Ordovician
The Ordovician (/ɔːrdəˈvɪʃi.ən, -doʊ-, -ˈvɪʃən/ or-də-VISH-ee-ən, -doh-, -VISH-ən)[9] is a geologic period and system, the second of six periods of the Paleozoic Era. The Ordovician spans 41.6 million years from the end of the Cambrian Period 485.4 million years ago (Ma) to the start of the Silurian Period 443.8 Mya.[10]
| Ordovician | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 A map of the world as it appeared in the Middle Ordovician, c. 460 Ma | |||||||||||
| Chronology | |||||||||||
| |||||||||||
| Etymology | |||||||||||
| Name formality | Formal | ||||||||||
| Name ratified | 1960 | ||||||||||
| Usage information | |||||||||||
| Celestial body | Earth | ||||||||||
| Regional usage | Global (ICS) | ||||||||||
| Time scale(s) used | ICS Time Scale | ||||||||||
| Definition | |||||||||||
| Chronological unit | Period | ||||||||||
| Stratigraphic unit | System | ||||||||||
| First proposed by | Charles Lapworth, 1879 | ||||||||||
| Time span formality | Formal | ||||||||||
| Lower boundary definition | FAD of the Conodont Iapetognathus fluctivagus | ||||||||||
| Lower boundary GSSP | Greenpoint section, Green Point, Newfoundland, Canada 49.6829°N 57.9653°W | ||||||||||
| Lower GSSP ratified | 2000[5] | ||||||||||
| Upper boundary definition | FAD of the Graptolite Akidograptus ascensus | ||||||||||
| Upper boundary GSSP | Dob's Linn, Moffat, U.K. 55.4400°N 3.2700°W | ||||||||||
| Upper GSSP ratified | 1984[6][7] | ||||||||||
| Atmospheric and climatic data | |||||||||||
| Sea level above present day | 180 m; rising to 220 m in Caradoc and falling sharply to 140 m in end-Ordovician glaciations[8] | ||||||||||
The Ordovician, named after the Welsh tribe of the Ordovices, was defined by Charles Lapworth in 1879 to resolve a dispute between followers of Adam Sedgwick and Roderick Murchison, who were placing the same rock beds in North Wales in the Cambrian and Silurian systems, respectively.[11] Lapworth recognized that the fossil fauna in the disputed strata were different from those of either the Cambrian or the Silurian systems, and placed them in a system of their own. The Ordovician received international approval in 1960 (forty years after Lapworth's death), when it was adopted as an official period of the Paleozoic Era by the International Geological Congress.
Life continued to flourish during the Ordovician as it did in the earlier Cambrian Period, although the end of the period was marked by the Ordovician–Silurian extinction events. Invertebrates, namely molluscs and arthropods, dominated the oceans, with members of the latter group probably starting their establishment on land during this time, becoming fully established by the Devonian. The first land plants are known from this period. The Great Ordovician Biodiversification Event considerably increased the diversity of life. Fish, the world's first true vertebrates, continued to evolve, and those with jaws may have first appeared late in the period. About 100 times as many meteorites struck the Earth per year during the Ordovician compared with today.[12]
Subdivisions
A number of regional terms have been used to subdivide the Ordovician Period. In 2008, the ICS erected a formal international system of subdivisions.[13] There exist Baltoscandic, British, Siberian, North American, Australian, Chinese, Mediterranean and North-Gondwanan regional stratigraphic schemes.[14]
ICS (global) subdivisions
- Upper Ordovician epoch (458.4 Ma – 443.8 Ma)
- Hirnantian stage/age (445.2 Ma – 443.8 Ma)
- Katian stage/age (453.0 Ma – 445.2 Ma)
- Sandbian stage/age (458.4 Ma – 453.0 Ma)
- Middle Ordovician epoch (470.0 Ma – 458.4 Ma)
- Darriwilian stage/age (467.3 Ma – 458.4 Ma)
- Dapingian stage/age (470.0 Ma – 467.3 Ma)
- Lower Ordovician epoch (485.4 Ma – 470.0 Ma)
- Floian stage/age (477.7 Ma – 470.0 Ma)
- Tremadocian stage/age (485.4 Ma – 477.7 Ma)
British stages and ages
The Ordovician Period in Britain was traditionally broken into Early (Tremadocian and Arenig), Middle (Llanvirn (subdivided into Abereiddian and Llandeilian) and Llandeilo) and Late (Caradoc and Ashgill) epochs. The corresponding rocks of the Ordovician System are referred to as coming from the Lower, Middle, or Upper part of the column.
The Tremadoc corresponds to the (modern) Tremadocian. The Floian corresponds to the early Arenig; the Arenig continues until the early Darriwilian, subsuming the Dapingian. The Llanvirn occupies the rest of the Darriwilian, and terminates with it at the start of the Late Ordovician. The Sandbian represents the first half of the Caradoc; the Caradoc ends in the mid-Katian, and the Ashgill represents the last half of the Katian, plus the Hirnantian.
The British ages (subdivisions of epochs) from youngest to oldest are:
"Late Ordovician"
- Hirnantian/Gamach (Ashgill)
- Rawtheyan/Richmond (Ashgill)
- Cautleyan/Richmond (Ashgill)
- Pusgillian/Maysville/Richmond (Ashgill)
"Middle Ordovician"
- Trenton (Caradoc)
- Onnian/Maysville/Eden (Caradoc)
- Actonian/Eden (Caradoc)
- Marshbrookian/Sherman (Caradoc)
- Longvillian/Sherman (Caradoc)
- Soudleyan/Kirkfield (Caradoc)
- Harnagian/Rockland (Caradoc)
- Costonian/Black River (Caradoc)
- Chazy (Llandeilo)
- Llandeilo (Llandeilo)
- Whiterock (Llanvirn)
- Llanvirn (Llanvirn)
"Early Ordovician"
- Cassinian (Arenig)
- Arenig/Jefferson/Castleman (Arenig)
- Tremadoc/Deming/Gaconadian (Tremadoc)
The Tremadoc corresponds to the (modern) Tremadocian. The Floian corresponds to the lower Arenig; the Arenig continues until the early Darriwilian, subsuming the Dapingian. The Llanvirn occupies the rest of the Darriwilian, and terminates with it at the base of the Late Ordovician. The Sandbian represents the first half of the Caradoc; the Caradoc ends in the mid-Katian, and the Ashgill represents the last half of the Katian, plus the Hirnantian.[15]
| ICS series | ICS stage | British series | British stage | North American series | North American stage | Australian stage | Chinese stage |
|---|---|---|---|---|---|---|---|
| Upper Ordovician | Hirnantian | Ashgill | Hirnantian | Cincinnatian | Gamachian | Bolindian | Hirnantian |
| Katian | Rawtheyan | Richmondian | Chientangkiangian | ||||
| Cautleyan | |||||||
| Pusgillian | Maysvillian | Eastonian | Neichianshanian | ||||
| Caradoc | Streffordian | Edenian | |||||
| Cheneyan | Mohawkian | Chatfieldian | |||||
| Sandbian | Burrellian | Gisbornian | |||||
| Turinian | |||||||
| Aurelucian | |||||||
| Whiterockian | |||||||
| Middle Ordovician | Darriwilian | Llanvirn | Llandeilo | Darriwilian | Darriwilian | ||
| Abereiddian | |||||||
| Arenig | Fennian | ||||||
| Dapingian | Yapeenian | Dapingian | |||||
| Whitlandian | Rangerian | Castlemainian | |||||
| Lower Ordovician | Floian | Ibexian | Blackhillsian | Chewtonian | Yiyangian | ||
| Bendigonian | |||||||
| Moridunian | |||||||
| Tulean | Lancefieldian | ||||||
| Tremadocian | Tremadoc | Migneintian | Xinchangian | ||||
