."[9] The last sentence of the book begins with a restatement of the hypothesis:
There is grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one ...
— [9]
The term "last universal common ancestor" or "LUCA" was first used in the 1990s for such a primordial organism.[10][11][12]
Inferring LUCA's features
An anaerobic thermophile
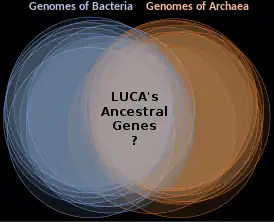
In 2016, Madeline C. Weiss and colleagues genetically analyzed 6.1 million protein-coding genes and 286,514 protein clusters from sequenced prokaryotic genomes representing many phylogenetic trees, and identified 355 protein clusters that were probably common to the LUCA. The results "depict LUCA as anaerobic, CO2-fixing, H2-dependent with a Wood–Ljungdahl pathway (the reductive acetyl-coenzyme A pathway), N2-fixing and thermophilic. LUCA's biochemistry was replete with FeS clusters and radical reaction mechanisms."[14] The cofactors also reveal "dependence upon transition metals, flavins, S-adenosyl methionine, coenzyme A, ferredoxin, molybdopterin, corrins and selenium. Its genetic code required nucleoside modifications and S-adenosylmethionine-dependent methylations."[14] The results are rather specific: they show that methanogenic clostridia was basal, near the root of the phylogenetic tree, in the 355 protein lineages examined, and that the LUCA may therefore have inhabited an anaerobic hydrothermal vent setting in a geochemically active environment rich in H2, CO2, and iron, where ocean water interacted with hot magma beneath the ocean floor.[14] It's even inferred that LUCA also grew from H2 and CO2 via the reverse incomplete Krebs cycle.[15] Other metabolic pathways inferred in LUCA are the pentose phosphate pathway, glycolysis, and gluconeogenesis.[16]
While the gross anatomy of the LUCA can only be reconstructed with much uncertainty, its biochemical mechanisms can be described in some detail, based on the "universal" properties currently shared by all independently living organisms on Earth.[17]
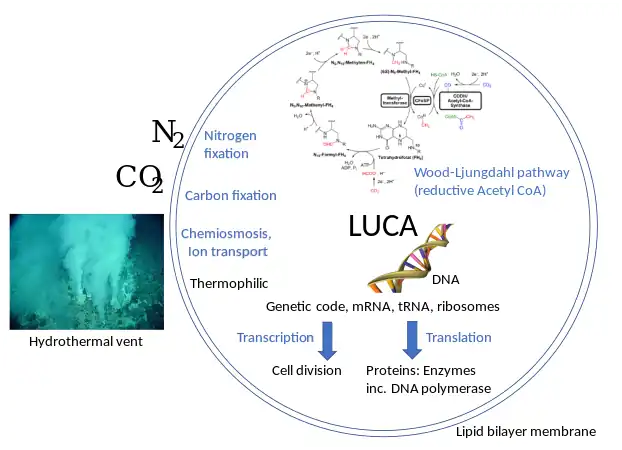
The LUCA certainly had genes and a genetic code.[13] Its genetic material was most likely DNA,[17] so that it lived after the RNA world.[lower-alpha 1][20] The DNA was kept double-stranded by an enzyme, DNA polymerase, which recognises the structure and directionality of DNA.[21] The integrity of the DNA was maintained by a group of repair enzymes including DNA topoisomerase.[22] If the genetic code was based on dual-stranded DNA, it was expressed by copying the information to single-stranded RNA. The RNA was produced by a DNA-dependent RNA polymerase using nucleotides similar to those of DNA.[17] It had multiple DNA-binding proteins, such as histone-fold proteins.[23] The genetic code was expressed into proteins. These were assembled from 20 free amino acids by translation of a messenger RNA via a mechanism of ribosomes, transfer RNAs, and a group of related proteins.[17]
The LUCA was likely capable of sexual interaction in the sense that adaptive gene functions were present that promoted the transfer of DNA between individuals of the population to facilitate genetic recombination. Homologous gene products that promote genetic recombination are present in bacteria, archaea and eukaryota, such as the RecA protein in bacteria, the RadA protein in archaea, and the Rad51 and Dmc1 proteins in eukaryota.[24]
As for the cell's gross structure, it contained a water-based cytoplasm effectively enclosed by a lipid bilayer membrane; it was capable of reproducing by cell division.[17] It tended to exclude sodium and concentrate potassium by means of specific ion transporters (or ion pumps). The cell multiplied by duplicating all its contents followed by cellular division. The cell used chemiosmosis to produce energy. It also reduced CO2 and oxidized H2 (methanogenesis or acetogenesis) via acetyl-thioesters.[25][26]
By phylogenetic bracketing, analysis of the presumed LUCA's offspring groups, LUCA appears to have been a small, single-celled organism. It likely had a ring-shaped coil of DNA floating freely within the cell. Morphologically, it would likely not have stood out within a mixed population of small modern-day bacteria. The originator of the three-domain system, Carl Woese, stated that in its genetic machinery, the LUCA would have been a "simpler, more rudimentary entity than the individual ancestors that spawned the three [domains] (and their descendants)".[1]
