Magnetoreception
Magnetoreception is a sense which allows an organism to detect the Earth's magnetic field. Animals with this sense include some arthropods, molluscs, and vertebrates (fish, amphibians, reptiles, birds, and mammals). The sense is mainly used for orientation and navigation, but it may help some animals to form regional maps. Experiments on migratory birds provide evidence that they make use of a cryptochrome protein in the eye, relying on the quantum radical pair mechanism to perceive magnetic fields. This effect is extremely sensitive to weak magnetic fields, and readily disturbed by radio-frequency interference, unlike a conventional iron compass.

Birds have iron-containing materials in their upper beaks. There is some evidence that this provides a magnetic sense, mediated by the trigeminal nerve, but the mechanism is unknown.
Cartilaginous fish including sharks and stingrays can detect small variations in electric potential with their electroreceptive organs, the ampullae of Lorenzini. These appear to be able to detect magnetic fields by induction. There is some evidence that these fish use magnetic fields in navigation.
History
Biologists have long wondered whether migrating animals such as birds and sea turtles have an inbuilt magnetic compass, enabling them to navigate using the Earth's magnetic field. Until late in the 20th century, evidence for this was essentially only behavioural: many experiments demonstrated that animals could indeed derive information from the magnetic field around them, but gave no indication of the mechanism. In 1972, Roswitha and Wolfgang Wiltschko showed that migratory birds responded to the direction and inclination (dip) of the magnetic field. In 1977, M. M. Walker and colleagues identified iron-based (magnetite) magnetoreceptors in the snouts of rainbow trout. In 2003, G. Fleissner and colleagues found iron-based receptors in the upper beaks of homing pigeons, both seemingly connected to the animal's trigeminal nerve. Research took a different direction in 2000, however, when Thorsten Ritz and colleagues suggested that a photoreceptor protein in the eye, cryptochrome, was a magnetoreceptor, working at a molecular scale by quantum entanglement.[1]
Proposed mechanisms
In animals
In animals, the mechanism for magnetoreception is still under investigation. Two main hypotheses are currently being discussed: one proposing a quantum compass based on a radical pair mechanism,[2] the other postulating a more conventional iron-based magnetic compass with magnetite particles.[3]
Cryptochrome
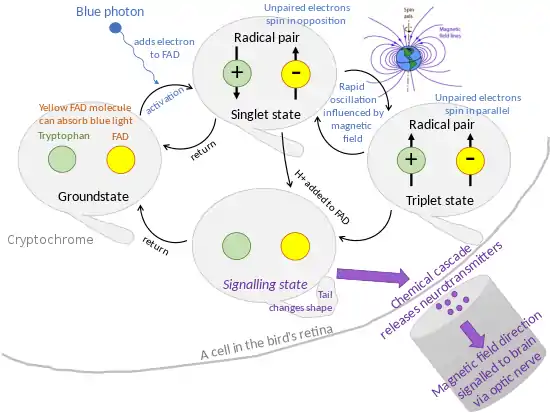
According to the first model, magnetoreception is possible via the radical pair mechanism,[5] which is well-established in spin chemistry. The mechanism requires two molecules, each with unpaired electrons, at a suitable distance from each other. When these can exist in states either with their spin axes in the same direction, or in opposite directions, the molecules oscillate rapidly between the two states. That oscillation is extremely sensitive to magnetic fields.[6][7][8][9] Because the Earth's magnetic field is extremely weak, at 0.5 gauss, the radical pair mechanism is currently the only credible way that the Earth's magnetic field could cause chemical changes.[9]
In 1978, Schulten and colleagues proposed that this was the mechanism of magnetoreception.[10] In 2000, scientists proposed that cryptochrome – a flavoprotein in the rod cells in the eyes of birds – was the "magnetic molecule" behind this effect.[11] It is the only protein known to form photoinduced radical-pairs in animals.[5] The function of cryptochrome varies by species, but its mechanism is always the same: exposure to blue light excites an electron in a chromophore, which causes the formation of a radical-pair whose electrons are quantum entangled, enabling the precision needed for magnetoreception.[12][13]
Many lines of evidence point to cryptochrome and radical pairs as the mechanism of magnetoreception in birds:[4]
- Despite 20 years of searching, no biomolecule other than cryptochrome has been identified capable of supporting radical pairs.[4]
- In cryptochrome, a yellow molecule flavin adenine dinucleotide (FAD) can absorb a photon of blue light, putting the cryptochrome into an activated state: an electron is transferred from a tryptophan amino acid to the FAD molecule, forming a radical pair.[4]
- Of the six types of cryptochrome in birds, cryptochrome-4a (Cry4a) binds FAD much more tightly than the rest.[4]
- Cry4a levels in migratory birds, which rely on navigation for their survival, are highest during the spring and autumn migration periods, when navigation is most critical.[4]
- The Cry4a protein from the European robin, a migratory bird, is much more sensitive to magnetic fields than similar but not identical Cry4a from pigeons and chickens, which are non-migratory.[4]
These findings together suggest that the Cry4a of migratory birds has been selected for its magnetic sensitivity.[4]
Behavioral experiments on migratory birds also support this theory. Caged migratory birds such as robins display migratory restlessness, known by ethologists as Zugunruhe, in spring and autumn: they often orient themselves in the direction in which they would migrate. In 2004, Thorsten Ritz showed that a weak radio-frequency electromagnetic field, chosen to be at the same frequency as the singlet-triplet oscillation of cryptochrome radical pairs, effectively interfered with the birds' orientation. The field would not have interfered with an iron-based compass. Further, birds are unable to detect a 180 degree reversal of the magnetic field, something they would straightforwardly detect with an iron-based compass.[4]
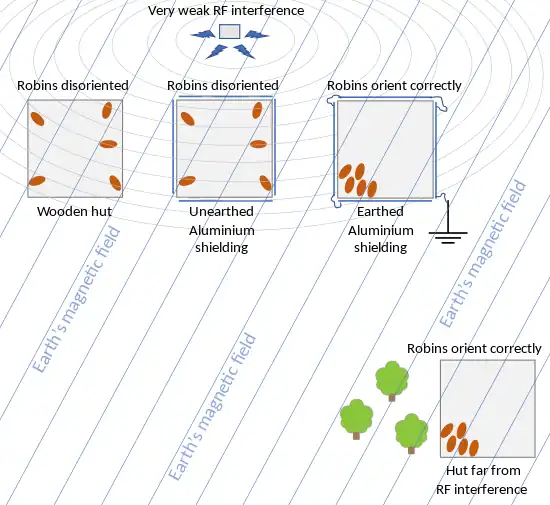
