Tetrachromacy
Tetrachromacy (from Greek tetra, meaning "four" and chroma, meaning "color") is the condition of possessing four independent channels for conveying color information, or possessing four types of cone cell in the eye. Organisms with tetrachromacy are called tetrachromats.
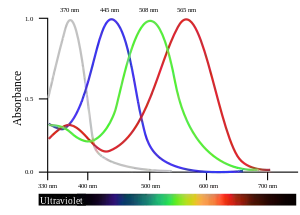
In tetrachromatic organisms, the sensory color space is four-dimensional, meaning that matching the sensory effect of arbitrarily chosen spectra of light within their visible spectrum requires mixtures of at least four primary colors.
Tetrachromacy is demonstrated among several species of birds,[2] fishes,[3] and reptiles.[3] The common ancestor of all vertebrates was a tetrachromat, but mammals evolved dichromacy, due to the nocturnal bottleneck, losing two of their four cones. Some primates then later evolved a third cone.[4]
Physiology
The normal explanation of tetrachromacy is that the organism's retina contains four types of higher-intensity light receptors (called cone cells in vertebrates as opposed to rod cells, which are lower-intensity light receptors) with different spectral sensitivity. This means that the organism may see wavelengths beyond those of a typical human's vision, and may be able to distinguish between colors that, to a normal human, appear to be identical. Species with tetrachromatic color vision may have an unknown physiological advantage over rival species.[5]
Humans
Apes (including humans) and Old World monkeys normally have three types of cone cell and are therefore trichromats. However, human tetrachromacy is suspected to exist in a small percentage of the population. Trichromats have 'red,' 'green', and 'blue' cone cells active in the long-, medium-, and short-wave part of the spectrum respectively, but at least one human has been identified with a well-separated fourth cone type.[6] Conclusively demonstrating that this extra cone type adds an additional subjective independent color dimension would confirm the existence of tetrachromacy among humans.
Tetrachromacy requires that there be four independent photoreceptor cell classes with different spectral sensitivity. However, there must also be the appropriate post-receptoral mechanism to compare the signals from the four classes of receptors. According to the opponent process theory, humans have three opponent channels, which grant trichromacy. Whether a fourth opponent channel is available to facilitate tetrachromacy is unclear.
Mice, which normally have only two cone pigments (and therefore two opponent channels), have been engineered to express a third cone pigment, and appear to demonstrate increased chromatic discrimination,[7] possibly indicating trichromacy and suggesting they were able to create or re-enable a third opponent channel. This would support the theory that humans should be able to utilize a fourth opponent channel for tetrachromatic vision. However, the original publication's claims about plasticity in the optic nerve have also been disputed.[8]
Tetrachromacy in carriers of CVD
It has been theorized that females who carry recessive opsin alleles that can cause color vision deficiency (CVD) could possess tetrachromacy. Female carriers of anomalous trichromacy (mild color blindness) possess heterozygous alleles of the genes that encode the L-opsin or M-opsin. These alleles often have a different spectral sensitivity, so if the carrier expresses both opsin alleles, they may exhibit tetrachromacy.
In humans, two cone cell pigment genes are present on the X chromosome: the classical type 2 opsin gene OPN1MW. People with two X chromosomes could possess multiple cone cell pigments, perhaps born as full tetrachromats who have four simultaneously-functioning kinds of cone cell, each type with a specific pattern of responsiveness to different wavelengths of light in the range of the visible spectrum.[9] One study suggested that 15% of the world's women might have the type of fourth cone whose sensitivity peak is between the standard red and green cones, giving, theoretically, a significant increase in color differentiation.[10] Another study suggests that as many as 50% of women and 8% of men may have four photopigments and corresponding increased chromatic discrimination compared to trichromats.[11] In 2010, after twenty years' study of women with four types of cones (non-functional tetrachromats), neuroscientist Gabriele Jordan identified a woman (subject cDa29) who could detect a greater variety of colors than trichromats could, corresponding with a functional or "true" tetrachromat.[6][12]
Variation in cone pigment genes is widespread in most human populations, but the most prevalent and pronounced tetrachromacy would derive from female carriers of major red/green pigment anomalies, usually classed as forms of "color blindness" (protanomaly or deuteranomaly). The biological basis for this phenomenon is X-inactivation of heterozygotic alleles for retinal pigment genes, which is the same mechanism that gives the majority of female New World monkeys trichromatic vision.[13]
In humans, preliminary visual processing occurs in the neurons of the retina. It is not known how these nerves would respond to a new color channel, that is, whether they could handle it separately or just combine it in with an existing channel. Similarly, visual information leaves the eye by way of the optic nerve, and a variety of final image processing takes place in the brain; it is not known whether the optic nerve or the areas of the brain have the spare capacity to handle or respond if presented with a new color channel.