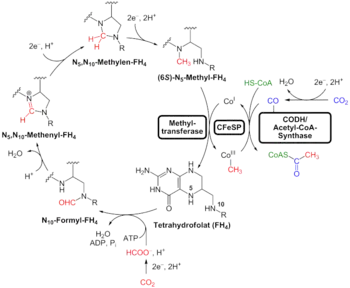
An alternative to the search for "universal" traits is to use genome analysis to identify phylogenetically ancient genes. This gives a picture of a LUCA that could live in a geochemically harsh environment and is like modern prokaryotes. Analysis of biochemical pathways implies the same sort of chemistry as does phylogenetic analysis. Weiss and colleagues write that "Experiments ... demonstrate that ... acetyl-CoA pathway [chemicals used in anaerobic respiration] formate, methanol, acetyl moieties, and even pyruvate arise spontaneously ... from CO2, native metals, and water", a combination present in hydrothermal vents.[13]
An experiment shows that Zn2+, Cr3+, and Fe can promote 6 of the 11 reactions of an ancient anabolic pathway called the reverse Krebs cycle in acidic conditions which implies that LUCA might have inhabited either hydrothermal vents or acidic metal-rich hydrothermal fields.[27]
Because both bacteria and archaea have differences in the structure of phospholipids and cell wall, ion pumping, most proteins involved in DNA replication, and glycolysis, it is inferred that LUCA had a permeable membrane without an ion pump. The emergence of Na+/H+ antiporters likely lead to the evolution of impermeable membranes present in eukaryotes, archaea, and bacteria. It's stated that "The late and independent evolution of glycolysis but not gluconeogenesis is entirely consistent with LUCA being powered by natural proton gradients across leaky membranes. Several discordant traits are likely to be linked to the late evolution of cell membranes, notably the cell wall, whose synthesis depends on the membrane and DNA replication".[28] Although LUCA likely had DNA, it is unknown if it could replicate DNA and is suggested to "might just have been a chemically stable repository for RNA-based replication".[29] It is likely that the permeable membrane of LUCA was composed of archaeal lipids (isoprenoids) and bacterial lipids (fatty acids). Isoprenoids would have enhanced stabilization of LUCA's membrane in the surrounding extreme habitat. Nick Lane and coauthors state that "The advantages and disadvantages of incorporating isoprenoids into cell membranes in different microenvironments may have driven membrane divergence, with the later biosynthesis of phospholipids giving rise to the unique G1P and G3P headgroups of archaea and bacteria respectively. If so, the properties conferred by membrane isoprenoids place the lipid divide as early as the origin of life".[30]
Alternative interpretations
Some other researchers have challenged Weiss et al.'s 2016 conclusions. Sarah Berkemer and Shawn McGlynn argue that Weiss et al. undersampled the families of proteins, so that the phylogenetic trees were not complete and failed to describe the evolution of proteins correctly.[31] A phylogenomic and geochemical analysis of a set of proteins that probably traced to the LUCA show that it had K+-dependent GTPases and the ionic composition and concentration of its intracellular fluid was seemingly high K+/Na+ ratio, NH+
4, Fe2+, CO2+, Ni2+, Mg2+, Mn2+, Zn2+, pyrophosphate, and PO3−
4 which would imply a terrestrial hot spring habitat. It possibly had a phosphate-based metabolism. Further, these proteins were unrelated to autotrophy (the ability of an organism to create its own organic matter), suggesting that the LUCA had a heterotrophic lifestyle (consuming organic matter) and that its growth was dependent on organic matter produced by the physical environment.[32] Nick Lane argues that Na+/H+ antiporters could readily explain the low concentration of Na+ in the LUCA and its descendants.[28] The emergence of LUCA in a terrestrial habitat is stated to not be possible to have occurred due to the ultraviolet irradiation and the Late Heavy Bombardment at the early Earth.[33]