| Stairsian | |||||||
| Cressagian | |||||||
| Skullrockian | |||||||
| Warendan |
Paleogeography and tectonics
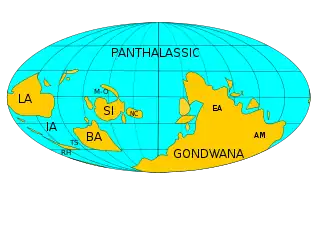
During the Ordovician, the southern continents were assembled into Gondwana, which reached from north of the equator to the South Pole. The Panthalassic Ocean, centered in the northern hemisphere, covered over half the globe.[17] At the start of the period, the continents of Laurentia (in present-day North America), Siberia, and Baltica (present-day northern Europe) were separated from Gondwana by over 5,000 kilometres (3,100 mi) of ocean. These smaller continents were also sufficiently widely separated from each other to develop distinct communities of benthic organisms.[18] The small continent of Avalonia had just rifted from Gondwana and began to move north towards Baltica and Laurentia, opening the Rheic Ocean between Gondwana and Avalonia.[19][20][21] Avalonia collided with Baltica towards the end of Ordovician.[22][23]
Other geographic features of the Ordovician world included the Tornquist Sea, which separated Avalonia from Baltica;[18] the Aegir Ocean, which separated Baltica from Siberia;[24] and an oceanic area between Siberia, Baltica, and Gondwana which expanded to become the Paleoasian Ocean in Carboniferous time. The Mongol-Okhotsk Ocean formed a deep embayment between Siberia and the Central Mongolian terranes. Most of the terranes of central Asia were part of an equatorial archipelago whose geometry is poorly constrained by the available evidence.[25]
The period was one of extensive, widespread tectonism and volcanism. However, orogenesis (mountain-building) was not primarily due to continent-continent collisions. Instead, mountains arose along active continental margins during accretion of arc terranes or ribbon microcontinents. Accretion of new crust was limited to the Iapetus margin of Laurentia; elsewhere, the pattern was of rifting in back-arc basins followed by remerger. This reflected episodic switching from extension to compression. The initiation of new subduction reflected a global reorganization of tectonic plates centered on the amalgamation of Gondwana.[26][18]
The Taconic orogeny, a major mountain-building episode, was well under way in Cambrian times.[27] This continued into the Ordovician, when at least two volcanic island arcs collided with Laurentia to form the Appalachian Mountains. Laurentia was otherwise tectonically stable. An island arc accreted to South China during the period, while subduction along north China (Sulinheer) resulted in the emplacement of ophiolites.[28]
The ash fall of the Millburg/Big Bentonite bed, at about 454 Ma, was the largest in the last 590 million years. This had a dense rock equivalent volume of as much as 1,140 cubic kilometres (270 cu mi). Remarkably, this appears to have had little impact on life.[29]
There was vigorous tectonic activity along northwest margin of Gondwana during the Floian, 478 Ma, recorded in the Central Iberian Zone of Spain. The activity reached as far as Turkey by the end of Ordovician. The opposite margin of Gondwana, in Australia, faced a set of island arcs.[18] The accretion of these arcs to the eastern margin of Gondwana was responsible for the Benambran Orogeny of eastern Australia.[30][31] Subduction also took place along what is now Argentina (Famatinian Orogeny) at 450 Ma.[32] This involved significant back arc rifting.[18] The interior of Gondwana was tectonically quiet until the Triassic.[18]
Towards the end of the period, Gondwana began to drift across the South Pole. This contributed to the Hibernian glaciation and the associated extinction event.[33]
Ordovician meteor event
The Ordovician meteor event is a proposed shower of meteors that occurred during the Middle Ordovician Epoch, about 467.5 ± 0.28 million years ago, due to the break-up of the L chondrite parent body.[34] It is not associated with any major extinction event.[35][36][37]
Geochemistry

The Ordovician was a time of calcite sea geochemistry in which low-magnesium calcite was the primary inorganic marine precipitate of calcium carbonate.[38] Carbonate hardgrounds were thus very common, along with calcitic ooids, calcitic cements, and invertebrate faunas with dominantly calcitic skeletons. Biogenic aragonite, like that composing the shells of most molluscs, dissolved rapidly on the sea floor after death.[39][40]
Unlike Cambrian times, when calcite production was dominated by microbial and non-biological processes, animals (and macroalgae) became a dominant source of calcareous material in Ordovician deposits.[41]
Climate and sea level
The Early Ordovician climate was very hot,[42] with intense greenhouse conditions and sea surface temperatures comparable to those during the Early Eocene Climatic Optimum.[43] By the late Early Ordovician, the Earth cooled,[44] giving way to a more temperate climate in the Middle Ordovician,[45] with the Earth likely entering the Early Palaeozoic Ice Age during the Sandbian,[46][47] and possibly as early as the Darriwilian[48] or even the Floian.[44] Evidence suggests that global temperatures rose briefly in the early Katian (Boda Event), depositing bioherms and radiating fauna across Europe.[49] Further cooling during the Hirnantian, at the end of the Ordovician, led to the Late Ordovician glaciation.[50]
The Ordovician saw the highest sea levels of the Paleozoic, and the low relief of the continents led to many shelf deposits being formed under hundreds of metres of water.[41] The sea level rose more or less continuously throughout the Early Ordovician, leveling off somewhat during the middle of the period.[41] Locally, some regressions occurred, but the sea level rise continued in the beginning of the Late Ordovician. Sea levels fell steadily due to the cooling temperatures for about 3 million years leading up to the Hirnantian glaciation. During this icy stage, sea level seems to have risen and dropped somewhat. Despite much study, the details remain unresolved.[41] In particular, some researches interpret the fluctuations in sea level as pre-Hibernian glaciation,[51] but sedimentary evidence of glaciation is lacking until the end of the period.[23] There is evidence of glaciers during the Hirnantian on the land we now know as Africa and South America, which were near the South Pole at the time, facilitating the formation of the ice caps of the Hirnantian glaciation.
As with North America and Europe, Gondwana was largely covered with shallow seas during the Ordovician. Shallow clear waters over continental shelves encouraged the growth of organisms that deposit calcium carbonates in their shells and hard parts. The Panthalassic Ocean covered much of the Northern Hemisphere, and other minor oceans included Proto-Tethys, Paleo-Tethys, Khanty Ocean, which was closed off by the Late Ordovician, Iapetus Ocean, and the new Rheic Ocean.
