Palmitoylation
Palmitoylation is the covalent attachment of fatty acids, such as palmitic acid, to cysteine (S-palmitoylation) and less frequently to serine and threonine (O-palmitoylation) residues of proteins, which are typically membrane proteins.[2] The precise function of palmitoylation depends on the particular protein being considered. Palmitoylation enhances the hydrophobicity of proteins and contributes to their membrane association. Palmitoylation also appears to play a significant role in subcellular trafficking of proteins between membrane compartments,[3] as well as in modulating protein–protein interactions.[4] In contrast to prenylation and myristoylation, palmitoylation is usually reversible (because the bond between palmitic acid and protein is often a thioester bond). The reverse reaction in mammalian cells is catalyzed by acyl-protein thioesterases (APTs) in the cytosol and palmitoyl protein thioesterases in lysosomes. Because palmitoylation is a dynamic, post-translational process, it is believed to be employed by the cell to alter the subcellular localization, protein–protein interactions, or binding capacities of a protein.
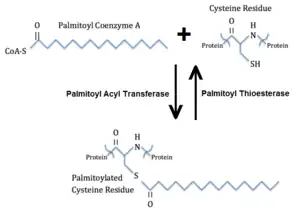
An example of a protein that undergoes palmitoylation is hemagglutinin, a membrane glycoprotein used by influenza to attach to host cell receptors.[5] The palmitoylation cycles of a wide array of enzymes have been characterized in the past few years, including H-Ras, Gsα, the β2-adrenergic receptor, and endothelial nitric oxide synthase (eNOS). In signal transduction via G protein, palmitoylation of the α subunit, prenylation of the γ subunit, and myristoylation is involved in tethering the G protein to the inner surface of the plasma membrane so that the G protein can interact with its receptor.[6]
Mechanism
S-palmitoylation is generally done by proteins with the DHHC domain. Exceptions exist in non-enzymatic reactions. Acyl-protein thioesterase (APT) catalyses the reverse reaction.[7] Other acyl groups such as stearate (C18:0) or oleate (C18:1) are also frequently accepted, more so in plant and viral proteins, making S-acylation a more useful name.[8][9]
Several structures of the DHHC domain have been determined using X-ray crystallography. It contains a linearly-arranged catalytic triad of Asp153, His154, and Cys156. It runs on a ping-pong mechanism, where the cysteine attacks the acyl-CoA to form an S-acylated DHHC, and then the acyl group is transferred to the substrate. DHHR enzymes exist, and it (as well as some DHHC enzymes) may use a ternary complex mechanism instead.[10]
An inhibitor of S-palmitoylation by DHHC is 2-Bromopalmitate (2-BP). 2-BP is a nonspecific inhibitor that also halts many other lipid-processing enzymes.[7]
The palmitoylome
A meta-analysis of 15 studies produced a compendium of approximately 2,000 mammalian proteins that are palmitoylated. The highest associations of the palmitoylome are with cancers and disorders of the nervous system. Approximately 40% of synaptic proteins were found in the palmitoylome.[11]
Biological function
Substrate presentation
Palmitoylation mediates the affinity of a protein for lipid rafts and facilitates the clustering of proteins.[12] The clustering can increase the proximity of two molecules. Alternatively, clustering can sequester a protein away from a substrate. For example, palmitoylation of phospholipase D (PLD) sequesters the enzyme away from its substrate phosphatidylcholine. When cholesterol levels decrease or PIP2 levels increase the palmitate mediated localization is disrupted, the enzyme trafficks to PIP2 where it encounters its substrate and is active by substrate presentation.[13][14][15]
Synapse formation
Scientists have appreciated the significance of attaching long hydrophobic chains to specific proteins in cell signaling pathways. A good example of its significance is in the clustering of proteins in the synapse. A major mediator of protein clustering in the synapse is the postsynaptic density (95kD) protein PSD-95. When this protein is palmitoylated it is restricted to the membrane. This restriction to the membrane allows it to bind to and cluster ion channels in the postsynaptic membrane. Also, in the presynaptic neuron, palmitoylation of SNAP-25 directs it to partition in the cell membrane [16] and allows the SNARE complex to dissociate during vesicle fusion. This provides a role for palmitoylation in regulating neurotransmitter release.[17]
Palmitoylation of delta catenin seems to coordinate activity-dependent changes in synaptic adhesion molecules, synapse structure, and receptor localizations that are involved in memory formation.[18]
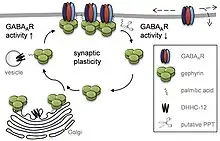
Palmitoylation of gephyrin has been reported to influence GABAergic synapses.[1]
References
- Dejanovic B, Semtner M, Ebert S, Lamkemeyer T, Neuser F, Lüscher B, Meier JC, Schwarz G (July 2014). "Palmitoylation of gephyrin controls receptor clustering and plasticity of GABAergic synapses". PLOS Biology. 12 (7): e1001908. doi:10.1371/journal.pbio.1001908. PMC 4099074. PMID 25025157.
- Linder, M.E., "Reversible modification of proteins with thioester-linked fatty acids," Protein Lipidation, F. Tamanoi and D.S. Sigman, eds., pp. 215-40 (San Diego, CA: Academic Press, 2000).
- Rocks O, Peyker A, Kahms M, Verveer PJ, Koerner C, Lumbierres M, Kuhlmann J, Waldmann H, Wittinghofer A, Bastiaens PI (2005). "An acylation cycle regulates localization and activity of palmitoylated Ras isoforms". Science. 307 (5716): 1746–1752. Bibcode:2005Sci...307.1746R. doi:10.1126/science.1105654. PMID 15705808. S2CID 12408991.