The identification of thermophilic genes in the LUCA has also been criticized, as they may instead represent genes that evolved later in archaea or bacteria, then migrated between these via horizontal gene transfers, as in Woese's 1998 hypothesis.[34] LUCA could have been a mesophile that fixed CO2 and relied on H2, and lived close to hydrothermal vents.[35] Further, the presence of the energy-handling enzymes CODH/acetyl-coenzyme A synthase in LUCA could be compatible not only with being an autotroph but also with life as a mixotroph or heterotroph.[36] Weiss et al. 2018 reply that no enzyme defines a trophic lifestyle, and that heterotrophs evolved from autotrophs.[29]
Age
Studies from 2000 to 2018 have suggested an increasingly ancient time for the LUCA. In 2000, estimates of the LUCA's age ranged from 3.5 to 3.8 billion years ago in the Paleoarchean,[37] a few hundred million years before the earliest fossil evidence of life, for which candidates range in age from 3.48 to 4.28 billion years ago.[38][39][40][41][42] This placed the origin of the first forms of life shortly after the Late Heavy Asteroid Bombardment which was thought to have repeatedly sterilized Earth's surface. However, a 2018 study from the University of Bristol, applying a molecular clock model, found that the data (102 species, 29 common protein-coding genes, mostly ribosomal) is compatible only with first life forms as early as within 0.001–0.043 billion years of the maximum possible age given by the Earth-sterilizing Moon-forming event about 4.5 billion years ago.[43]
Root of the tree of life
The most commonly accepted tree of life, based on several molecular studies, has its root between a monophyletic domain Bacteria and a clade formed by Archaea and Eukaryota.[45] A small minority of studies place the root in the domain bacteria, in the phylum Bacillota,[46] or state that the phylum Chloroflexota (formerly Chloroflexi) is basal to a clade with Archaea and Eukaryotes and the rest of bacteria (as proposed by Thomas Cavalier-Smith).[47] Recent genomic analyses recover a two-domain system with the domains Archaea and Bacteria; in this view of the tree of life, Eukaryotes are derived from Archaea.[48][49][50]
With the later gene pool of the LUCA's descendants, sharing a common framework of the AT/GC rule and the standard twenty amino acids, horizontal gene transfer would have become feasible and could have been common.
The nature of LUCA remains disputed. In 1994, on the basis of primordial metabolism (sensu Wächtershäuser), Otto Kandler proposed a successive divergence of the three domains of life[1] from a multiphenotypical population of pre-cells, reached by evolutionary improvements (cellularization).[51][52][53] These phenotypically diverse pre-cells were metabolising, self-reproducing entities exhibiting frequent mutual exchange of genetic information. In this scenario there was no "first cell". Kandler's pre-cell theory is supported by Wächtershäuser.[54][55] In 1998, Carl Woese, based on the RNA world concept, proposed that no individual organism could be considered a LUCA, and that the genetic heritage of all modern organisms derived through horizontal gene transfer among an ancient community of organisms.[56] Other authors concur that there was a "complex collective genome"[57] at the time of the LUCA, and that horizontal gene transfer was important in the evolution of later groups;[57] Nicolas Glansdorff states that LUCA "was in a metabolically and morphologically heterogeneous community, constantly shuffling around genetic material" and "remained an evolutionary entity, though loosely defined and constantly changing, as long as this promiscuity lasted."[58]
While the nature of the LUCA remains a subject of research,[4] the theory of a universal common ancestry of life is, of course, widely accepted. In 2010, based on "the vast array of molecular sequences now available from all domains of life,"[59] D. L. Theobald published a "formal test" of universal common ancestry (UCA).[5] This theory deals with the common descent of all extant terrestrial organisms, each being a genealogical descendant of a single species from the distant past. His formal test favored the existence of a universal common ancestry over a wide class of alternative hypotheses that included horizontal gene transfer. Basic biochemical principles make it likely that all organisms do have a common ancestry.