Life

For most of the Late Ordovician life continued to flourish, but at and near the end of the period there were mass-extinction events that seriously affected conodonts and planktonic forms like graptolites. The trilobites Agnostida and Ptychopariida completely died out, and the Asaphida were much reduced. Brachiopods, bryozoans and echinoderms were also heavily affected, and the endocerid cephalopods died out completely, except for possible rare Silurian forms. The Ordovician–Silurian extinction events may have been caused by an ice age that occurred at the end of the Ordovician Period, due to the expansion of the first terrestrial plants,[52] as the end of the Late Ordovician was one of the coldest times in the last 600 million years of Earth's history.
Fauna



On the whole, the fauna that emerged in the Ordovician were the template for the remainder of the Palaeozoic. The fauna was dominated by tiered communities of suspension feeders, mainly with short food chains. The ecological system reached a new grade of complexity far beyond that of the Cambrian fauna, which has persisted until the present day.[41] Though less famous than the Cambrian explosion, the Ordovician radiation (also known as the Great Ordovician Biodiversification Event)[18] was no less remarkable; marine faunal genera increased fourfold, resulting in 12% of all known Phanerozoic marine fauna.[53] Several animals also went through a miniaturization process, becoming much smaller than their Cambrian counterparts. Another change in the fauna was the strong increase in filter-feeding organisms.[54] The trilobite, inarticulate brachiopod, archaeocyathid, and eocrinoid faunas of the Cambrian were succeeded by those that dominated the rest of the Paleozoic, such as articulate brachiopods, cephalopods, and crinoids. Articulate brachiopods, in particular, largely replaced trilobites in shelf communities. Their success epitomizes the greatly increased diversity of carbonate shell-secreting organisms in the Ordovician compared to the Cambrian.[55]

Ordovician geography had its effect on the diversity of fauna; Ordovician invertebrates displayed a very high degree of provincialism.[56] The widely separated continents of Laurentia and Baltica, then positioned close to the tropics and boasting many shallow seas rich in life, developed distinct trilobite faunas from the trilobite fauna of Gondwana,[57] and Gondwana developed distinct fauna in its tropical and temperature zones.[58] The Tien Shan terrane maintained a biogeographic affinity with Gondwana,[59] and the Alborz margin of Gondwana was linked biogeographically to South China.[60] Southeast Asia's fauna also maintained strong affinities to Gondwana's.[61] North China was biogeographically connected to Laurentia and the Argentinian margin of Gondwana.[62] A Celtic biogeographic province also existed, separate from the Laurentian and Baltican ones.[63] However, tropical articulate brachiopods had a more cosmopolitan distribution, with less diversity on different continents. During the Middle Ordovician, beta diversity began a significant decline as marine taxa began to disperse widely across space.[64] Faunas become less provincial later in the Ordovician, partly due to the narrowing of the Iapetus Ocean,[65] though they were still distinguishable into the late Ordovician.[66]

Trilobites in particular were rich and diverse. Trilobites in the Ordovician were very different from their predecessors in the Cambrian. Many trilobites developed bizarre spines and nodules to defend against predators such as primitive eurypterids and nautiloids while other trilobites such as Aeglina prisca evolved to become swimming forms. Some trilobites even developed shovel-like snouts for ploughing through muddy sea bottoms. Another unusual clade of trilobites known as the trinucleids developed a broad pitted margin around their head shields.[67] Some trilobites such as Asaphus kowalewski evolved long eyestalks to assist in detecting predators whereas other trilobite eyes in contrast disappeared completely.[68] Molecular clock analyses suggest that early arachnids started living on land by the end of the Ordovician.[69] Although solitary corals date back to at least the Cambrian, reef-forming corals appeared in the early Ordovician, including the earliest known octocorals,[70][71] corresponding to an increase in the stability of carbonate and thus a new abundance of calcifying animals.[41] Brachiopods surged in diversity, adapting to almost every type of marine environment.[72][73][74] Even after GOBE, there is evidence suggesting that Ordovician brachiopods maintained elevated rates of speciation.[75] Molluscs, which appeared during the Cambrian or even the Ediacaran, became common and varied, especially bivalves, gastropods, and nautiloid cephalopods.[76][77] Cephalopods diversified from shallow marine tropical environments to dominate almost all marine environments.[78] Graptolites, which evolved in the preceding Cambrian period, thrived in the oceans.[79] This includes the distinctive Nemagraptus gracilis graptolite fauna, which was distributed widely during peak sea levels in the Sandbian.[80][23] Some new cystoids and crinoids appeared. It was long thought that the first true vertebrates (fish — Ostracoderms) appeared in the Ordovician, but recent discoveries in China reveal that they probably originated in the Early Cambrian.[81] The first gnathostome (jawed fish) may have appeared in the Late Ordovician epoch.[82] Chitinozoans, which first appeared late in the Wuliuan, exploded in diversity during the Tremadocian, quickly becoming globally widespread.[83][84] Several groups of endobiotic symbionts appeared in the Ordovician.[85][86]
In the Early Ordovician, trilobites were joined by many new types of organisms, including tabulate corals, strophomenid, rhynchonellid, and many new orthid brachiopods, bryozoans, planktonic graptolites and conodonts, and many types of molluscs and echinoderms, including the ophiuroids ("brittle stars") and the first sea stars. Nevertheless, the arthropods remained abundant; all the Late Cambrian orders continued, and were joined by the new group Phacopida. The first evidence of land plants also appeared (see evolutionary history of life).
In the Middle Ordovician, the trilobite-dominated Early Ordovician communities were replaced by generally more mixed ecosystems, in which brachiopods, bryozoans, molluscs, cornulitids, tentaculitids and echinoderms all flourished, tabulate corals diversified and the first rugose corals appeared. The planktonic graptolites remained diverse, with the Diplograptina making their appearance. One of the earliest known armoured agnathan ("ostracoderm") vertebrates, Arandaspis, dates from the Middle Ordovician.[87] During the Middle Ordovician there was a large increase in the intensity and diversity of bioeroding organisms. This is known as the Ordovician Bioerosion Revolution.[88] It is marked by a sudden abundance of hard substrate trace fossils such as Trypanites, Palaeosabella, Petroxestes and Osprioneides. Bioerosion became an important process, particularly in the thick calcitic skeletons of corals, bryozoans and brachiopods, and on the extensive carbonate hardgrounds that appear in abundance at this time.