From 2007 onwards, Henrik Mouritsen attempted to replicate this experiment. Instead, he found that robins were unable to orient themselves in the wooden huts he used. Suspecting extremely weak radio-frequency interference from other electrical equipment on the campus, he tried shielding the huts with aluminium sheeting, which blocks electrical noise but not magnetic fields. When he earthed the sheeting, the robins oriented correctly; when the earthing was removed, the robins oriented at random. Finally, when the robins were tested in a hut far from electrical equipment, the birds oriented correctly. These effects imply a radical-pair compass, not an iron one.[4]
In 2016, Wiltschko and colleagues showed that cryptochrome can be activated in the dark, removing the objection that the blue light-activated mechanism would not work when birds were migrating at night. A different radical pair is formed by re-oxidation of fully-reduced FADH−. Experiments with European robins, using flickering light and a magnetic field switched off when the light was on, showed that the birds detected the field without light. The birds were unaffected by local anaesthesia of the upper beak, showing that in these test conditions orientation was not from iron-based receptors in the beak. In their view, cryptochrome and its radical pairs provide the only model that can explain the avian magnetic compass.[12] A scheme with three radicals rather than two has been proposed as more resistant to spin relaxation and explaining the observed behaviour better.[14]
Iron-based
The second proposed model for magnetoreception relies on clusters composed of iron, a natural mineral with strong magnetism, used by magnetotactic bacteria. Iron clusters have been observed in the upper beak of homing pigeons,[15] and other taxa.[16][5][17][18] Iron-based systems could form a magnetoreceptive basis for many species including turtles.[9] Both the exact location and ultrastructure of birds' iron-containing magnetoreceptors remain unknown; they are believed to be in the upper beak, and to be connected to the brain by the trigeminal nerve. This system is in addition to the cryptochrome system in the retina of birds. Iron-based systems of unknown function might also exist in other vertebrates.[19]
Electromagnetic induction

Another possible mechanism of magnetoreception in animals is electromagnetic induction in cartilaginous fish, namely sharks, stingrays, and chimaeras. These fish have electroreceptive organs, the ampullae of Lorenzini, which can detect small variations in electric potential. The organs are mucus-filled and consist of canals that connect pores in the skin of the mouth and nose to small sacs within the animal's flesh. They are used to sense the weak electric fields of prey and predators. These organs have been predicted to sense magnetic fields, by means of Faraday's law of induction: as a conductor moves through a magnetic field an electric potential is generated. In this case the conductor is the animal moving through a magnetic field, and the potential induced (Vind) depends on the time (t)-varying rate of magnetic flux (Φ) through the conductor according to

The ampullae of Lorenzini detect very small fluctuations in the potential difference between the pore and the base of the electroreceptor sac. An increase in potential results in a decrease in the rate of nerve activity. This is analogous to the behavior of a current-carrying conductor.[21][22][23] Sandbar sharks, Carcharinus plumbeus, have been shown to be able to detect magnetic fields; the experiments provided non-definitive evidence that the animals had a magnetoreceptor, rather than relying on induction and electroreceptors.[23] Electromagnetic induction has not been studied in non-aquatic animals.[9]
The yellow stingray, Urobatis jamaicensis, is able to distinguish between the intensity and inclination angle of a magnetic field in the laboratory. This suggests that cartilaginous fishes may use the Earth's magnetic field for navigation.[20]
Passive alignment in bacteria
![{\displaystyle \scriptstyle [1{\overline {1}}0]}](../I/f99ce4c507bedc2d8c9e600715fe15e2d2666907.svg)
Magnetotactic bacteria of multiple taxa contain sufficient magnetic material in the form of magnetosomes, nanometer-sized particles of magnetite,[25] that the Earth's magnetic field passively aligns them, just as it does with a compass needle. The bacteria are thus not actually sensing the magnetic field.[26][27]
A possible but unexplored mechanism of magnetoreception in animals is through endosymbiosis with magnetotactic bacteria, whose DNA is widespread in animals. This would involve having these bacteria living inside an animal, and their magnetic alignment being used as part of a magnetoreceptive system.[28]
Unanswered questions
It remains likely that two or more complementary mechanisms play a role in magnetic field detection in animals. Of course, this potential dual mechanism theory raises the questions of to what degree each method is responsible for the stimulus, and how they produce a signal in response to the weak magnetic field of the Earth.[9]
In addition, it is possible that magnetic senses may be different for different species. Some species may only be able to detect north and south, while others may only be able to differentiate between the equator and the poles. Although the ability to sense direction is important in migratory navigation, many animals have the ability to sense small fluctuations in earth's magnetic field to map their position to within a few kilometers.[9][29]
Taxonomic range
Magnetoreception is widely-distributed taxonomically. It is present in many of the animals so far investigated. These include arthropods, molluscs, and among vertebrates in fish, amphibians, reptiles, birds, and mammals. Its status in other groups remains unknown.[30]