LUCA and viruses
The origin of viruses remains disputed. Since viruses need host cells for their replication, it is likely that they emerged after the formation of cells. Viruses may even have multiple origins and different types of viruses may have evolved independently over the history of life.[3]
There are different hypotheses for the origins of viruses, for instance an early viral origin from the RNA world or a later viral origin from selfish DNA.[3]
Based on how viruses are currently distributed across the bacteria and archaea, the LUCA may well have been prey to multiple viruses, ancestral to those that now have those two domains as their hosts.[60] Furthermore, extensive virus evolution seems to have preceded the LUCA, since the jelly-roll structure of capsid proteins is shared by RNA and DNA viruses across all three domains of life.[61][62] LUCA's viruses were probably mainly dsDNA viruses in the groups called Duplodnaviria and Varidnaviria. Two other single-stranded DNA virus groups within the Monodnaviria, the Microviridae and the Tubulavirales, likely infected the last bacterial common ancestor. The last archaeal common ancestor was probably host to spindle-shaped viruses. All of these could well have affected the LUCA, in which case each must since have been lost in the host domain where it is no longer extant. By contrast, RNA viruses do not appear to have been important parasites of LUCA, even though straightforward thinking might have envisaged viruses as beginning with RNA viruses directly derived from an RNA world. Instead, by the time the LUCA lived, RNA viruses had probably already been out-competed by DNA viruses.[60]
See also
- Abiogenesis – Natural process by which life arises from non-living matter
- Cellularization – Scientific theory to explain the origin and formation of cells
- Chemoton – Abstract model for the fundamental unit of life
- Darwinian threshold – Transition period during the evolution of the first cells
- Last eukaryotic common ancestor – the last common ancestor of animals, plants, and fungi
- Mitochondrial Eve – Matrilineal most recent common ancestor of all living humans
- Pre-cell – Hypothetical life before complete cells
- Timeline of the evolutionary history of life – Major events during the development of life
- Urmetazoan – Hypothetical last common ancestor of all animals
- Y-chromosomal Adam – Patrilineal most recent common ancestor of all living humans
Notes
References
- Woese, C.R.; Kandler, O.; Wheelis, M.L. (June 1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". PNAS. 87 (12): 4576–4579. Bibcode:1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC 54159. PMID 2112744.
- Madigan, Michael T.; Martinko, John M.; Bender, Kelly S.; Buckley, Daniel H.; Stahl, David A. (2015). Brock Biology of Microorganisms (14 ed.). Boston: Pearson Education Limited. pp. 29, 374, 381. ISBN 978-1-292-01831-7.
- Madigan, Michael T.; Aiyer, Jennifer; Buckley, Daniel H.; Sattley, Matthew; Stahl, David A. (2022). Brock Biology of Microorganisms (16 ed.). Harlow: Pearson Education Limited. pp. Unit 3, chapter 13: 431 (LUCA), 428, 438, 439 (viruses). ISBN 978-1-292-40479-0.
- Harold, Franklin M. (2014). In Search of Cell History: The Evolution of Life's Building Blocks. Chicago, London: University of Chicago Press. ISBN 978-0-226-17428-0.
- Theobald, D. L. (May 2010). "A formal test of the theory of universal common ancestry". Nature. 465 (7295): 219–222. Bibcode:2010Natur.465..219T. doi:10.1038/nature09014. PMID 20463738. S2CID 4422345.
- Gregory, T. Ryan (2008). "Understanding evolutionary trees". Evolution: Education and Outreach. 1 (2): 121–137. doi:10.1007/s12052-008-0035-x. S2CID 15488906.