 Upper Ordovician edrioasteroid Cystaster stellatus on a cobble from the Kope Formation in northern Kentucky with the cyclostome bryozoan Corynotrypa in the background
Upper Ordovician edrioasteroid Cystaster stellatus on a cobble from the Kope Formation in northern Kentucky with the cyclostome bryozoan Corynotrypa in the background Middle Ordovician fossiliferous shales and limestones at Fossil Mountain, west-central Utah
Middle Ordovician fossiliferous shales and limestones at Fossil Mountain, west-central Utah Outcrop of Upper Ordovician rubbly limestone and shale, southern Indiana
Outcrop of Upper Ordovician rubbly limestone and shale, southern Indiana Outcrop of Upper Ordovician limestone and minor shale, central Tennessee
Outcrop of Upper Ordovician limestone and minor shale, central Tennessee



 Brachiopods and bryozoans in an Ordovician limestone, southern Minnesota
Brachiopods and bryozoans in an Ordovician limestone, southern Minnesota Vinlandostrophia ponderosa, Maysvillian (Upper Ordovician) near Madison, Indiana (scale bar is 5.0 mm)
Vinlandostrophia ponderosa, Maysvillian (Upper Ordovician) near Madison, Indiana (scale bar is 5.0 mm) The Ordovician cystoid Echinosphaerites (an extinct echinoderm) from northeastern Estonia; approximately 5 cm in diameter
The Ordovician cystoid Echinosphaerites (an extinct echinoderm) from northeastern Estonia; approximately 5 cm in diameter Prasopora, a trepostome bryozoan from the Ordovician of Iowa
Prasopora, a trepostome bryozoan from the Ordovician of Iowa An Ordovician strophomenid brachiopod with encrusting inarticulate brachiopods and a bryozoan
An Ordovician strophomenid brachiopod with encrusting inarticulate brachiopods and a bryozoan The heliolitid coral Protaraea richmondensis encrusting a gastropod; Cincinnatian (Upper Ordovician) of southeastern Indiana
The heliolitid coral Protaraea richmondensis encrusting a gastropod; Cincinnatian (Upper Ordovician) of southeastern Indiana Zygospira modesta, atrypid brachiopods, preserved in their original positions on a trepostome bryozoan from the Cincinnatian (Upper Ordovician) of southeastern Indiana
Zygospira modesta, atrypid brachiopods, preserved in their original positions on a trepostome bryozoan from the Cincinnatian (Upper Ordovician) of southeastern Indiana Graptolites (Amplexograptus) from the Ordovician near Caney Springs, Tennessee
Graptolites (Amplexograptus) from the Ordovician near Caney Springs, Tennessee
Flora
Green algae were common in the Late Cambrian (perhaps earlier) and in the Ordovician. Terrestrial plants probably evolved from green algae, first appearing as tiny non-vascular forms resembling liverworts, in the middle to late Ordovician.[90] Fossil spores found in Ordovician sedimentary rock are typical of bryophytes.[91]

Among the first land fungi may have been arbuscular mycorrhiza fungi (Glomerales), playing a crucial role in facilitating the colonization of land by plants through mycorrhizal symbiosis, which makes mineral nutrients available to plant cells; such fossilized fungal hyphae and spores from the Ordovician of Wisconsin have been found with an age of about 460 million years ago, a time when the land flora most likely only consisted of plants similar to non-vascular bryophytes.[92]
End of the period
The Ordovician came to a close in a series of extinction events that, taken together, comprise the second largest of the five major extinction events in Earth's history in terms of percentage of genera that became extinct. The only larger one was the Permian–Triassic extinction event.
The extinctions occurred approximately 447–444 million years ago and mark the boundary between the Ordovician and the following Silurian Period. At that time all complex multicellular organisms lived in the sea, and about 49% of genera of fauna disappeared forever; brachiopods and bryozoans were greatly reduced, along with many trilobite, conodont and graptolite families.
The most commonly accepted theory is that these events were triggered by the onset of cold conditions in the late Katian, followed by an ice age, in the Hirnantian faunal stage, that ended the long, stable greenhouse conditions typical of the Ordovician.
The ice age was possibly not long-lasting. Oxygen isotopes in fossil brachiopods show its duration may have been only 0.5 to 1.5 million years.[93] Other researchers (Page et al.) estimate more temperate conditions did not return until the late Silurian.
The late Ordovician glaciation event was preceded by a fall in atmospheric carbon dioxide (from 7000 ppm to 4400 ppm).[94][95] The dip may have been caused by a burst of volcanic activity that deposited new silicate rocks, which draw CO2 out of the air as they erode.[95] Another possibility is that bryophytes and lichens, which colonized land in the middle to late Ordovician, may have increased weathering enough to draw down CO2 levels.[90] The drop in CO2 selectively affected the shallow seas where most organisms lived. As the southern supercontinent Gondwana drifted over the South Pole, ice caps formed on it, which have been detected in Upper Ordovician rock strata of North Africa and then-adjacent northeastern South America, which were south-polar locations at the time.
As glaciers grew, the sea level dropped, and the vast shallow intra-continental Ordovician seas withdrew, which eliminated many ecological niches. When they returned, they carried diminished founder populations that lacked many whole families of organisms. They then withdrew again with the next pulse of glaciation, eliminating biological diversity with each change.[96] Species limited to a single epicontinental sea on a given landmass were severely affected.[40] Tropical lifeforms were hit particularly hard in the first wave of extinction, while cool-water species were hit worst in the second pulse.[40]
Those species able to adapt to the changing conditions survived to fill the ecological niches left by the extinctions. For example, there is evidence the oceans became more deeply oxygenated during the glaciation, allowing unusual benthic organisms (Hirnantian fauna) to colonize the depths. These organisms were cosmopolitan in distribution and present at most latitudes.[66]
At the end of the second event, melting glaciers caused the sea level to rise and stabilise once more. The rebound of life's diversity with the permanent re-flooding of continental shelves at the onset of the Silurian saw increased biodiversity within the surviving Orders. Recovery was characterized by an unusual number of "Lazarus taxa", disappearing during the extinction and reappearing well into the Silurian, which suggests that the taxa survived in small numbers in refugia.[97]
An alternate extinction hypothesis suggested that a ten-second gamma-ray burst could have destroyed the ozone layer and exposed terrestrial and marine surface-dwelling life to deadly ultraviolet radiation and initiated global cooling.[98]
Recent work considering the sequence stratigraphy of the Late Ordovician argues that the mass extinction was a single protracted episode lasting several hundred thousand years, with abrupt changes in water depth and sedimentation rate producing two pulses of last occurrences of species.[99]
References
- Wellman, C.H.; Gray, J. (2000). "The microfossil record of early land plants". Phil. Trans. R. Soc. B. 355 (1398): 717–732. doi:10.1098/rstb.2000.0612. PMC 1692785. PMID 10905606.
- Korochantseva, Ekaterina; Trieloff, Mario; Lorenz, Cyrill; Buykin, Alexey; Ivanova, Marina; Schwarz, Winfried; Hopp, Jens; Jessberger, Elmar (2007). "L-chondrite asteroid breakup tied to Ordovician meteorite shower by multiple isochron 40 Ar- 39 Ar dating". Meteoritics & Planetary Science. 42 (1): 113–130. Bibcode:2007M&PS...42..113K. doi:10.1111/j.1945-5100.2007.tb00221.x.
- Lindskog, A.; Costa, M. M.; Rasmussen, C.M.Ø.; Connelly, J. N.; Eriksson, M. E. (2017-01-24). "Refined Ordovician timescale reveals no link between asteroid breakup and biodiversification". Nature Communications. 8: 14066. doi:10.1038/ncomms14066. ISSN 2041-1723. PMC 5286199. PMID 28117834.
It has been suggested that the Middle Ordovician meteorite bombardment played a crucial role in the Great Ordovician Biodiversification Event, but this study shows that the two phenomena were unrelated
- "Chart/Time Scale". www.stratigraphy.org. International Commission on Stratigraphy.