The ability to detect and respond to magnetic fields may exist in plants, possibly as in animals mediated by cryptochrome. Experiments by different scientists have identified multiple effects, including changes to growth rate, seed germination, mitochondrial structure, and responses to gravity (geotropism). The results have sometimes been controversial, and no mechanism has been definitely identified. The ability may be widely distributed, but its taxonomic range in plants is unknown.[31]
| Eukaryotes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| cryptochrome |
_(cropped).jpg.webp)


In molluscs
The giant sea slug Tochuina gigantea (formerly T. tetraquetra), a mollusc, orients its body between north and east prior to a full moon.[32] A 1991 experiment offered a right turn to geomagnetic south and a left turn to geomagnetic east (a Y-shaped maze). 80% of Tochuina made a turn to magnetic east. When the field was reversed, the animals displayed no preference for either turn.[33][34] Tochuina's nervous system is composed of individually identifiable neurons, four of which are stimulated by changes in the applied magnetic field, and two which are inhibited by such changes.[34] The tracks of the similar species Tritonia exsulans become more variable in direction when close to strong rare-earth magnets placed in their natural habitat, suggesting that the animal uses its magnetic sense continuously to help it travel in a straight line.[35]
In insects
The fruit fly Drosophila melanogaster may be able to orient to magnetic fields. In one choice test, flies were loaded into an apparatus with two arms that were surrounded by electric coils. Current was run through each of the coils, but only one was configured to produce a 5-Gauss magnetic field (about ten times stronger than the Earth's magnetic field) at a time. The flies were trained to associate the magnetic field with a sucrose reward. Flies with an altered cryptochrome, such as with an antisense mutation, were not sensitive to magnetic fields.[36]
Magnetoreception has been studied in detail in insects including honey bees, ants and termites.[37] Ants and bees navigate using their magnetic sense both locally (near their nests) and when migrating.[38] In particular, the Brazilian stingless bee Schwarziana quadripunctata is able to detect magnetic fields using the thousands of hair-like sensilla on its antennae.[39][40]
In fish
Studies of magnetoreception in bony fish have been conducted mainly with salmon. Both sockeye salmon (Oncorhynchus nerka) and Chinook salmon (Oncorhynchus tschawytscha) have a compass sense. This was demonstrated in experiments in the 1980s by changing the axis of a magnetic field around a circular tank of young fish; they reoriented themselves in line with the field.[41][42]
In amphibians
Some of the earliest studies of amphibian magnetoreception were conducted with cave salamanders (Eurycea lucifuga). Researchers housed groups of cave salamanders in corridors aligned with either magnetic north–south, or magnetic east–west. In tests, the magnetic field was experimentally rotated by 90°, and salamanders were placed in cross-shaped structures (one corridor along the new north–south axis, one along the new east–west axis). The salamanders responded to the field's rotation.[43]
Red-spotted newts (Notophthalmus viridescens) respond to drastic increases in water temperature by heading for land. The behaviour is disrupted if the magnetic field is experimentally altered, showing that the newts use the field for orientation.[44][45]
Both European toads (Bufo bufo) and natterjack toads (Epidalea calamita) toads rely on vision and olfaction when migrating to breeding sites, but magnetic fields may also play a role. When randomly displaced 150 metres (490 ft) from their breeding sites, these toads can navigate their way back,[46] but this ability can be disrupted by fitting them with small magnets.[47]
In reptiles

The majority of study on magnetoreception in reptiles involves turtles. Early support for magnetoreception in turtles was provided in a 1991 study on hatchling loggerhead turtles which demonstrated that loggerheads can use the magnetic field as a compass to determine direction.[48] Subsequent studies have demonstrated that loggerhead and green turtles can also use the magnetic field of the earth as a map, because different parameters of the Earth's magnetic field vary with geographic location. The map in sea turtles was the first ever described though similar abilities have now been reported in lobsters,fish, and birds.[49] Magnetoreception by land turtles was shown in a 2010 experiment on Terrapene carolina, a box turtle. After teaching a group of these box turtles to swim to either the east or west end of an experimental tank, a strong magnet disrupted the learned routes.[50][51]
Orientation toward the sea, as seen in turtle hatchlings, may rely partly on magnetoreception. In loggerhead and leatherback turtles, breeding takes place on beaches, and, after hatching, offspring crawl rapidly to the sea. Although differences in light density seem to drive this behaviour, magnetic alignment appears to play a part. For instance, the natural directional preferences held by these hatchlings (which lead them from beaches to the sea) reverse upon experimental inversion of the magnetic poles.[52]
In birds