- Lamarck, Jean Baptiste Pierre Antoine de Monet de (1994) [1809]. Philosophie zoologique (PDF). Paris. p. 737.
{{cite book}}: CS1 maint: location missing publisher (link) - Noble, Denis (1 July 2020). "Editorial: Charles Darwin, Jean-Baptiste Lamarck, and 21st century arguments on the fundamentals of biology". Progress in Biophysics and Molecular Biology. 153: 1–4. doi:10.1016/j.pbiomolbio.2020.02.005. PMID 32092299. S2CID 211475380. Retrieved 23 December 2022.
- Darwin, Charles (1859). The Origin of Species by Means of Natural Selection. John Murray. pp. 484, 490.
- Wikham, Gene Stephen (March 1995). The molecular phylogenetic analysis of naturally occurring hyperthermophilic microbial communities (PhD thesis). Indiana University. p. 4. ProQuest 304192982
- Forterre, Patrick (1997). "Archaea: What can we learn from their sequences?". Current Opinion in Genetics & Development. 7 (6): 764–770. doi:10.1016/s0959-437x(97)80038-x. PMID 9468785.
- Koonin, Eugene V.; Galperin, Michael Y. (2003). Sequence - Evolution - Function: Computational approaches in comparative genomics. Boston, MA: Kluwer. p. 252. ISBN 978-1-4757-3783-7. OCLC 55642057.
- Weiss, M. C.; Preiner, M.; Xavier, J. C.; et al. (2018). "The last universal common ancestor between ancient Earth chemistry and the onset of genetics". PLOS Genetics. 14 (8): e1007518. doi:10.1371/journal.pgen.1007518. PMC 6095482. PMID 30114187.
- Weiss, Madeline C.; Sousa, F. L.; Mrnjavac, N.; et al. (2016). "The physiology and habitat of the last universal common ancestor" (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/nmicrobiol.2016.116. PMID 27562259. S2CID 2997255.
- Harrison, Stuart A.; Palmeira, Raquel Nunes; Halpern, Aaron; Lane, Nick (1 November 2022). "A biophysical basis for the emergence of the genetic code in protocells". Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1863 (8): 148597. doi:10.1016/j.bbabio.2022.148597. ISSN 0005-2728. PMID 35868450.
- Harrison, Stuart A.; Lane, Nick (12 December 2018). "Life as a guide to prebiotic nucleotide synthesis". Nature Communications. 9 (1): 5176. Bibcode:2018NatCo...9.5176H. doi:10.1038/s41467-018-07220-y. ISSN 2041-1723. PMC 6289992. PMID 30538225.
- Wächtershäuser, Günter (1998). "Towards a Reconstruction of Ancestral Genomes by Gene Cluster Alignment". Systematic and Applied Microbiology. 21 (4): 473–474, IN1, 475–477. doi:10.1016/S0723-2020(98)80058-1.
- Marshall, Michael. "Life began with a planetary mega-organism". New Scientist. Archived from the original on 25 July 2016. Retrieved 31 July 2016.
- Koonin, Eugene V.; Martin, William (1 December 2005). "On the origin of genomes and cells within inorganic compartments". Trends in Genetics. 21 (12): 647–654. doi:10.1016/j.tig.2005.09.006. PMC 7172762. PMID 16223546.
- Garwood, Russell J. (2012). "Patterns In Palaeontology: The first 3 billion years of evolution". Palaeontology Online. 2 (11): 1–14. Archived from the original on 26 June 2015. Retrieved 25 June 2015.
- Koonin, Eugene V.; Krupovic, M.; Ishino, S.; Ishino, Y. (2020). "The replication machinery of LUCA: common origin of DNA replication and transcription". BMC Biology. 18 (1): 61. doi:10.1186/s12915-020-00800-9. PMC 7281927. PMID 32517760.
- Ahmad, Muzammil; Xu, Dongyi; Wang, Weidong (23 May 2017). "Type IA topoisomerases can be "magicians" for both DNA and RNA in all domains of life". RNA Biology. 14 (7): 854–864. doi:10.1080/15476286.2017.1330741. PMC 5546716. PMID 28534707.