Tetrachromacy may also enhance vision in dim lighting, or in looking at a screen.[14]
Conditional tetrachromacy
Despite being trichromats, humans can experience slight tetrachromacy at low light intensities, using their mesopic vision. In mesopic vision, both cone cells and rod cells are active. While rods typically don't contribute to color vision, they may in these specific light conditions, giving a small region of tetrachromacy in the color space.[15] Human rod cell sensitivity is greatest at 500 nm (bluish-green) wavelength, which is significantly different from the peak spectral sensitivity of the cones (typically 420, 530 and 560 nm).
Blocked tetrachromacy
Although many birds are tetrachromats with a fourth color in the ultraviolet, humans cannot see ultraviolet light directly because the lens of the eye blocks most light in the wavelength range of 300–400 nm; shorter wavelengths are blocked by the cornea.[16] The photoreceptor cells of the retina are sensitive to near ultraviolet light, and people lacking a lens (a condition known as aphakia) see near ultraviolet light (down to 300 nm) as whitish blue, or for some wavelengths, whitish violet, probably because all three types of cones are roughly equally sensitive to ultraviolet light (with blue cone cells slightly more sensitive).[17]
While an extended visible range does not denote tetrachromacy, some believe that visual pigments are available with sensitivity in near-UV wavelengths that would enable tetrachromacy in the case of aphakia.[18] However, there is no peer-reviewed evidence supporting this claim.
Other animals

Fish
Fish, specifically teleosts, are typically tetrachromats.[3] Exceptions include:
- Sharks and rays – range from monochromacy to trichromacy[3]
- Deep-sea fish – often rod monochromats
- Cichlid – arguably pentachromacy or higher[3]
Birds
Some species of birds, such as the zebra finch and the Columbidae, use the ultraviolet wavelength 300–400 nm specific to tetrachromatic color vision as a tool during mate selection and foraging.[19] When selecting for mates, ultraviolet plumage and skin coloration show a high level of selection.[20] A typical bird eye responds to wavelengths from about 300 to 700 nm. In terms of frequency, this corresponds to a band in the vicinity of 430–1000 THz. Most birds have retinas with four spectral types of cone cell that are believed to mediate tetrachromatic color vision. Bird color vision is further improved by filtering by pigmented oil droplets in the photoreceptors. The oil droplets filter incident light before it reaches the visual pigment in the outer segments of the photoreceptors.
The four cone types, and the specialization of pigmented oil droplets, give birds better color vision than that of humans.[21][22] However, more recent research has suggested that tetrachromacy in birds only provides birds with a larger visual spectrum than that in humans (humans cannot see ultraviolet light, 300-400 nm), while the spectral resolution (the "sensitivity" to nuances) is similar.[23]
Many diurnal birds of prey such as eagles, falcons and hawks have limited ability to perceive ultraviolet light, as this contributes heavily to chromatic aberration which reduces visual acuity.[24]
Pentachromacy and greater
The dimensionality of color vision has no upper bound, but vertebrates with color vision greater than tetrachromacy are rare. The next level is pentachromacy, which is five-dimensional color vision requiring at least 5 different classes of photoreceptor as well as 5 independent channels of color information through the primary visual system.
A female that is heterozygous for both the LWS and MWS opsins (and therefore a carrier for both protanomaly and deuteranomaly) would express five opsins of different spectral sensitivity. However, for her to be a true (strong) pentachromat, these opsins would need to be segregated into different photoreceptor cells and she would need to have the appropriate post-receptoral mechanisms to handle 5 opponent process channels, which is contentious.
Some birds (notably pigeons) have five or more kinds of color receptors in their retinae, and are therefore believed to be pentachromats, though psychophysical evidence of functional pentachromacy is lacking.[25] Research also indicates that some lampreys, members of the Petromyzontiformes, may be pentachromats.[26]
Invertebrates can have large numbers of different opsin classes, including 15 opsins in bluebottle butterflies[27] or 33 in mantis shrimp.[28] However, it has not been shown that color vision in these invertebrates is of a dimension commensurate with the number of opsins.