Homing pigeons use magnetic fields as part of their complex navigation system.[53] William Keeton showed that time-shifted homing pigeons (acclimatised in the laboratory to a different time-zone) are unable to orient themselves correctly on a clear, sunny day; this is attributed to time-shifted pigeons being unable to compensate accurately for the movement of the sun during the day. Conversely, time-shifted pigeons released on overcast days navigate correctly, suggesting that pigeons can use magnetic fields to orient themselves; this ability can be disrupted with magnets attached to the birds' backs.[54][55] Pigeons can detect magnetic anomalies as weak as 1.86 gauss.[56]
For a long time the trigeminal system was the suggested location for a magnetite-based magnetoreceptor in the pigeon. This was based on two findings: First, magnetite-containing cells were reported in specific locations in the upper beak.[57] However, the cells proved to be immune system macrophages, not neurons able to detect magnetic fields.[58][59] Second, pigeon magnetic field detection is impaired by sectioning the trigeminal nerve and by application of lidocaine, an anaesthetic, to the olfactory mucosa.[60] However, lidocaine treatment might lead to unspecific effects and not represent a direct interference with potential magnetoreceptors.[59] As a result, an involvement of the trigeminal system is still debated. In the search for magnetite receptors, a large iron-containing organelle (the cuticulosome) of unknown function was found in the inner ear of pigeons.[61][62] Areas of the pigeon brain that respond with increased activity to magnetic fields are the posterior vestibular nuclei, dorsal thalamus, hippocampus, and visual hyperpallium.[63]
Domestic hens have iron mineral deposits in the sensory dendrites in the upper beak and are capable of magnetoreception.[64][65] Beak trimming causes loss of the magnetic sense.[66]
In mammals
Some mammals are capable of magnetoreception. When woodmice are removed from their home area and deprived of visual and olfactory cues, they orient towards their homes until an inverted magnetic field is applied to their cage.[67] When the same mice are allowed access to visual cues, they are able to orient themselves towards home despite the presence of inverted magnetic fields. This indicates that woodmice use magnetic fields to orient themselves when no other cues are available. The magnetic sense of woodmice is likely based on a radical-pair mechanism.[68]

The Zambian mole-rat, a subterranean mammal, uses magnetic fields to aid in nest orientation.[70] In contrast to woodmice, Zambian mole-rats do not rely on radical-pair based magnetoreception, perhaps due to their subterranean lifestyle. Experimental exposure to magnetic fields leads to an increase in neural activity within the superior colliculus, as measured by immediate gene expression. The activity level of neurons within two levels of the superior colliculus, the outer sublayer of the intermediate gray layer and the deep gray layer, were elevated in a non-specific manner when exposed to various magnetic fields. However, within the inner sublayer of the intermediate gray layer (InGi) there were two or three clusters of cells that respond in a more specific manner. The more time the mole rats were exposed to a magnetic field, the greater the immediate early gene expression within the InGi.[69]
Bats may use magnetic fields to orient themselves. They use echolocation to navigate over short distances. When big brown bats (Eptesicus fuscus) are taken from their home roosts and exposed to magnetic fields rotated 90 degrees from magnetic north, they are disoriented; it is unclear whether they use the magnetic sense as a map, a compass, or a compass calibrator.[71] Another bat species, the greater mouse-eared bat (Myotis myotis), appear to use the earth's magnetic field to provide their compass, but to calibrate this with the direction of sunset.[72]
Red foxes (Vulpes vulpes) may use magnetoreception when predating small rodents like mice and voles. They attack this kind of prey using a specific high-jump, preferring a north-eastern compass direction. Successful attacks are tightly clustered to the north.[73]
It is unknown whether humans can sense magnetic fields.[74] The ethmoid bone in the nose contains magnetic materials.[75] Magnetosensitive cryptochrome 2 (cry2) is present in the human retina.[76] Human alpha brain waves are affected by magnetic fields, but it is not known whether behaviour is affected.[74][76]
References
- Winklhofer, Michael (3 February 2010). "Magnetoreception". Journal of the Royal Society Interface. 7 (suppl_2): S131-4. doi:10.1098/rsif.2010.0010.focus. PMC 2843998. PMID 20129954.
- Wiltschko, Roswitha; Wiltschko, Wolfgang (27 September 2019). "Magnetoreception in Birds". Journal of the Royal Society Interface. 16 (158): 20190295. doi:10.1098/rsif.2019.0295. PMC 6769297. PMID 31480921.
- Wiltschko, Wolfgang; Wiltschko, Roswitha (August 2008). "Magnetic orientation and magnetoreception in birds and other animals". Journal of Comparative Physiology A. 191 (8): 675–693. doi:10.1007/s00359-005-0627-7. PMID 15886990. S2CID 206960525.
- Hore, Peter J.; Mouritsen, Henrik (April 2022). "The Quantum Nature of Bird Migration". Scientific American: 24–29.
- Hore, Peter J.; Mouritsen, Henrik (5 July 2016). "The Radical-Pair Mechanism of Magnetoreception". Annual Review of Biophysics. 45 (1): 299–344. doi:10.1146/annurev-biophys-032116-094545. PMID 27216936. S2CID 7099782.
- Rodgers, Christopher (1 January 2009). "Magnetic field effects in chemical systems". Pure and Applied Chemistry. 81 (1): 19–43. doi:10.1351/PAC-CON-08-10-18. S2CID 96850994.
- Steiner, Ulrich E.; Ulrich, Thomas (1 January 1989). "Magnetic field effects in chemical kinetics and related phenomena". Chemical Reviews. 89 (1): 51–147. doi:10.1021/cr00091a003.
- Woodward, J. R. (1 September 2002). "Radical pairs in solution". Progress in Reaction Kinetics and Mechanism. 27 (3): 165–207. doi:10.3184/007967402103165388. S2CID 197049448.