- Lupas, Andrei N.; Alva, Vikram (2018). "Histones predate the split between bacteria and archaea". Bioinformatics. 35 (14): 2349–2353. doi:10.1093/bioinformatics/bty1000. PMID 30520969.
- Bernstein, H., Bernstein, C. (2017). Sexual Communication in Archaea, the Precursor to Eukaryotic Meiosis. In: Witzany, G. (eds) Biocommunication of Archaea. Springer, Cham. https://doi.org/10.1007/978-3-319-65536-9_7
- Martin, W.; Russell, M. J. (October 2007). "On the origin of biochemistry at an alkaline hydrothermal vent". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 362 (1486): 1887–1925. doi:10.1098/rstb.2006.1881. PMC 2442388. PMID 17255002.
- Lane, Nick; Allen, J. F.; Martin, W. (April 2010). "How did LUCA make a living? Chemiosmosis in the origin of life". BioEssays. 32 (4): 271–280. doi:10.1002/bies.200900131. PMID 20108228.
- Muchowska, Kamila B.; Varma, Sreejith J.; Chevallot-Beroux, Elodie; Lethuillier-Karl, Lucas; Li, Guang; Moran, Joseph (2 October 2017). "Metals promote sequences of the reverse Krebs cycle". Nature Ecology & Evolution. 1 (11): 1716–1721. doi:10.1038/s41559-017-0311-7. ISSN 2397-334X. PMC 5659384. PMID 28970480.
- Sojo, Víctor; Pomiankowski, Andrew; Lane, Nick (12 August 2014). "A Bioenergetic Basis for Membrane Divergence in Archaea and Bacteria". PLOS Biology. 12 (8): e1001926. doi:10.1371/journal.pbio.1001926. ISSN 1545-7885. PMC 4130499. PMID 25116890.
- Weiss, Madeline C.; Preiner, Martina; Xavier, Joana C.; Zimorski, Verena; Martin, William F. (16 August 2018). "The last universal common ancestor between ancient Earth chemistry and the onset of genetics". PLOS Genetics. 14 (8): e1007518. doi:10.1371/journal.pgen.1007518. PMC 6095482. PMID 30114187. S2CID 52019935.
- Jordan, S. F.; Nee, E.; Lane, N. (18 October 2019). "Isoprenoids enhance the stability of fatty acid membranes at the emergence of life potentially leading to an early lipid divide". Interface Focus. 9 (6). doi:10.1098/rsfs.2019.0067. ISSN 2042-8901. PMC 6802135. PMID 31641436.
 This article incorporates text from this source, which is available under the CC BY 4.0 license.
This article incorporates text from this source, which is available under the CC BY 4.0 license.
- Berkemer, Sarah J.; McGlynn, Shawn E. (2020). "A New Analysis of Archaea–Bacteria Domain Separation: Variable Phylogenetic Distance and the Tempo of Early Evolution". Molecular Biology and Evolution. 37 (8): 2332–2340. doi:10.1093/molbev/msaa089. PMC 7403611. PMID 32316034.
- Mulkidjanian, Armen Y.; Bychkov, Andrew Yu.; Dibrova, Daria V.; Galperin, Michael Y.; Koonin, Eugene V. (2012). "Origin of first cells at terrestrial, anoxic geothermal fields". Proceedings of the National Academy of Sciences. 109 (14): E821-30. Bibcode:2012PNAS..109E.821M. doi:10.1073/pnas.1117774109. PMC 3325685. PMID 22331915.
- Lane, Nick; Allen, John F.; Martin, William (27 January 2010). "How did LUCA make a living? Chemiosmosis in the origin of life". BioEssays. 32 (4): 271–280. doi:10.1002/bies.200900131. PMID 20108228.
- Gogarten, Johann Peter; Deamer, David (2016). "Is LUCA a thermophilic progenote?". Nature Microbiology. 1 (12): 16229. doi:10.1038/nmicrobiol.2016.229. PMID 27886195. S2CID 205428194.
- Camprubí, E.; de Leeuw, J. W.; House, C. H.; Raulin, F.; Russell, M. J.; Spang, A.; Tirumalai, M. R.; Westall, F. (12 December 2019). "The Emergence of Life". Space Science Reviews. 215 (8): 56. Bibcode:2019SSRv..215...56C. doi:10.1007/s11214-019-0624-8. ISSN 1572-9672.