See also
References
- Figure data, uncorrected absorbance curve fits, from Hart, NS; Partridge, JC; Bennett, ATD; Cuthill, IC (2000). "Visual pigments, cone oil droplets and ocular media in four species of estrildid finch". Journal of Comparative Physiology A. 186 (7–8): 681–694. doi:10.1007/s003590000121. PMID 11016784. S2CID 19458550.
- Goldsmith, Timothy H. (2006). "What Birds See". Scientific American (July 2006): 69–75.
- Bowmaker, James K. (September 2008). "Evolution of vertebrate visual pigments". Vision Research. 48 (20): 2022–2041. doi:10.1016/j.visres.2008.03.025. PMID 18590925. S2CID 52808112.
- Jacobs, G. H. (2009). "Evolution of colour vision in mammals". Phil. Trans. R. Soc. B. 364 (1531): 2957–2967. doi:10.1098/rstb.2009.0039. PMC 2781854. PMID 19720656.
- Backhaus, W., Kliegl, R., Werner, J.S. (1998). "Color vision: perspective from different disciplines": 163–182.
{{cite journal}}: Cite journal requires|journal=(help)CS1 maint: multiple names: authors list (link) - Jordan, Gabriele; Deeb, Samir S.; Bosten, Jenny M.; Mollon, J. D. (July 2010). "The dimensionality of color vision in carriers of anomalous trichromacy". Journal of Vision. 10 (12): 12. doi:10.1167/10.8.12. PMID 20884587.
- Jacobs, Gerald H.; Williams, Gary A.; Cahill, Hugh; Nathans, Jeremy (23 March 2007). "Emergence of Novel Color Vision in Mice Engineered to Express a Human Cone Photopigment". Science. 315 (5819): 1723–1725. Bibcode:2007Sci...315.1723J. doi:10.1126/science.1138838. PMID 17379811. S2CID 85273369.
- Makous, W. (12 October 2007). "Comment on 'Emergence of Novel Color Vision in Mice Engineered to Express a Human Cone Photopigment'". Science. 318 (5848): 196. Bibcode:2007Sci...318..196M. doi:10.1126/science.1146084. PMID 17932271.
- Jameson, K. A., Highnote, S. M., & Wasserman, L. M. (2001). "Richer color experience in observers with multiple photopigment opsin genes" (PDF). Psychonomic Bulletin and Review. 8 (2): 244–261. doi:10.3758/BF03196159. PMID 11495112. S2CID 2389566. Archived from the original (PDF) on 14 February 2012.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Jordan, G. (July 1993). "A study of women heterozygous for colour deficiencies". Vision Research. Elsevier. 33 (11): 1495–1508. doi:10.1016/0042-6989(93)90143-K. PMID 8351822. S2CID 17648762.
- Backhaus, Werner G. K.; Backhaus, Werner; Kliegl, Reinhold; Werner, John Simon (1998). Color Vision: Perspectives from Different Disciplines. Walter de Gruyter. ISBN 9783110161007.
- San Diego woman Concetta Antico diagnosed with 'super vision' Published on 22 Nov 2013
- Richard C. Francis (2011). "Chapter 8. X-Women". Epigenetics: The Ultimate Mystery of Inheritance. New York and London: W. W. Norton. pp. 93–104. ISBN 978-0-393-07005-7.
- Robson, David (September 5, 2014). "The Women with Superhuman Vision". BBC News. Archived from the original on September 13, 2014. Retrieved December 30, 2017.
- Hansjochem Autrum & Richard Jung (1973). Integrative Functions and Comparative Data. Vol. 7. Springer-Verlag. p. 226. ISBN 978-0-387-05769-9.
- M A Mainster (2006). "Violet and blue light blocking intraocular lenses: photoprotection versus photoreception". British Journal of Ophthalmology. 90 (6): 784–792. doi:10.1136/bjo.2005.086553. PMC 1860240. PMID 16714268.
- Hambling, David (29 May 2002). "Let the light shine in". The Guardian.
- Fulton, James T. (31 July 2009). "THE HUMAN IS A BLOCKED TETRACHROMAT". Retrieved 1 June 2022.