- Rodgers, C. T.; Hore, Peter J. (2009). "Chemical magnetoreception in birds: The radical pair mechanism". PNAS. 106 (2): 353–360. Bibcode:2009PNAS..106..353R. doi:10.1073/pnas.0711968106. PMC 2626707. PMID 19129499.
- Schulten, Klaus; Swenberg, Charles E.; Weiler, Albert (1 January 1978). "A Biomagnetic Sensory Mechanism Based on Magnetic Field Modulated Coherent Electron Spin Motion". Zeitschrift für Physikalische Chemie. 111 (1): 1–5. doi:10.1524/zpch.1978.111.1.001. S2CID 124644286.
- Solov'yov, Ilia; Schulten, Klaus. "Cryptochrome and Magnetic Sensing". Theoretical and Computational Biophysics Group, University of Illinois Urbana-Champaign. Retrieved 10 January 2022.
- Wiltschko, Roswitha; Ahmad, Margaret; Nießner, Christine; Gehring, Dennis; Wiltschko, Wolfgang (2016). "Light-dependent magnetoreception in birds: the crucial step occurs in the dark". Journal of the Royal Society Interface. 13 (118): 20151010. doi:10.1098/rsif.2015.1010. PMC 4892254. PMID 27146685. A supplement to the paper summarizes alternative hypotheses on avian compass mechanisms.
- Hiscock, Hamish G.; Worster, Susannah; Kattnig, Daniel R.; Steers, Charlotte; Jin, Ye; Manolopoulos, David E.; Mouritsen, Henrik; Hore, P. J. (26 April 2016). "The quantum needle of the avian magnetic compass". PNAS. 113 (17): 4634–4639. Bibcode:2016PNAS..113.4634H. doi:10.1073/pnas.1600341113. PMC 4855607. PMID 27044102.
- Kattnig, Daniel R. (26 October 2017). "Radical-Pair-Based Magnetoreception Amplified by Radical Scavenging: Resilience to Spin Relaxation". The Journal of Physical Chemistry B. 121 (44): 10215–10227. doi:10.1021/acs.jpcb.7b07672. hdl:10871/30371. PMID 29028342.
- Fleissner, Gerta; Holtkamp-Rötzler, Elke; Hanzlik, Marianne; Winklhofer, Michael; Fleissner, Günther; Petersen, Nikolai; Wiltschko, Wolfgang (26 February 2003). "Ultrastructural analysis of a putative magnetoreceptor in the beak of homing pigeons". Journal of Comparative Neurology. 458 (4): 350–360. doi:10.1002/cne.10579. PMID 12619070. S2CID 36992055.
- Fleissner, Guenther; Wellenreuther, Gerd; Heyers, Dominik; et al. (16 February 2010). "Avian Magnetoreception: Elaborate Iron Mineral Containing Dendrites in the Upper Beak Seem to Be a Common Feature of Birds". PLOS One. 5 (2): e9231. Bibcode:2010PLoSO...5.9231F. doi:10.1371/journal.pone.0009231. PMC 2821931. PMID 20169083.
- Solov’yov, Ilia A.; Greiner, Walter (September 2007). "Theoretical Analysis of an Iron Mineral-Based Magnetoreceptor Model in Birds". Biophysical Journal. 93 (5): 1493–1509. Bibcode:2007BpJ....93.1493S. doi:10.1529/biophysj.107.105098. PMC 1948037. PMID 17496012.
- Keays, David Anthony; Shaw, Jeremy; Lythgoe, Mark; Saunders, Martin; Cadiou, Herve; Pichler, Paul; Breuss, Martin; Sugar, Cristina; Edelman, Nathaniel (April 2012). "Clusters of iron-rich cells in the upper beak of pigeons are macrophages not magnetosensitive neurons". Nature. 484 (7394): 367–370. Bibcode:2012Natur.484..367T. doi:10.1038/nature11046. PMID 22495303. S2CID 205228624.
- Kishkinev, D. A.; Chernetsov, N. S. (2015). "Magnetoreception systems in birds: A review of current research". Biology Bulletin Reviews. 5 (1): 46–62. doi:10.1134/s2079086415010041. S2CID 18229682.
- Newton, Kyle C.; Gill, Andrew B.; Kajiura, Stephen M. (2020). "Electroreception in marine fishes: chondrichthyans". Journal of Fish Biology. 95 (1): 135–154. doi:10.1111/jfb.14068. PMID 31169300. S2CID 174812242.
- Blonder, Barbara I.; Alevizon, William S. (1988). "Prey Discrimination and Electroreception in the Stingray Dasyatis sabina". Copeia. 1988 (1): 33–36. doi:10.2307/1445919. JSTOR 1445919.
- Kalmijn, A. J. (1 October 1971). "The Electric Sense of Sharks and Rays". Journal of Experimental Biology. 55 (2): 371–383. doi:10.1242/jeb.55.2.371. PMID 5114029.
- Anderson, James M.; Clegg, Tamrynn M.; Véras, Luisa V. M. V. Q.; Holland, Kim N. (8 September 2017). "Insight into shark magnetic field perception from empirical observations". Scientific Reports. 7 (1): 11042. Bibcode:2017NatSR...711042A. doi:10.1038/s41598-017-11459-8. PMC 5591188. PMID 28887553.
- Pósfai, Mihály; Lefèvre, Christopher T.; Trubitsyn, Denis; Bazylinski, Dennis A.; Frankel, Richard B. (2013). "Phylogenetic significance of composition and crystal morphology of magnetosome minerals". Frontiers in Microbiology. 4: 344. doi:10.3389/fmicb.2013.00344. PMC 3840360. PMID 24324461.