- Adam, Panagiotis S.; Borrel, Guillaume; Gribaldo, Simonetta (6 February 2018). "Evolutionary history of carbon monoxide dehydrogenase/acetyl-CoA synthase, one of the oldest enzymatic complexes". PNAS. 115 (6): E1166–E1173. Bibcode:2018PNAS..115E1166A. doi:10.1073/pnas.1716667115. PMC 5819426. PMID 29358391.
- Doolittle, W. F. (February 2000). "Uprooting the tree of life". Scientific American. 282 (2): 90–95. Bibcode:2000SciAm.282b..90D. doi:10.1038/scientificamerican0200-90. JSTOR 26058605. PMID 10710791.
- Noffke, N.; Christian, D.; Wacey, D.; Hazen, R. M. (December 2013). "Microbially induced sedimentary structures recording an ancient ecosystem in the ca. 3.48 billion-year-old Dresser Formation, Pilbara, Western Australia". Astrobiology. 13 (12): 1103–1124. Bibcode:2013AsBio..13.1103N. doi:10.1089/ast.2013.1030. PMC 3870916. PMID 24205812.
- Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T. (2013). "Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks". Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe...7...25O. doi:10.1038/ngeo2025.
- Hassenkam, T.; Andersson, M. P.; Dalby, K. N.; et al. (2017). "Elements of Eoarchean life trapped in mineral inclusions". Nature. 548 (7665): 78–81. Bibcode:2017Natur.548...78H. doi:10.1038/nature23261. PMID 28738409. S2CID 205257931.
- Bell, Elizabeth A.; Boehnke, Patrick; Harrison, T. Mark; Mao, Wendy L. (24 November 2015). "Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon". PNAS. 112 (47): 14518–14521. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481.
- Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; et al. (2 March 2017). "Evidence for early life in Earth's oldest hydrothermal vent precipitates" (PDF). Nature. 543 (7643): 60–64. Bibcode:2017Natur.543...60D. doi:10.1038/nature21377. PMID 28252057. S2CID 2420384. Archived (PDF) from the original on 23 July 2018. Retrieved 25 June 2019.
- Betts, Holly C.; Puttick, Mark N.; Clark, James W.; Williams, Tom A.; Donoghue, Philip C. J.; Pisani, Davide (2018). "Integrated genomic and fossil evidence illuminates life's early evolution and eukaryote origin". Nature Ecology & Evolution. 2 (10): 1556–1562. doi:10.1038/s41559-018-0644-x. PMC 6152910. PMID 30127539.
- Smets, Barth F.; Barkay, Tamar (September 2005). "Horizontal gene transfer: perspectives at a crossroads of scientific disciplines". Nature Reviews Microbiology. 3 (9): 675–678. doi:10.1038/nrmicro1253. PMID 16145755. S2CID 2265315.
- Brown, J. R.; Doolittle, W. F. (1995). "Root of the Universal Tree of Life Based on Ancient Aminoacyl-tRNA Synthetase Gene Duplications". PNAS. 92 (7): 2441–2445. Bibcode:1995PNAS...92.2441B. doi:10.1073/pnas.92.7.2441. PMC 42233. PMID 7708661.
- Valas, R. E.; Bourne, P. E. (2011). "The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon". Biology Direct. 6: 16. doi:10.1186/1745-6150-6-16. PMC 3056875. PMID 21356104.
- Cavalier-Smith, Tom (2006). "Rooting the tree of life by transition analyses". Biology Direct. 1: 19. doi:10.1186/1745-6150-1-19. PMC 1586193. PMID 16834776.
- Raymann, Kasie; Brochier-Armanet, Céline; Gribaldo, Simonetta (26 May 2015). "The two-domain tree of life is linked to a new root for the Archaea". Proceedings of the National Academy of Sciences. 112 (21): 6670–6675. Bibcode:2015PNAS..112.6670R. doi:10.1073/pnas.1420858112. ISSN 0027-8424. PMC 4450401. PMID 25964353.