- Cooper, Roger; Nowlan, Godfrey; Williams, S. H. (March 2001). "Global Stratotype Section and Point for base of the Ordovician System" (PDF). Episodes. 24 (1): 19–28. doi:10.18814/epiiugs/2001/v24i1/005. Retrieved 6 December 2020.
- Lucas, Sepncer (6 November 2018). "The GSSP Method of Chronostratigraphy: A Critical Review". Frontiers in Earth Science. 6: 191. Bibcode:2018FrEaS...6..191L. doi:10.3389/feart.2018.00191.
- Holland, C. (June 1985). "Series and Stages of the Silurian System" (PDF). Episodes. 8 (2): 101–103. doi:10.18814/epiiugs/1985/v8i2/005. Retrieved 11 December 2020.
- Haq, B. U.; Schutter, SR (2008). "A Chronology of Paleozoic Sea-Level Changes". Science. 322 (5898): 64–68. Bibcode:2008Sci...322...64H. doi:10.1126/science.1161648. PMID 18832639. S2CID 206514545.
- "Ordovician". Dictionary.com Unabridged (Online). n.d.
- "International Chronostratigraphic Chart v.2015/01" (PDF). International Commission on Stratigraphy. January 2015.
- Charles Lapworth (1879) "On the Tripartite Classification of the Lower Palaeozoic Rocks", Geological Magazine, new series, 6 : 1-15. From pp. 13-14: "North Wales itself — at all events the whole of the great Bala district where Sedgwick first worked out the physical succession among the rocks of the intermediate or so-called Upper Cambrian or Lower Silurian system; and in all probability, much of the Shelve and the Caradoc area, whence Murchison first published its distinctive fossils — lay within the territory of the Ordovices; … Here, then, have we the hint for the appropriate title for the central system of the Lower Paleozoic. It should be called the Ordovician System, after this old British tribe."
- "New type of meteorite linked to ancient asteroid collision". Science Daily. 15 June 2016. Retrieved 20 June 2016.
- Details on the Dapingian are available at Wang, X.; Stouge, S.; Chen, X.; Li, Z.; Wang, C. (2009). "Dapingian Stage: standard name for the lowermost global stage of the Middle Ordovician Series". Lethaia. 42 (3): 377–380. doi:10.1111/j.1502-3931.2009.00169.x.
- "The Ordovician Period". Subcommission on Ordovician Stratigraphy. International Commission on Stratigraphy. 2020. Retrieved 7 June 2021.
- Ogg; Ogg; Gradstein, eds. (2008). The Concise Geological Timescale.
- Goldman, D.; Sadler, P.M.; Leslie, S.A.; Melchin, M.J.; Agterberg, F.P.; Gradstein, F.M. (2020), "The Ordovician Period", Geologic Time Scale 2020, Elsevier, pp. 631–694, doi:10.1016/b978-0-12-824360-2.00020-6, ISBN 978-0-12-824360-2, retrieved 2023-06-08
- Torsvik, Trond H.; Cocks, L. Robin M. (2017). Earth history and palaeogeography. Cambridge, United Kingdom: Cambridge University Press. p. 102. ISBN 9781107105324.
- Torsvik & Cocks 2017, p. 102.
- Pollock, Jeffrey C.; Hibbard, James P.; Sylvester, Paul J. (May 2009). "Early Ordovician rifting of Avalonia and birth of the Rheic Ocean: U–Pb detrital zircon constraints from Newfoundland". Journal of the Geological Society. 166 (3): 501–515. Bibcode:2009JGSoc.166..501P. doi:10.1144/0016-76492008-088. S2CID 129091590.
- Nance, R. Damian; Gutiérrez-Alonso, Gabriel; Keppie, J. Duncan; Linnemann, Ulf; Murphy, J. Brendan; Quesada, Cecilio; Strachan, Rob A.; Woodcock, Nigel H. (March 2012). "A brief history of the Rheic Ocean". Geoscience Frontiers. 3 (2): 125–135. doi:10.1016/j.gsf.2011.11.008.
- Torsvik & Cocks 2017, p. 103.
- Trela, Wieslaw (15 July 2005). "Condensation and phosphatization of the Middle and Upper Ordovician limestones on the Malopolska Block (Poland): Response to paleoceanographic conditions". Sedimentary Geology. 117 (3–4): 219–236. doi:10.1016/j.sedgeo.2005.05.005. Retrieved 21 May 2023.
- Torsvik & Cocks 2017, p. 112.
- Torsvik, Trond H.; Rehnström, Emma F. (March 2001). "Cambrian palaeomagnetic data from Baltica: implications for true polar wander and Cambrian palaeogeography". Journal of the Geological Society. 158 (2): 321–329. Bibcode:2001JGSoc.158..321T. doi:10.1144/jgs.158.2.321. S2CID 54656066.
- Torsvik & Cocks 2017, pp. 102, 106.
- van Staal, C.R.; Hatcher, R.D. Jr. (2010). "Global setting of Ordovician orogenesis". Geol Soc Am Spec Pap. 466: 1–11. doi:10.1130/2010.2466(01). ISBN 9780813724669.
- Torsvik & Cocks 2017, pp. 93–94.
- Torsvik & Cocks 2017, pp. 106–109.
- Huff, Warren D.; Bergström, Stig M.; Kolata, Dennis R. (1992-10-01). "Gigantic Ordovician volcanic ash fall in North America and Europe: Biological, tectonomagmatic, and event-stratigraphic significance". Geology. 20 (10): 875–878. Bibcode:1992Geo....20..875H. doi:10.1130/0091-7613(1992)020<0875:GOVAFI>2.3.CO;2.
- Glen, R. A.; Meffre, S.; Scott, R. J. (March 2007). "Benambran Orogeny in the Eastern Lachlan Orogen, Australia". Australian Journal of Earth Sciences. 54 (2–3): 385–415. Bibcode:2007AuJES..54..385G. doi:10.1080/08120090601147019. S2CID 129843558.
- Torsvik & Cocks 2017, p. 105.
- Ramos, Victor A. (2018). "The Famatinian Orogen Along the Protomargin of Western Gondwana: Evidence for a Nearly Continuous Ordovician Magmatic Arc Between Venezuela and Argentina". The Evolution of the Chilean-Argentinean Andes. Springer Earth System Sciences: 133–161. doi:10.1007/978-3-319-67774-3_6. ISBN 978-3-319-67773-6.
- Torsvik & Cocks 2017, pp. 103–105.
- Lindskog, A.; Costa, M. M.; Rasmussen, C.M.Ø.; Connelly, J. N.; Eriksson, M. E. (2017-01-24). "Refined Ordovician timescale reveals no link between asteroid breakup and biodiversification". Nature Communications. 8: 14066. Bibcode:2017NatCo...814066L. doi:10.1038/ncomms14066. ISSN 2041-1723. PMC 5286199. PMID 28117834.
- Heck, Philipp R.; Schmitz, Birger; Baur, Heinrich; Halliday, Alex N.; Wieler, Rainer (2004). "Fast delivery of meteorites to Earth after a major asteroid collision". Nature. 430 (6997): 323–5. Bibcode:2004Natur.430..323H. doi:10.1038/nature02736. PMID 15254530. S2CID 4393398.