- Blakemore, Richard (1975). "Magnetotactic Bacteria". Science. 190 (4212): 377–379. Bibcode:1975Sci...190..377B. doi:10.1126/science.170679. PMID 170679. S2CID 5139699.
- Bellini, Salvatore (27 March 2009). "On a unique behavior of freshwater bacteria". Chinese Journal of Oceanology and Limnology. 27 (1): 3–5. Bibcode:2009ChJOL..27....3B. doi:10.1007/s00343-009-0003-5. S2CID 86828549.
- Bellini, Salvatore (27 March 2009). "Further studies on "magnetosensitive bacteria"". Chinese Journal of Oceanology and Limnology. 27 (1): 6–12. Bibcode:2009ChJOL..27....6B. doi:10.1007/s00343-009-0006-2. S2CID 86147382.
- Natan, Eviatar; Fitak, Robert Rodgers; Werber, Yuval; Vortman, Yoni (28 September 2020). "Symbiotic magnetic sensing: raising evidence and beyond". Philosophical Transactions of the Royal Society B. 375 (1808): 20190595. doi:10.1098/rstb.2019.0595. PMC 7435164. PMID 32772668.
- Gould, J. L. (2008). "Animal navigation: The evolution of magnetic orientation". Current Biology. 18 (11): R482–R48. doi:10.1016/j.cub.2008.03.052. PMID 18522823. S2CID 10961495.
- Wiltschko, Roswitha; Wiltschko, Wolfgang (2012). "Chapter 8 – Magnetoreception". In Carlos López-Larrea (ed.). Sensing in Nature. Advances in Experimental Medicine and Biology. Vol. 739. Springer. doi:10.1007/978-1-4614-1704-0. ISBN 978-1-4614-1703-3. S2CID 41131723.
- Maffei, Massimo E. (4 September 2014). "Magnetic field effects on plant growth, development, and evolution". Frontiers in Plant Science. 5: 445. doi:10.3389/fpls.2014.00445. PMC 4154392. PMID 25237317.
- Lohmann, K. J.; Willows, A. O. D. (1987). "Lunar-Modulated Geomagnetic Orientation by a Marine Mollusk". Science. 235 (4786): 331–334. Bibcode:1987Sci...235..331L. doi:10.1126/science.3798115. PMID 3798115.
- Lohmann, K. J.; Willows; Pinter, R. B. (1991). "An identifiable molluscan neuron responds to changes in earth-strength magnetic fields". Journal of Experimental Biology. 161: 1–24. doi:10.1242/jeb.161.1.1. PMID 1757771.
- Wang, John H.; Cain, Shaun D.; Lohmann, Kenneth J. (22 February 2004). "Identifiable neurons inhibited by Earth-strength magnetic stimuli in the mollusc Tritonia diomedea". Journal of Experimental Biology. 207 (6): 1043–1049. doi:10.1242/jeb.00864. PMID 14766962. S2CID 13439801.
- Wyeth, Russell C.; Holden, Theora; Jalala, Hamed; Murray, James A. (1 April 2021). "Rare-Earth Magnets Influence Movement Patterns of the Magnetically Sensitive Nudibranch Tritonia exsulans in Its Natural Habitat". The Biological Bulletin. 240 (2): 105–117. doi:10.1086/713663. PMID 33939940. S2CID 233485664.
- Gegear, Robert J.; Casselman, Amy; Waddell, Scott; Reppert, Steven M. (August 2008). "Cryptochrome mediates light-dependent magnetosensitivity in Drosophila". Nature. 454 (7207): 1014–1018. Bibcode:2008Natur.454.1014G. doi:10.1038/nature07183. PMC 2559964. PMID 18641630.
- Pereira-Bomfim, M.D.G.C.; Antonialli-Junior, W.F.; Acosta-Avalos, D. (2015). "Effect of magnetic field on the foraging rhythm and behavior of the swarm-founding paper wasp Polybia paulista Ihering (hymenoptera: vespidae)". Sociobiology. 62 (1): 99–104.
- Wajnberg, E.; Acosta-Avalos, D.; Alves, O.C.; de Oliveira, J.F.; Srygley, R.B.; Esquivel, D.M. (2010). "Magnetoreception in eusocial insects: An update". Journal of the Royal Society Interface. 7 (Suppl 2): S207–S225. doi:10.1098/rsif.2009.0526.focus. PMC 2843992. PMID 20106876.
- Esquivel, Darci M.S.; Wajnberg, E.; do Nascimento, F.S.; Pinho, M.B.; Lins de Barros, H.G.P.; Eizemberg, R. (2005). "Do Magnetic Storms Change Behavior of the Stingless Bee Guiriçu (Schwarziana quadripunctata)?". Naturwissenschaften. 94 (2): 139–142. doi:10.1007/s00114-006-0169-z. PMID 17028885. S2CID 10746883.
- Lucano, M.J.; Cernicchiaro, G.; Wajnberg, E.; Esquivel, D.M.S. (2005). "Stingless Bee Antennae: A Magnetic Sensory Organ?". BioMetals. 19 (3): 295–300. doi:10.1007/s10534-005-0520-4. PMID 16799867. S2CID 10162385.
- Quinn, Thomas P. (1980). "Evidence for celestial and magnetic compass orientation in lake migrating sockeye salmon fry". Journal of Comparative Physiology A. 137 (3): 243–248. doi:10.1007/bf00657119. S2CID 44036559.