- Hug, Laura A.; Baker, Brett J.; Anantharaman, Karthik; Brown, Christopher T.; Probst, Alexander J.; Castelle, Cindy J.; Butterfield, Cristina N.; Hernsdorf, Alex W.; Amano, Yuki; Ise, Kotaro; Suzuki, Yohey; Dudek, Natasha; Relman, David A.; Finstad, Kari M.; Amundson, Ronald (11 April 2016). "A new view of the tree of life". Nature Microbiology. 1 (5): 16048. doi:10.1038/nmicrobiol.2016.48. ISSN 2058-5276. PMID 27572647. S2CID 3833474.
- Williams, Tom A.; Foster, Peter G.; Cox, Cymon J.; Embley, T. Martin (11 December 2013). "An archaeal origin of eukaryotes supports only two primary domains of life". Nature. 504 (7479): 231–236. Bibcode:2013Natur.504..231W. doi:10.1038/nature12779. ISSN 1476-4687. PMID 24336283. S2CID 4461775.
- Kandler, Otto (1994). "The early diversification of life". In Stefan Bengtson (ed.). Early Life on Earth. Nobel Symposium 84. New York: Columbia U.P. pp. 152–160.
- Kandler, Otto (1995). "Cell Wall Biochemistry in Archaea and its Phylogenetic Implications". Journal of Biological Physics. 20 (1–4): 165–169. doi:10.1007/BF00700433. S2CID 83906865.
- Kandler, Otto (1998). "The early diversification of life and the origin of the three domains: A proposal". In Jürgen Wiegel; Michael W.W. Adams (eds.). Thermophiles: The keys to molecular evolution and the origin of life?. London: Taylor and Francis Ltd. pp. 19–31. ISBN 978-0-203-48420-3.
- Wächtershäuser, Günter (2003). "From pre-cells to Eukarya – a tale of two lipids". Molecular Microbiology. 47 (1): 13–22. doi:10.1046/j.1365-2958.2003.03267.x. PMID 12492850. S2CID 37944519.
- Wächtershäuser, Günter (October 2006). "From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 361 (1474): 1787–1808. doi:10.1098/rstb.2006.1904. PMC 1664677. PMID 17008219.
- Woese, Carl (June 1998). "The universal ancestor". PNAS. 95 (12): 6854–6859. Bibcode:1998PNAS...95.6854W. doi:10.1073/pnas.95.12.6854. PMC 22660. PMID 9618502.
- Egel, Richard (March 2012). "Primal Eukaryogenesis: On the Communal Nature of Precellular States, Ancestral to Modern Life". Life. 2 (1): 170–212. Bibcode:2012Life....2..170E. doi:10.3390/life2010170. PMC 4187143. PMID 25382122.
- Glansdorff, Nicolas; Xu, Ying; Labedan, Bernard (9 July 2008). "The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner". Biology Direct. 3 (1): 29. doi:10.1186/1745-6150-3-29. PMC 2478661. PMID 18613974. S2CID 18250196.
- Steel, M.; Penny, D. (May 2010). "Origins of life: Common ancestry put to the test". Nature. 465 (7295): 168–169. Bibcode:2010Natur.465..168S. doi:10.1038/465168a. PMID 20463725. S2CID 205055573.
- Krupovic, M.; Dolja, V. V.; Koonin, Eugene V. (2020). "The LUCA and its complex virome" (PDF). Nature Reviews Microbiology. 18 (11): 661–670. doi:10.1038/s41579-020-0408-x. PMID 32665595. S2CID 220516514.
- Forterre, Patrick; Prangishvili, David (2009). "The origin of viruses". Research in Microbiology. 160 (7): 466–472. doi:10.1016/j.resmic.2009.07.008. PMID 19647075. S2CID 2767388.
- Durzyńska, Julia; Goździcka-Józefiak, Anna (16 October 2015). "Viruses and cells intertwined since the dawn of evolution". Virology Journal. 12 (1): 169. doi:10.1186/s12985-015-0400-7. PMC 4609113. PMID 26475454.
Further reading
- Lane, Nick (2016) [2015]. The Vital Question. London: Profile Books. ISBN 978-1781250372.