- Basu, J., "Protein palmitoylation and dynamic modulation of protein function," Current Science, Vol. 87, No. 2, pp. 212-17 (25 July 2004), http://www.ias.ac.in/currsci/jul252004/contents.htm
- Palese, Peter; García-Sastre, Adolfo (1999). "INFLUENZA VIRUSES (ORTHOMYXOVIRIDAE) | Molecular Biology". Encyclopedia of Virology. pp. 830–836. doi:10.1006/rwvi.1999.0157. ISBN 9780122270307. Archived from the original on 2012-09-12.
- Wall, MA; Coleman, DE; Lee, E; Iñiguez-Lluhi, JA; Posner, BA; Gilman, AG; Sprang, SR (Dec 15, 1995). "The structure of the G protein heterotrimer Gi alpha 1 beta 1 gamma 2". Cell. 83 (6): 1047–58. doi:10.1016/0092-8674(95)90220-1. PMID 8521505.
- Lanyon-Hogg, T., Faronato, M., Serwa, R. A., & Tate, E. W. (2017). Dynamic Protein Acylation: New Substrates, Mechanisms, and Drug Targets. Trends in Biochemical Sciences, 42(7), 566–581. doi:10.1016/j.tibs.2017.04.004
- Li, Y; Qi, B (2017). "Progress toward Understanding Protein S-acylation: Prospective in Plants". Frontiers in Plant Science. 8: 346. doi:10.3389/fpls.2017.00346. PMC 5364179. PMID 28392791.
- "Proteolipids - proteins modified by covalent attachment to lipids - N-myristoylated, S-palmitoylated, prenylated proteins, ghrelin, hedgehog proteins". www.lipidmaps.org.co.uk. Retrieved 19 July 2021.
- Rana, MS; Lee, CJ; Banerjee, A (28 February 2019). "The molecular mechanism of DHHC protein acyltransferases". Biochemical Society Transactions. 47 (1): 157–167. doi:10.1042/BST20180429. PMID 30559274. S2CID 56175691.
- Sanders SS, Martin DD, Butland SL, Lavallée-Adam M, Calzolari D, Kay C, Yates JR, Hayden MR (August 2015). "Curation of the Mammalian Palmitoylome Indicates a Pivotal Role for Palmitoylation in Diseases and Disorders of the Nervous System and Cancers". PLOS Computational Biology. 11 (8): e1004405. Bibcode:2015PLSCB..11E4405S. doi:10.1371/journal.pcbi.1004405. PMC 4537140. PMID 26275289.
- Levental, I.; Lingwood, D.; Grzybek, M.; Coskun, U.; Simons, K. (3 December 2010). "Palmitoylation regulates raft affinity for the majority of integral raft proteins". Proceedings of the National Academy of Sciences. 107 (51): 22050–22054. Bibcode:2010PNAS..10722050L. doi:10.1073/pnas.1016184107. PMC 3009825. PMID 21131568.
- Petersen, EN; Chung, HW; Nayebosadri, A; Hansen, SB (15 December 2016). "Kinetic disruption of lipid rafts is a mechanosensor for phospholipase D." Nature Communications. 7: 13873. Bibcode:2016NatCo...713873P. doi:10.1038/ncomms13873. PMC 5171650. PMID 27976674.
- Robinson, CV; Rohacs, T; Hansen, SB (September 2019). "Tools for Understanding Nanoscale Lipid Regulation of Ion Channels". Trends in Biochemical Sciences. 44 (9): 795–806. doi:10.1016/j.tibs.2019.04.001. PMC 6729126. PMID 31060927.
- Petersen, EN; Pavel, MA; Wang, H; Hansen, SB (28 October 2019). "Disruption of palmitate-mediated localization; a shared pathway of force and anesthetic activation of TREK-1 channels". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1862 (1): 183091. doi:10.1016/j.bbamem.2019.183091. PMC 6907892. PMID 31672538.
- Greaves, Jennifer (March 2011). "Differential palmitoylation regulates intracellular patterning of SNAP25". Journal of Cell Science. 124 (8): 1351–1360. doi:10.1242/jcs.079095. PMC 3065388. PMID 21429935.
- "Molecular Mechanisms of Synaptogenesis." Edited by Alexander Dityatev and Alaa El-Husseini. Springer: New York, NY. 2006. pg. 72-75
- Brigidi GS, Sun Y, Beccano-Kelly D, Pitman K, Jobasser M, Borgland SL, Milnerwood AJ, Bamji SX (January 23, 2014). "Palmitoylation of [delta]-catenin by DHHC5 mediates activity-induced synapse plasticity". Nature Neuroscience. 17 (4): 522–532. doi:10.1038/nn.3657. PMC 5025286. PMID 24562000.
Further reading
- Smotrys J, Linder A (2004). "Palmitoylation of Intracellular Signaling Proteins: Regulation and Function". Annu Rev Biochem. 73: 559–87. doi:10.1146/annurev.biochem.73.011303.073954. PMID 15189153.
- Resh, M. (2006) "Palmitoylation of Ligands, Receptors, and Intracellular Signaling Molecules". Sci STK. 359 October 31.
- Linder M, Deschenes R (2007). "Palmitoylation: policing protein stability and traffic". Nature Reviews Molecular Cell Biology. 8 (1): 74–84. doi:10.1038/nrm2084. PMID 17183362. S2CID 26339042.