- Haack, Henning; Farinella, Paolo; Scott, Edward R. D.; Keil, Klaus (1996). "Meteoritic, Asteroidal, and Theoretical Constraints on the 500 MA Disruption of the L Chondrite Parent Body". Icarus. 119 (1): 182–91. Bibcode:1996Icar..119..182H. doi:10.1006/icar.1996.0010.
- Korochantseva, Ekaterina V.; Trieloff, Mario; Lorenz, Cyrill A.; Buykin, Alexey I.; Ivanova, Marina A.; Schwarz, Winfried H.; Hopp, Jens; Jessberger, Elmar K. (2007). "L-chondrite asteroid breakup tied to Ordovician meteorite shower by multiple isochron 40Ar-39Ar dating". Meteoritics & Planetary Science. 42 (1): 113–30. Bibcode:2007M&PS...42..113K. doi:10.1111/j.1945-5100.2007.tb00221.x. S2CID 54513002.
- Jones, David S.; Brothers, R. William; Ahm, Anne-Sofie Crüger; Slater, Nicholas; Higgins, John A.; Fike, David A. (9 December 2019). "Sea level, carbonate mineralogy, and early diagenesis controlled δ13C records in Upper Ordovician carbonates". Geology. 48 (2): 194–199. doi:10.1130/G46861.1. S2CID 213408515.
- Stanley, S.; Hardie, L. (1998). "Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry". Palaeogeography, Palaeoclimatology, Palaeoecology. 144 (1–2): 3–19. Bibcode:1998PPP...144....3S. doi:10.1016/S0031-0182(98)00109-6.
- Stanley, S. M.; Hardie, L. A. (1999). "Hypercalcification; paleontology links plate tectonics and geochemistry to sedimentology". GSA Today. 9: 1–7.
- Munnecke, Axel; Calner, M.; Harper, David A. T.; Servais, Thomas (2010). "Ordovician and Silurian sea-water chemistry, sea level, and climate: A synopsis". Palaeogeography, Palaeoclimatology, Palaeoecology. 296 (3–4): 389–413. Bibcode:2010PPP...296..389M. doi:10.1016/j.palaeo.2010.08.001. Retrieved 16 August 2023.
- M. Marcilly, Chloé; Maffre, Pierre; Le Hir, Guillaume; Pohl, Alexandre; Fluteau, Frédéric; Goddéris, Yves; Donnadieu, Yannick; H. Heimdal, Thea; Torsvik, Trond H. (15 September 2022). "Understanding the early Paleozoic carbon cycle balance and climate change from modelling". Earth and Planetary Science Letters. 594: 117717. doi:10.1016/j.epsl.2022.117717. ISSN 0012-821X. Retrieved 17 September 2023.
- Bergmann, Kristin D.; Finnegan, Seth; Creel, Roger; Eiler, John M.; Hughes, Nigel C.; Popov, Leonid E.; Fischer, Woodward W. (1 March 2018). "A paired apatite and calcite clumped isotope thermometry approach to estimating Cambro-Ordovician seawater temperatures and isotopic composition". Geochimica et Cosmochimica Acta. 224: 18–41. Bibcode:2018GeCoA.224...18B. doi:10.1016/j.gca.2017.11.015.
- Elrick, Maya (1 October 2022). "Orbital-scale climate changes detected in Lower and Middle Ordovician cyclic limestones using oxygen isotopes of conodont apatite". Palaeogeography, Palaeoclimatology, Palaeoecology. 603: 111209. Bibcode:2022PPP...603k1209E. doi:10.1016/j.palaeo.2022.111209.
- Goldberg, Samuel L.; Present, Theodore M.; Finnegan, Seth; Bergmann, Kristin D. (2021-02-09). "A high-resolution record of early Paleozoic climate". Proceedings of the National Academy of Sciences of the United States of America. 118 (6): e2013083118. Bibcode:2021PNAS..11813083G. doi:10.1073/pnas.2013083118. ISSN 0027-8424. PMC 8017688. PMID 33526667.
- Vandenbroucke, Thijs R. A.; Armstrong, Howard A.; Williams, Mark; Paris, Florentin; Sabbe, Koen; Zalasiewicz, Jan A.; Nõlvak, Jaak; Verniers, Jacques (15 August 2010). "Epipelagic chitinozoan biotopes map a steep latitudinal temperature gradient for earliest Late Ordovician seas: Implications for a cooling Late Ordovician climate". Palaeogeography, Palaeoclimatology, Palaeoecology. 294 (3–4): 202–219. Bibcode:2010PPP...294..202V. doi:10.1016/j.palaeo.2009.11.026. Retrieved 29 December 2022.
- Rosenau, Nicholas A.; Hermann, Achim D.; Leslie, Stephen A. (15 January 2012). "Conodont apatite δ18O values from a platform margin setting, Oklahoma, USA: Implications for initiation of Late Ordovician icehouse conditions". Palaeogeography, Palaeoclimatology, Palaeoecology. 315–316: 172–180. Bibcode:2012PPP...315..172R. doi:10.1016/j.palaeo.2011.12.003. Retrieved 29 December 2022.
- Pohl, Alexandre; Donnadieu, Yannick; Le Hir, Guillaume; Ladant, Jean-Baptiste; Dumas, Christophe; Alvarez-Solas, Jorge; Vandenbroucke, Thijs R. A. (28 May 2016). "Glacial onset predated Late Ordovician climate cooling". Paleoceanography and Paleoclimatology. 31 (6): 800–821. Bibcode:2016PalOc..31..800P. doi:10.1002/2016PA002928. S2CID 133243759.
- Fortey, Richard A.; Cocks, L. Robin M. (2005). "Late Ordovician global warming—The Boda event". Geology. 33 (5): 405. Bibcode:2005Geo....33..405F. doi:10.1130/G21180.1.
- Trotter, J. A.; Williams, I. S.; Barnes, C. R.; Lecuyer, C.; Nicoll, R. S. (2008-07-25). "Did Cooling Oceans Trigger Ordovician Biodiversification? Evidence from Conodont Thermometry". Science. 321 (5888): 550–554. Bibcode:2008Sci...321..550T. doi:10.1126/science.1155814. ISSN 0036-8075. PMID 18653889. S2CID 28224399.
- Rasmussen, Christian M. Ø.; Ullmann, Clemens V.; Jakobsen, Kristian G.; Lindskog, Anders; Hansen, Jesper; Hansen, Thomas; Eriksson, Mats E.; Dronov, Andrei; Frei, Robert; Korte, Christoph; Nielsen, Arne T.; Harper, David A.T. (May 2016). "Onset of main Phanerozoic marine radiation sparked by emerging Mid Ordovician icehouse". Scientific Reports. 6 (1): 18884. Bibcode:2016NatSR...618884R. doi:10.1038/srep18884. PMC 4702064. PMID 26733399.
- "Humble moss helped to cool Earth and spurred on life". BBC News. 2 February 2012.