- Taylor, P. B. (May 1986). "Experimental evidence for geomagnetic orientation in juvenile salmon, Oncorhynchus tschawytscha Walbaum". Journal of Fish Biology. 28 (5): 607–623. doi:10.1111/j.1095-8649.1986.tb05196.x.
- Phillips, John B. (1977). "Use of the earth's magnetic field by orienting cave salamanders (Eurycea lucifuga)". Journal of Comparative Physiology. 121 (2): 273–288. doi:10.1007/bf00609616. S2CID 44654348.
- Phillips, John B. (1986). "Magnetic compass orientation in the Eastern red-spotted newt (Notophthalmus viridescens)". Journal of Comparative Physiology A. 158 (1): 103–109. doi:10.1007/bf00614524. PMID 3723427. S2CID 25252103.
- Phillips, John B. (15 August 1986). "Two magnetoreception pathways in a migratory salamander". Science. 233 (4765): 765–767. Bibcode:1986Sci...233..765P. doi:10.1126/science.3738508. PMID 3738508. S2CID 28292152.
- Sinsch, Ulrich (1987). "Orientation behaviour of toads (Bufo bufo) displaced from the breeding site". Journal of Comparative Physiology A. 161 (5): 715–727. doi:10.1007/bf00605013. PMID 3119823. S2CID 26102029.
- Sinsch, Ulrich (January 1992). "Sex-biassed site fidelity and orientation behaviour in reproductive natterjack toads (Bufo calamita)". Ethology Ecology & Evolution. 4 (1): 15–32. doi:10.1080/08927014.1992.9525347.
- Lohmann, K.J. (1991). "Magnetic orientation by hatchling loggerhead sea turtles (Caretta caretta)". Journal of Experimental Biology. 155: 37–49. doi:10.1242/jeb.155.1.37. PMID 2016575.
- Lohmann, Kenneth J.; Goforth, Kayla M.; Mackiewicz, Alayna G.; Lim, Dana S.; Lohmann, Catherine M.F. (2022). "agnetic maps in animal navigation". J Comp Physiol A. 208 (1): 41–67. doi:10.1007/s00359-021-01529-8. PMC 8918461. PMID 34999936.
- Mathis, Alicia; Moore, Frank R. (26 April 2010). "Geomagnetism and the Homeward Orientation of the Box Turtle, Terrapene Carolina". Ethology. 78 (4): 265–274. doi:10.1111/j.1439-0310.1988.tb00238.x.
- G., Stehli, F. (1996). Magnetite Biomineralization and Magnetoreception in Organisms: A new biomagnetism. Springer. ISBN 978-1-4613-0313-8. OCLC 958527742.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Merrill, Maria W.; Salmon, Michael (30 September 2010). "Magnetic orientation by hatchling loggerhead sea turtles (Caretta caretta) from the Gulf of Mexico". Marine Biology. 158 (1): 101–112. doi:10.1007/s00227-010-1545-y. S2CID 84391053.
- Walcott, C. (1996). "Pigeon homing: observations, experiments and confusions". Journal of Experimental Biology. 199 (Pt 1): 21–27. doi:10.1242/jeb.199.1.21. PMID 9317262.
- Keeton, W. T. (1971). "Magnets interfere with pigeon homing". PNAS. 68 (1): 102–106. Bibcode:1971PNAS...68..102K. doi:10.1073/pnas.68.1.102. PMC 391171. PMID 5276278.
- Gould, J. L. (1984). "Magnetic field sensitivity in animals". Annual Review of Physiology. 46: 585–598. doi:10.1146/annurev.ph.46.030184.003101. PMID 6370118.
- Mora, C. V.; Davison, M.; Wild, J. M.; Walker, M. M. (2004). "Magnetoreception and its trigeminal mediation in the homing pigeon". Nature. 432 (7016): 508–511. Bibcode:2004Natur.432..508M. doi:10.1038/nature03077. PMID 15565156. S2CID 2485429.
- Fleissner, Gerta; Holtkamp-Rötzler, Elke; Hanzlik, Marianne; Winklhofer, Michael; Fleissner, Günther; Petersen, Nikolai; Wiltschko, Wolfgang (26 February 2003). "Ultrastructural analysis of a putative magnetoreceptor in the beak of homing pigeons". Journal of Comparative Neurology. 458 (4): 350–360. doi:10.1002/cne.10579. PMID 12619070. S2CID 36992055.
- Treiber, Christoph Daniel; Salzer, Marion Claudia; Riegler, Johannes; et al. (11 April 2012). "Clusters of iron-rich cells in the upper beak of pigeons are macrophages not magnetosensitive neurons". Nature. 484 (7394): 367–370. Bibcode:2012Natur.484..367T. doi:10.1038/nature11046. PMID 22495303. S2CID 205228624.
- Engels, Svenja; Treiber, Christoph Daniel; Salzer, Marion Claudia; et al. (1 August 2018). "Lidocaine is a nocebo treatment for trigeminally mediated magnetic orientation in birds". Journal of the Royal Society Interface. 15 (145): 20180124. doi:10.1098/rsif.2018.0124. PMC 6127160. PMID 30089685.
- Wiltschko, Roswitha; Schiffner, Ingo; Fuhrmann, Patrick; Wiltschko, Wolfgang (September 2010). "The Role of the Magnetite-Based Receptors in the Beak in Pigeon Homing". Current Biology. 20 (17): 1534–1538. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2010.06.073. PMID 20691593. S2CID 15896143.