- Bennett, Andrew T. D.; Cuthill, Innes C.; Partridge, Julian C.; Maier, Erhard J. (1996). "Ultraviolet vision and mate choice in zebra finches". Nature. 380 (6573): 433–435. Bibcode:1996Natur.380..433B. doi:10.1038/380433a0. S2CID 4347875.
- Bennett, Andrew T. D.; Théry, Marc (2007). "Avian Color Vision and Coloration: Multidisciplinary Evolutionary Biology" (PDF). The American Naturalist. 169 (S1): S1–S6. doi:10.1086/510163. ISSN 0003-0147. JSTOR 510163. S2CID 2484928.
- Cuthill, Innes C.; Partridge, Julian C.; Bennett, Andrew T. D.; Church, Stuart C.; Hart, Nathan S.; Hunt, Sarah (2000). J. B. Slater, Peter; Rosenblatt, Jay S.; Snowdon, Charles T.; Roper, Timothy J. (eds.). Ultraviolet Vision in Birds. p. 159. doi:10.1016/S0065-3454(08)60105-9. ISBN 978-0-12-004529-7.
{{cite book}}:|work=ignored (help) - Vorobyev, M. (November 1998). "Tetrachromacy, oil droplets and bird plumage colours". Journal of Comparative Physiology A. 183 (5): 621–33. doi:10.1007/s003590050286. PMID 9839454. S2CID 372159.
- Olsson, Peter; Lind, Olle; Kelber, Almut (2015-01-15). "Bird colour vision: behavioural thresholds reveal receptor noise". Journal of Experimental Biology. 218 (2): 184–193. doi:10.1242/jeb.111187. ISSN 0022-0949. PMID 25609782.
- Lind, Olle; Mitkus, Mindaugas; Olsson, Peter; Kelber, Almut (2013-05-15). "Ultraviolet sensitivity and colour vision in raptor foraging". journals.biologists.com. Retrieved 2023-06-13.
- Emmerton, Jacky; Delhis, Juan D. (1980). "Wavelength discrimination in the ?visible? and ultraviolet spectrum by pigeons". Journal of Comparative Physiology A. 141 (1): 47–52. doi:10.1007/BF00611877. S2CID 12462107.
- Davies, Wayne L.; Cowing, Jill A.; Carvalho, Livia S.; Potter, Ian C.; Trezise, Ann E. O.; Hunt, David M.; Collin, Shaun P. (September 2007). "Functional characterization, tuning, and regulation of visual pigment gene expression in an anadromous lamprey". The FASEB Journal. 21 (11): 2713–2724. doi:10.1096/fj.06-8057com. PMID 17463225. S2CID 5932623.
- Chen, Pei-Ju; Awata, Hiroko; Matsushita, Atsuko; Yang, En-Cheng; Arikawa, Kentaro (8 March 2016). "Extreme Spectral Richness in the Eye of the Common Bluebottle Butterfly, Graphium sarpedon". Frontiers in Ecology and Evolution. 4. doi:10.3389/fevo.2016.00018.
- Hansen, Sarah (17 July 2020). "Mantis Shrimp Eyes Get Even Wilder: UMBC Team Finds Twice The Expected Number Of Light-detecting Proteins - UMBC: University Of Maryland, Baltimore County". UMBC. Retrieved 7 October 2022.
External links
- Goldsmith, Timothy H. "What Birds See" Scientific American July 2006. An article about the tetrachromatic vision of birds
- Thompson, Evan (2000). "Comparative color vision: Quality space and visual ecology." In Steven Davis (Ed.), Color Perception: Philosophical, Psychological, Artistic and Computational Perspectives, pp. 163–186. Oxford: Oxford University Press.
- Looking for Madam Tetrachromat By Glenn Zorpette. Red Herring magazine, 1 November 2000
- "Exploring the fourth dimension". University of Bristol School of Biological Sciences. March 20, 2009.
- Colors - The Perfect Yellow By Radiolab, 21 May 2012 (Explores tetrachromacy in humans)
- The dimensionality of color vision in carriers of anomalous trichromacy--Gabriele Jordan et al--Journal of Vision August 12, 2010:
- On Tetrachromacy Ágnes Holba & B. Lukács
Color topics | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Color science |
|  | ||||||||
| Color philosophy |
| |||||||||
| Color terms |
| |||||||||
| Color organizations | ||||||||||
| Lists | ||||||||||
| Shades of: | ||||||||||
| Related | ||||||||||
| ||||||||||