- Dixon, Dougal; et al. (2001). Atlas of Life on Earth. New York: Barnes & Noble Books. p. 87. ISBN 978-0-7607-1957-2.
- Palaeos Paleozoic : Ordovician : The Ordovician Period Archived 21 December 2007 at the Wayback Machine
- Cooper, John D.; Miller, Richard H.; Patterson, Jacqueline (1986). A Trip Through Time: Principles of Historical Geology. Columbus: Merrill Publishing Company. pp. 247, 255–259. ISBN 978-0-675-20140-7.
- Heim, Noel A. (8 April 2016). "A null biogeographic model for quantifying the role of migration in shaping patterns of global taxonomic richness and differentiation diversity, with implications for Ordovician biogeography". Paleobiology. 34 (2): 195–209. doi:10.1666/0094-8373(2008)034[0195:ANBMFQ]2.0.CO;2. Retrieved 18 May 2023.
- Cocks, L. Robin M.; Torsvik, Trond H. (December 2021). "Ordovician palaeogeography and climate change". Gondwana Research. 100: 53–72. doi:10.1016/j.gr.2020.09.008. Retrieved 17 September 2023.
- Cocks, L. R. M.; Fortey, R. A. (January 1990). "Biogeography of Ordovician and Silurian faunas". Geological Society, London, Memoirs. 12 (1): 97–104. doi:10.1144/GSL.MEM.1990.012.01.08. ISSN 0435-4052. Retrieved 17 September 2023.
- Fortey, Richard A.; Cocks, L.Robin M. (June 2003). "Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions". Earth-Science Reviews. 61 (3–4): 245–307. doi:10.1016/S0012-8252(02)00115-0. Retrieved 17 September 2023.
- Ghobadi Pour, M.; Popov, L. E.; Álvaro, J. J.; Amini, A.; Hairapetian, V.; Jahangir, H. (23 December 2022). "Ordovician of North Iran: New lithostratigraphy, palaeogeography and biogeographical links with South China and the Mediterranean peri-Gondwana margin". Bulletin of Geosciences. 97 (4): 465–538. Retrieved 17 September 2023.
- Burrett, Clive; Stait, Bryan (October 1985). "South East Asia as a part of an Ordovician Gondwanaland—a palaeobiogeographic test of a tectonic hypothesis". Earth and Planetary Science Letters. 75 (2–3): 184–190. doi:10.1016/0012-821X(85)90100-1. Retrieved 17 September 2023.
- Ebbestad, Jan Ove R.; Frýda, Jiří; Wagner, Peter J.; Horný, Radvan J.; Isakar, Mare; Stewart, Sarah; Percival, Ian G.; Bertero, Verònica; Rohr, David M.; Peel, John S.; Blodgett, Robert B.; Högström, Anette E. S. (November 2013). "Biogeography of Ordovician and Silurian gastropods, monoplacophorans and mimospirids". Geological Society, London, Memoirs. 38 (1): 199–220. doi:10.1144/M38.15. ISSN 0435-4052. Retrieved 17 September 2023.
- Harper, D.A.T.; Mac Niocaill, C.; Williams, S.H. (May 1996). "The palaeogeography of early Ordovician Iapetus terranes: an integration of faunal and palaeomagnetic constraints". Palaeogeography, Palaeoclimatology, Palaeoecology. 121 (3–4): 297–312. doi:10.1016/0031-0182(95)00079-8. Retrieved 17 September 2023.
- Penny, Amelia; Kröger, Björn (18 November 2019). "Impacts of spatial and environmental differentiation on early Palaeozoic marine biodiversity". Nature Ecology and Evolution. 3 (1): 1655–1660. doi:10.1038/s41559-019-1035-7. hdl:10138/325369. Retrieved 3 June 2023.
- Pedersen, R.B.; Bruton, D.L.; Furnes, H. (March 1992). "Ordovician faunas, island arcs and ophiolites in the Scandinavian Caledonides". Terra Nova. 4 (2): 217–222. doi:10.1111/j.1365-3121.1992.tb00475.x. ISSN 0954-4879. Retrieved 17 September 2023.
- Torsvik & Cocks 2017, p. 112-113.
- "Palaeos Paleozoic : Ordovician : The Ordovician Period". April 11, 2002. Archived from the original on December 21, 2007.
- "A Guide to the Orders of Trilobites".
- Garwood, Russell J.; Sharma, Prashant P.; Dunlop, Jason A.; Giribet, Gonzalo (5 May 2014). "A Paleozoic Stem Group to Mite Harvestmen Revealed through Integration of Phylogenetics and Development". Current Biology. 24 (9): 1017–1023. doi:10.1016/j.cub.2014.03.039. PMID 24726154.
- Taylor, P.D.; Berning, B.; Wilson, M.A. (2013). "Reinterpretation of the Cambrian 'bryozoan' Pywackia as an octocoral". Journal of Paleontology. 87 (6): 984–990. Bibcode:2013JPal...87..984T. doi:10.1666/13-029. S2CID 129113026.
- Bergström, Stig M.; Bergström, Jan; Kumpulainen, Risto; Ormö, Jens; Sturkell, Erik (2007). "Maurits Lindström – A renaissance geoscientist". GFF. 129 (2): 65–70. doi:10.1080/11035890701292065. S2CID 140593975.
- Song, Zhenyu; Xiao, Yunpeng; Xiao, Chuantao (19 February 2020). "Early–Middle Ordovician brachiopod diversification in the middle Yangtze region of South China". Canadian Journal of Earth Sciences. 57 (8): 999–1009. Bibcode:2020CaJES..57..999S. doi:10.1139/cjes-2019-0141. S2CID 213757467. Retrieved 22 November 2022.
- Harper, David A. T.; Zhan, Ren-Bin; Jin, Jisuo (March–June 2015). "The Great Ordovician Biodiversification Event: Reviewing two decades of research on diversity's big bang illustrated by mainly brachiopod data". Palaeoworld. 24 (1–2): 75–85. doi:10.1016/j.palwor.2015.03.003. Retrieved 12 November 2022.
- Zhan, Renbin; Rong, Jiayu; Cheng, Jinghui; Chen, Pengfei (May 2005). "Early-Mid Ordovician brachiopod diversification in South China". Science China Earth Sciences. 48 (5): 662–675. doi:10.1360/03yd0586. S2CID 130038222. Retrieved 12 November 2022.
- Patzkowsky, Mark E.; Holland, Steven M. (Fall 1997). "Patterns of turnover in Middle and Upper Ordovician brachiopods of the eastern United States: a test of coordinated stasis". Paleobiology. 23 (4): 420–443. doi:10.1017/S0094837300019825. Retrieved 23 July 2023.
- Novack-Gottshall, Philip M.; Miller, Arnold I. (Fall 2003). "Comparative geographic and environmental diversity dynamics of gastropods and bivalves during the Ordovician Radiation". Paleobiology. 29 (4): 576–604. doi:10.1666/0094-8373(2003)029<0576:CGAEDD>2.0.CO;2. S2CID 85724505. Retrieved 8 December 2022.
- Crick, Rex M. (Spring 1981). "Diversity and evolutionary rates of Cambro-Ordovician nautiloids". Paleobiology. 7 (2): 216–229. doi:10.1017/S0094837300003997. S2CID 83933056. Retrieved 8 December 2022.