- Lauwers, Mattias; Pichler, Paul; Edelman, Nathaniel Bernard; et al. (May 2013). "An Iron-Rich Organelle in the Cuticular Plate of Avian Hair Cells". Current Biology. 23 (10): 924–929. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2013.04.025. PMID 23623555. S2CID 9052155.
- Nimpf, Simon; Malkemper, Erich Pascal; Lauwers, Mattias; et al. (15 November 2017). "Subcellular analysis of pigeon hair cells implicates vesicular trafficking in cuticulosome formation and maintenance". eLife. 6. doi:10.7554/elife.29959. PMC 5699870. PMID 29140244.
- Wu, L.-Q.; Dickman, J. D. (2011). "Magnetoreception in an avian brain in part mediated by inner ear lagena". Current Biology. 21 (5): 418–23. doi:10.1016/j.cub.2011.01.058. PMC 3062271. PMID 21353559.
- Falkenberg, G.; Fleissner, G.; Schuchardt, K.; et al. (2010). "Avian magnetoreception: Elaborate iron mineral containing dendrites in the upper beak seem to be a common feature of birds". PLOS One. 5 (2): e9231. Bibcode:2010PLoSO...5.9231F. doi:10.1371/journal.pone.0009231. PMC 2821931. PMID 20169083.
- Wiltschko, Wolfgang; Freire, Rafael; Munro, Ursula; Ritz, Thorsten; Rogers, Lesley; Thalau, Peter; Wiltschko, Roswitha (1 July 2007). "The magnetic compass of domestic chickens, Gallus gallus". Journal of Experimental Biology. 210 (13): 2300–2310. doi:10.1242/jeb.004853. hdl:10453/5735. PMID 17575035. S2CID 9163408.
- Freire, R.; Eastwood, M. A.; Joyce, M. (2011). "Minor beak trimming in chickens leads to loss of mechanoreception and magnetoreception". Journal of Animal Science. 89 (4): 1201–1206. doi:10.2527/jas.2010-3129. PMID 21148779.
- Mather, J. G.; Baker, R. R. (1981). "Magnetic sense of direction in woodmice for route-based navigation". Nature. 291 (5811): 152–155. Bibcode:1981Natur.291..152M. doi:10.1038/291152a0. S2CID 4262309.
- Malkemper, E. Pascal; Eder, Stephan H. K.; Begall, Sabine; Phillips, John B.; Winklhofer, Michael; Hart, Vlastimil; Burda, Hynek (29 April 2015). "Magnetoreception in the wood mouse ( Apodemus sylvaticus ): influence of weak frequency-modulated radio frequency fields". Scientific Reports. 5 (1): 9917. Bibcode:2015NatSR...4E9917M. doi:10.1038/srep09917. PMC 4413948. PMID 25923312.
- Nemec, P.; Altmann, J.; Marhold, S.; Burda, H.; Oelschlager, H. H. (2001). "Neuroanatomy of magnetoreception: The superior colliculus involved in magnetic orientation in a mammal". Science. 294 (5541): 366–368. Bibcode:2001Sci...294..366N. doi:10.1126/science.1063351. PMID 11598299. S2CID 41104477.
- Marhold, S.; Wiltschko, Wolfgang; Burda, H. (1997). "A magnetic polarity compass for direction finding in a subterranean mammal". Naturwissenschaften. 84 (9): 421–423. Bibcode:1997NW.....84..421M. doi:10.1007/s001140050422. S2CID 44399837.
- Holland, R. A.; Thorup, K.; Vonhof, M. J.; Cochran, W. W.; Wikelski, M. (2006). "Bat orientation using Earth's magnetic field". Nature. 444 (7120): 702. Bibcode:2006Natur.444..702H. doi:10.1038/444702a. PMID 17151656. S2CID 4379579.
- Holland, Richard A.; Borissov, Ivailo; Siemers, Björn M. (29 March 2010). "A nocturnal mammal, the greater mouse-eared bat, calibrates a magnetic compass by the sun". PNAS. 107 (15): 6941–6945. Bibcode:2010PNAS..107.6941H. doi:10.1073/pnas.0912477107. ISSN 0027-8424. PMC 2872435. PMID 20351296.
- Cressey, Daniel (12 January 2011). "Fox 'rangefinder' sense expands the magnetic menagerie". Nature Publishing Group / Macmillan. Archived from the original on 24 June 2014. Retrieved 6 June 2014.
- Wang, Connie X.; Hilburn, Isaac A.; Wu, Daw-An; et al. (2019). "Transduction of the Geomagnetic Field as Evidenced from alpha-Band Activity in the Human Brain". eNeuro. Society for Neuroscience. 6 (2): ENEURO.0483–18.2019. doi:10.1523/eneuro.0483-18.2019. ISSN 2373-2822. PMC 6494972. PMID 31028046.
- Carrubba, S.; Frilot, C.; Chesson, A.L.; Marino, A.A. (5 January 2007). "Evidence of a nonlinear human magnetic sense". Neuroscience. 144 (1): 356–357. doi:10.1016/j.neuroscience.2006.08.068. PMID 17069982. S2CID 34652156.
- Foley, Lauren E.; Gegear, Robert J.; Reppert, Steven M. (2011). "Human cryptochrome exhibits light-dependent magnetosensitivity". Nature Communications. 2: 356. Bibcode:2011NatCo...2..356F. doi:10.1038/ncomms1364. PMC 3128388. PMID 21694704.