Mesozoic marine revolution
The Mesozoic marine revolution (MMR) refers to the increase in shell-crushing (durophagous) and boring predation in benthic organisms throughout the Mesozoic era (251 Mya to 66 Mya), along with bulldozing and sediment remodelling in marine habitats.[1] The term was first coined by Geerat J. Vermeij,[2] who based his work on that of Steven M. Stanley.[3][4][5] While the MMR was initially restricted to the Cretaceous (145 Mya to 66 Mya), more recent studies have suggested that the beginning of this ecological arms race extends as far back as the Triassic,[6][7][8] with the MMR now being considered to have started in the Anisian[9] or the Aalenian.[10] It is an important transition between the Palaeozoic evolutionary fauna and the Modern evolutionary fauna that occurred throughout the Mesozoic.


The Mesozoic marine revolution was not the first bout of increased predation pressure; that occurred around the end of the Ordovician.[11] There is some evidence of adaptation to durophagy during the Palaeozoic, particularly in crinoids.[12]
Causes
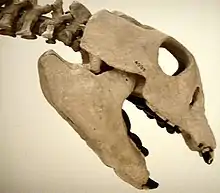

The Mesozoic marine revolution was driven by the evolution of shell-crushing behaviour among Mesozoic marine predators, the technique being perfected in the Late Cretaceous. This forced shelled marine invertebrates to develop defences against such predation or be wiped out. The consequences of this can be seen in many invertebrates today. Such predators are thought to include: Triassic placodonts, Triassic ichthyosaurs, Triassic omphalosaurids, Triassic plesiosaurs, Jurassic pliosaurs, Late Cretaceous mosasaurs and Cretaceous ptychodontoid sharks.[2] Many gastropods also evolved to feed on prey with shells.[13] However, because most durophagous predators were generalists, their effect on anti-predator shell architecture has been viewed by some as diffuse and not as extensive as other authors have suggested.[14]
It is thought that the break-up of Pangaea and the formation of new oceans throughout the Mesozoic brought together previously isolated marine communities, forcing them to compete and adapt. The increased shelf space caused by sea-level rise and a hyper-greenhouse climate provided more iterations and chances to evolve, resulting in increasing diversity.[2]
The evolution of angiosperms in the Cretaceous enhanced the hydrological cycling, speeding up rates of weathering and nutrient flow into the oceans, which has been cited as a possible driver of the MMR.[15]
Another proposal is the evolution of hermit crabs. These exploit the shells of dead gastropods, effectively doubling the life-span of the shell. This allows durophagous predators nearly twice the prey, making it a viable niche to exploit.[2]
Effects
The net result of the Mesozoic marine revolution was a change from the sedentary epifaunal lifestyle of the Palaeozoic evolutionary fauna to the infaunal/planktonic mode of life of the modern fauna.[5] Non-mobile types that failed to re-attach to their substrate (such as brachiopods) when removed were picked off as easy prey, whereas those that could hide from predation or be mobile enough to escape had an evolutionary advantage.[2] Per capita mean metabolic rates among marine gastropods living in shallow water increased by approximately 150% from the Late Triassic to the Late Cretaceous.[16]
Three major trends can be associated with this:[17]
- Reduction in suspension feeding epifauna[17]
- Increasing abundance of infauna[17]
- An intermediate stage of mobile epifauna.[17][18]
Major casualties of the Mesozoic marine revolution include: sessile crinoids, gastropods, brachiopods and epifaunal bivalves.
Affected taxa

Gastropods
Benthic gastropods were heavily preyed upon throughout the Mesozoic Marine Revolution, the weaker shelled types being pushed out of the benthic zone into more isolated habitats. The Palaeozoic archaeogastropods were subsequently replaced by neritaceans, mesogastropods and neogastropods.[2] The former typically have symmetrical, umbilicate shells that are mechanically weaker than the latter. These lack an umbilicus and also developed the ability to modify the interior of their shells, allowing them to develop sculptures on their exterior to act as defence against predators.[2]
Another development among Muricidae was the ability to bore through shells and consume prey. These marks (while relatively rare) generally occur on sessile invertebrates, implying that they put pressure on Palaeozoic-type faunas during the Mesozoic Marine Revolution.[19]
Crinoids
The Mesozoic Marine Revolution heavily affected the crinoids, making the majority of their forms extinct. Their sessile nature made them easy prey for durophagous predators since the Triassic.[9] Survivors (such as the comatulids) could swim or crawl, behaved nocturnally or had autotomy (the ability to shed limbs in defence).[12]
The shift in the range of sessile stalked crinoids during the late Mesozoic from the shallow shelf to habitats further offshore suggests that they were forced by increased predation pressure in shallow water to migrate to a deep water refuge environment where predation pressure was lower and their mode of life more viable.[20] This migration was not globally synchronous and delayed in the Southern Hemisphere; it did not occur until the Late Eocene in Australia and Antarctica, and until the Early Miocene in Zealandia.[21]
Brachiopods
Brachiopods, the dominant benthic organism of the Palaeozoic, suffered badly during the Mesozoic Marine Revolution. Their sessile foot-attached nature made them easy prey to durophagous predators.[2] The fact that they could not re-attach to a substrate if an attack failed meant their chances of survival were slim. Unlike bivalves, brachiopods never adapted to an infaunal habit (excluding lingulids) and so remained vulnerable throughout the Mesozoic Marine Revolution. As a result of increased predation pressure on top of heightened competition with bivalves, brachiopods became a minor component of most marine faunas by the Cenozoic despite their incredible diversity and abundance during the Palaeozoic and early Mesozoic.[22]
Bivalves
Bivalves adapted more readily than the brachiopods to this ecological transition. Many bivalves adopted an infaunal habit, using their siphons to gather nutrients from the sediment-water interface while remaining safe.[2][5] Corbulids developed layers of conchiolin within their shells to better resist predation.[23] Others still, like Pecten, developed the ability to jump a short distance away from predators by contracting their valves.