- Kröger, Björn; Yun-Bai, Zhang (March 2009). "Pulsed cephalopod diversification during the Ordovician". Palaeogeography, Palaeoclimatology, Palaeoecology. 273 (1–2): 174–183. Bibcode:2009PPP...273..174K. doi:10.1016/j.palaeo.2008.12.015.
- Heward, A. P.; Fortey, R. A.; Miller, C. G.; Booth, G. A. (June 2023). "New Middle Ordovician (Darriwilian) faunas from the Sultanate of Oman". Proceedings of the Geologists' Association. 134 (3): 251–268. doi:10.1016/j.pgeola.2023.02.004. Retrieved 16 August 2023.
- Finney, Stanley C.; Bergström, Stig M. (1986). "Biostratigraphy of the Ordovician Nemagraptus gracilis Zone". Geological Society, London, Special Publications. 20 (1): 47–59. Bibcode:1986GSLSP..20...47F. doi:10.1144/GSL.SP.1986.020.01.06. S2CID 129733589.
- "12.7: Vertebrate Evolution". Biology LibreTexts. 2016-10-05. Retrieved 2022-06-07.
- Brazeau, M. D.; Friedman, M. (2015). "The origin and early phylogenetic history of jawed vertebrates". Nature. 520 (7548): 490–497. Bibcode:2015Natur.520..490B. doi:10.1038/nature14438. PMC 4648279. PMID 25903631.
- Nõlvak, Jaak; Liang, Yan; Hints, Olle (1 July 2019). "Early diversification of Ordovician chitinozoans on Baltica: New data from the Jägala waterfall section, northern Estonia". Palaeogeography, Palaeoclimatology, Palaeoecology. 525: 14–24. Bibcode:2019PPP...525...14N. doi:10.1016/j.palaeo.2019.04.002. S2CID 135138918. Retrieved 12 November 2022.
- Liang, Yan; Servais, Thomas; Tang, Peng; Lu, Jianbo; Wang, Wenhui (December 2017). "Tremadocian (Early Ordovician) chitinozoan biostratigraphy of South China: An update". Review of Palaeobotany and Palynology. 247: 149–163. doi:10.1016/j.revpalbo.2017.08.008. Retrieved 12 November 2022.
- Vinn, O.; Mõtus, M.-A. (2012). "Diverse early endobiotic coral symbiont assemblage from the Katian (Late Ordovician) of Baltica". Palaeogeography, Palaeoclimatology, Palaeoecology. 321–322: 137–141. Bibcode:2012PPP...321..137V. doi:10.1016/j.palaeo.2012.01.028. Retrieved 2014-06-11.
- Vinn, O.; Wilson, M.A.; Mõtus, M.-A.; Toom, U. (2014). "The earliest bryozoan parasite: Middle Ordovician (Darriwilian) of Osmussaar Island, Estonia". Palaeogeography, Palaeoclimatology, Palaeoecology. 414: 129–132. Bibcode:2014PPP...414..129V. doi:10.1016/j.palaeo.2014.08.021. Retrieved 2014-01-09.
- Ritchie, Alexander; Gilbert-Tomlinson, Joyce (24 November 1976). "First Ordovician vertebrates from the Southern Hemisphere". Alcheringa. 1 (4): 351–368. doi:10.1080/03115517708527770. Retrieved 12 November 2022.
- Wilson, M. A.; Palmer, T. J. (2006). "Patterns and processes in the Ordovician Bioerosion Revolution" (PDF). Ichnos. 13 (3): 109–112. doi:10.1080/10420940600850505. S2CID 128831144. Archived from the original (PDF) on 2008-12-16.
- Wilson, M. A.; Palmer, T. J. (2001). "Domiciles, not predatory borings: a simpler explanation of the holes in Ordovician shells analyzed by Kaplan and Baumiller, 2000". PALAIOS. 16 (5): 524–525. Bibcode:2001Palai..16..524W. doi:10.1669/0883-1351(2001)016<0524:DNPBAS>2.0.CO;2. S2CID 130036115.
- Porada, P.; Lenton, T. M.; Pohl, A.; Weber, B.; Mander, L.; Donnadieu, Y.; Beer, C.; Pöschl, U.; Kleidon, A. (November 2016). "High potential for weathering and climate effects of non-vascular vegetation in the Late Ordovician". Nature Communications. 7 (1): 12113. Bibcode:2016NatCo...712113P. doi:10.1038/ncomms12113. PMC 4941054. PMID 27385026.
- Steemans, P.; Herisse, A. L.; Melvin, J.; Miller, M. A.; Paris, F.; Verniers, J.; Wellman, C. H. (2009-04-17). "Origin and Radiation of the Earliest Vascular Land Plants". Science. 324 (5925): 353. Bibcode:2009Sci...324..353S. doi:10.1126/science.1169659. hdl:1854/LU-697223. PMID 19372423. S2CID 206518080.
- Redecker, D.; Kodner, R.; Graham, L. E. (2000). "Glomalean fungi from the Ordovician". Science. 289 (5486): 1920–1921. Bibcode:2000Sci...289.1920R. doi:10.1126/science.289.5486.1920. PMID 10988069. S2CID 43553633.
- Stanley, Steven M. (1999). Earth System History. New York: W.H. Freeman and Company. pp. 358, 360. ISBN 978-0-7167-2882-5.
- Young, Seth A.; Saltzman, Matthew R.; Ausich, William I.; Desrochers, André; Kaljo, Dimitri (2010). "Did changes in atmospheric CO2 coincide with latest Ordovician glacial–interglacial cycles?". Palaeogeography, Palaeoclimatology, Palaeoecology. 296 (3–4): 376–388. Bibcode:2010PPP...296..376Y. doi:10.1016/j.palaeo.2010.02.033.
- Jeff Hecht, High-carbon ice age mystery solved, New Scientist, 8 March 2010 (retrieved 30 June 2014)
- Emiliani, Cesare. (1992). Planet Earth : Cosmology, Geology, & the Evolution of Life & the Environment (Cambridge University Press) p. 491
- Torsvik & Cocks 2017, pp. 122–123.
- Melott, Adrian; et al. (2004). "Did a gamma-ray burst initiate the late Ordovician mass extinction?". International Journal of Astrobiology. 3 (1): 55–61. arXiv:astro-ph/0309415. Bibcode:2004IJAsB...3...55M. doi:10.1017/S1473550404001910. hdl:1808/9204. S2CID 13124815.
- Holland, Steven M; Patzkowsky, Mark E (2015). "The stratigraphy of mass extinction". Palaeontology. 58 (5): 903–924. doi:10.1111/pala.12188. S2CID 129522636.
External links
- Ogg, Jim (June 2004). "Overview of Global Boundary Stratotype Sections and Points (GSSP's)". Archived from the original on 2006-04-23. Retrieved 2006-04-30.
- Mehrtens, Charlotte. "Chazy Reef at Isle La Motte". An Ordovician reef in Vermont.
- Ordovician fossils of the famous Cincinnatian Group
- Ordovician (chronostratigraphy scale)