Like brachiopods, epifaunal varieties of bivalves were preyed upon heavily. Among epifaunal types (such as mussels and oysters), the ability to fuse to the substrate made them more difficult to consume for smaller predators. Epifaunal bivalves were preyed on heavily before the Norian but extinction rates diminish after this.[17]
Echinoids
Echinoids do not suffer major predation (save for general infaunalisation) during the Mesozoic Marine Revolution but it is clear from bromalites (fossilised ‘vomit’) that cidaroids were consumed by predators.[24] Echinoids radiate into predatory niches and are thought to have perfected coral grazing in the Late Cretaceous.[2] Cidaroids too may have contributed to the downfall of the crinoids.[9]
References
- Erwin, Douglas H. (June 2008). "Macroevolution of ecosystem engineering, niche construction and diversity". Trends in Ecology & Evolution. 23 (6): 304–310. doi:10.1016/j.tree.2008.01.013. PMID 18457902. Retrieved 4 April 2023.
- Vermeij, G. J. (1977). "The Mesozoic Marine Revolution: Evidence from Snails, Predators and Grazers". Paleobiology. 3 (3): 245–258. doi:10.1017/S0094837300005352. S2CID 54742050.
- Stanley, S. M. (2008). "Predation defeats competition on the seafloor". Paleobiology. 34 (1): 1–21. doi:10.1666/07026.1. S2CID 83713101.
- Stanley, S. M. (1974). "What has happened to the articulate brachiopods?". GSA Abstracts with Programs. 8: 966–967.
- Leighton, L. R.; Webb, A. E.; Sawyer, J. A. (2013). "Ecological Effects of the Palaeozoic-Modern faunal transition:Comparing predation on Palaeozoic brachiopods and mollusc". Geology. 41 (2): 275–278. Bibcode:2013Geo....41..275L. doi:10.1130/g33750.1.
- Tackett, Lydia S.; Tintori, Andrea (1 January 2019). "Low drilling frequency in Norian benthic assemblages from the southern Italian Alps and the role of specialized durophages during the Late Triassic". Palaeogeography, Palaeoclimatology, Palaeoecology. 513: 25–34. doi:10.1016/j.palaeo.2018.06.034.
- Tackett, Lydia S. (1 April 2016). "Late Triassic durophagy and the origin of the Mesozoic Marine Revolution". PALAIOS. 31 (4): 122–124. doi:10.2110/palo.2016.003. S2CID 88004603. Retrieved 9 December 2022.
- Cueille, Marie; Green, Emily; Duffin, Christopher J.; Hildebrandt, Claudia; Benton, Michael James (December 2020). "Fish and crab coprolites from the latest Triassic of the UK: From Buckland to the Mesozoic Marine Revolution". Proceedings of the Geologists' Association. 131 (6): 699–721. doi:10.1016/j.pgeola.2020.07.011. S2CID 226298575. Retrieved 7 January 2023.
- Salamon, M. A.; Niedzwiedzki, R.; Gorzelak, D.; Lach, R.; Surmik, D. (2012). "Bromalites from the Middle Triassic of Poland and the rise of the Mesozoic Marine Revolution". Palaeogeography, Palaeoclimatology, Palaeoecology. 321: 142–150. Bibcode:2012PPP...321..142S. doi:10.1016/j.palaeo.2012.01.029.
- Fantasia, Alicia; Mattioli, Emanuela; Spangenberg, Jorge E.; Adatte, Thierry; Bernárdez, Enrique; Ferreira, Jorge; Thibault, Nicolas; Krencker, François-Nicolas; Bodin, Stéphane (January 2022). "The middle-late Aalenian event: A precursor of the Mesozoic Marine Revolution". Global and Planetary Change. 208: 103705. doi:10.1016/j.gloplacha.2021.103705.
- Vinn, O. (2008). "Attempted predation on Early Paleozoic cornulitids". Palaeogeography, Palaeoclimatology, Palaeoecology. 273 (1–2): 87–91. doi:10.1016/j.palaeo.2008.12.004.
- Baumiller, T.; Salamon, M.; Gorzelak, P.; Mooi, R.; Messing, C.; Gahn, F. (2010). "Post-Paleozoic crinoid radiation in response to benthic predation preceded the Mesozoic marine revolution". Proceedings of the National Academy of Sciences of the United States of America. 107 (13): 5893–5896. Bibcode:2010PNAS..107.5893B. doi:10.1073/pnas.0914199107. PMC 2851891. PMID 20231453.
- Karapunar, Baran; Werner, Winfried; Simonsen, Sönke; Bade, Manfred; Lücke, Markus; Rebbe, Thomas; Schubert, Siegfried; Rojas, Alexis (11 April 2023). "Drilling predation on Early Jurassic bivalves and behavioral patterns of the presumed gastropod predator—evidence from Pliensbachian soft-bottom deposits of northern Germany". Paleobiology: 1–23. doi:10.1017/pab.2023.6. ISSN 0094-8373. Retrieved 16 September 2023.
- Walker, Sally E.; Brett, Carlton E. (October 2002). "Post-Paleozoic Patterns in Marine Predation: Was there a Mesozoic and Cenozoic Marine Predatory Revolution?". The Paleontological Society Papers. 8: 119–194. doi:10.1017/S108933260000108X. ISSN 1089-3326. Retrieved 16 September 2023.
- Boyce, C. Kevin; Lee, Jung-Eun (1 June 2011). "Could Land Plant Evolution Have Fed the Marine Revolution?". Paleontological Research. 15 (2): 100–105. doi:10.2517/1342-8144-15.2.100. ISSN 1342-8144. Retrieved 16 September 2023.
- Finnegan, Seth; McClain, Craig M.; Kosnik, Matthew A.; Payne, Jonathan L. (Spring 2011). "Escargots through time: an energetic comparison of marine gastropod assemblages before and after the Mesozoic Marine Revolution". Paleobiology. 37 (2): 252–269. doi:10.1666/09066.1. ISSN 0094-8373. Retrieved 16 September 2023.
- Tackett, L. S.; Bottjer, D. J. (2012). "Faunal Succession of Norian (Late Triassic) level-bottom benthics in the Lombardian basin: Implications for the timing, rate and nature of the early Mesozoic Marine Revolution". PALAIOS. 27 (8): 585–593. Bibcode:2012Palai..27..585T. doi:10.2110/palo.2012.p12-028r. S2CID 130270634.
- Aberhan, Martin; Kiessling, Wolfgang; Fürsich, Franz T. (Spring 2006). "Testing the role of biological interactions in the evolution of mid-Mesozoic marine benthic ecosystems". Paleobiology. 32 (2): 259–277. doi:10.1666/05028.1. ISSN 0094-8373. Retrieved 16 September 2023.
- Harper, E. M.; Forsythe, G. T.; Palmer, T. (1998). "Taphonomy and the Mesozoic Marine Revolution: Preservation masks the Importance of Boring Predators". PALAIOS. 13 (4): 352–360. doi:10.2307/3515323. JSTOR 3515323.
- Oji, T. (1996). "Is predation intensity reduced with increasing depth? Evidence from the west Atlantic stalked crinoid Endoxocrinus parrae (Gervais) and implications for the Mesozoic Marine Revolution". Paleobiology. 22 (3): 339–351. doi:10.1017/S0094837300016328. S2CID 86870809.
- Whittle, Rowan J.; Hunter, Aaron W.; Cantrill, David J.; McNamara, Kenneth J. (17 May 2018). "Globally discordant Isocrinida (Crinoidea) migration confirms asynchronous Marine Mesozoic Revolution". Communications Biology. 1: 46. doi:10.1038/s42003-018-0048-0. PMC 6123680. PMID 30271929.
- Manojlovic, Marko; Clapham, Matthew E. (23 November 2020). "The role of bioturbation-driven substrate disturbance in the Mesozoic brachiopod decline". Paleobiology. 47 (1): 86–100. doi:10.1017/pab.2020.50.
- Bardhan, Subhendu; Saha, Sandip; Das, Shiladri S.; Saha, Ranita (14 April 2021). "Paleoecology of naticid–molluscan prey interaction during the Late Jurassic (Oxfordian) in Kutch, India: evolutionary implications". Journal of Paleontology. 95 (5): 974–993. doi:10.1017/jpa.2021.24. ISSN 0022-3360. Retrieved 16 September 2023.
- Borczsz, T.; Zaton, M. (2013). "The oldest record of predation on echinoids: Evidence from the M. Jurassic of Poland". Lethaia. 46: 141–145. doi:10.1111/let.12007.