Pyrococcus furiosus
Pyrococcus furiosus is a heterotrophic, strictly anaerobic, extremophilic, model species of archaea. It is classified as a hyperthermophile because it thrives best under extremely high temperatures, and is notable for having an optimum growth temperature of 100 °C (a temperature that would destroy most living organisms).[1] P. furiosus belongs to the Pyrococcus genus, most commonly found in extreme environmental conditions of hydrothermal vents. It is one of the few prokaryotic organisms that has enzymes containing tungsten, an element rarely found in biological molecules.
| Pyrococcus furiosus | |
|---|---|
 | |
| Pyrococcus furiosus | |
| Scientific classification | |
| Domain: | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | |
| Family: | |
| Genus: | |
| Species: | P. furiosus |
| Binomial name | |
| Pyrococcus furiosus Fiala and Stetter, 1986 | |
Pyrococcus furiosus has many potential industrial applications, owing to its unique thermostable properties. P. furiosus is used in the process of DNA amplification by way of polymerase chain reaction (PCR) because of its proofreading activity. Utilizing P. furiosus in PCR DNA amplification instead of the traditionally used Taq DNA polymerase allows for a significantly more accurate process.[2] The thermodynamic stability of P. furiosus' enzymes is useful in the creation of diols for laboratory and industrial purposes. Certain superoxide dismutases found in P. furiosus can be introduced into plants to increase their tolerance in environmentally stressful conditions and ultimately their survival.[3]
Properties
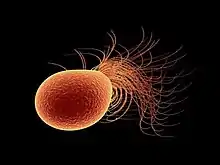
Pyrococcus furiosus is a strictly anaerobic, heterotrophic, sulfur-reducing archaea originally isolated from heated sediments in Vulcano, Italy by Fiala and Stetter. It is noted for its rapid doubling time of 37 minutes under optimal conditions, meaning that every 37 minutes the number of individual organisms is multiplied by two, yielding an exponential growth curve. Each organism is surrounded by a cellular envelope composed of glycoprotein called an S-layer. It appears as mostly regular cocci—meaning that it is roughly spherical—of 0.8 µm to 1.5 µm diameter with monopolar polytrichous flagellation.[1]
A glycoprotein notable to archaea species makes up the majority of the composition of P. furiosus flagella. Aside from potentially using them for swimming, these flagella were observed under lab conditions in use for unique applications such as forming cell to cell connections during stationary growth phase. They are additionally utilized as cable-like connectors to adhere to various solid surfaces such as sand grains in the habitat in which this species was discovered. This may lead to the formation of microcolonial biofilm-like structures.
P. furiosus grows between 70 °C (158 °F) and 103 °C (217 °F), with an optimum temperature of 100 °C (212 °F), and between pH 5 and 9 (with an optimum at pH 7). Since it uses fermentation of carbohydrates, it grows well on yeast extract, maltose, cellobiose, β-glucans, starch, and protein sources (tryptone, peptone, casein, and meat extracts) through the Embden-Meyerhoff pathway. This is a relatively wide range of sources when compared to other archaea. Growth is very slow, or nonexistent, on amino acids, organic acids, alcohols, and most carbohydrates (including glucose, fructose, lactose, and galactose). The metabolic products of P. furiosus are CO2 and H2. The presence of hydrogen severely inhibits its growth and metabolism; this effect can be circumvented, however, by introducing sulfur into the organism's environment. In this case, H2S can be produced through its metabolic processes seemingly for the purpose of detoxication or energy conservation, not energy production. While many other hyperthermophiles depend on sulfur for growth, P. furiosus does not.[4]
P. furiosus is also notable for an unusual and intriguingly simple respiratory system, which obtains energy by reducing protons to hydrogen gas and uses this energy to create an electrochemical gradient across its cell membrane, thereby driving ATP synthesis. This could be a very early evolutionary precursor of respiratory systems in all higher organisms today.[5]
Genomics
The sequencing of the complete genome of Pyrococcus furiosus was completed in 2001 by scientists at the University of Maryland Biotechnology Institute. The Maryland team found that the genome has 1,908 kilobases, including 2,065 open reading frames (ORFs) that encode proteins.[6] A study performed in 2005 revealed 17 new ORFs specific to Pyrococcus furiosus that were not originally annotated, bringing the number of ORFs up to 2,082.[7]
A lab strain of Pyrococcus furiosus named COM1 is commonly used for its "high plasticity" and ability to take up foreign DNA, owing to its high recombination and transposon activity. It has 1,571 more base pairs than the referenced NCBI genome, and 10 more insertion sequences (ISs). These ISs have deactivated 13 genes and many more are altered, but the strain's growth is yet comparable to its parent strain.[8]
Enzymes
Alcohol dehydrogenases
Pyrococcus furiosus possesses several highly thermostable alcohol dehydrogenases (ADHs): the short-chain AdhA, the iron-containing AdhB, the zinc-containing AdhC, and more.[9][10] Each of these ADHs are NADP-dependent, and serve to replenish NADP+ by using the NADPH produced by fermentation to reduce aldehydes to alcohols. The aldehydes are also products of fermentation and are toxic to the cell, so removal is necessary. P. furiosus ADHs typically have a broad range of aldehyde substrates they can use, and they can also catalyze the reverse reaction (oxidation of alcohols) using ethanol, 1,3-propanediol, and other alcohols for substrate. As with most of the archaea's enzymes, the ADHs are sensitive to oxygen.[11]
Oxidoreductases
Pyrococcus furiosus has five unique tungsten-containing oxidoreductases that are part of its NAD(P)H-independent glycolytic pathway. These enzymes function optimally above 90 °C. The first to be discovered was aldehyde ferredoxin oxidoreductase, or AOR, which utilizes tungsten, sulfur, and iron to catalyze the oxidation of aldehydes and reduce ferredoxin (this being the electron carrier instead of NAD(P)H).[12] As this was the first, all tungsten-containing oxidoreductases are said to be part of the AOR family. The next oxidoreductase to be discovered was glyceraldehyde-3-phosphate ferredoxin oxidoreductase, or GAPOR, which utilizes tungsten and iron to catalyze the oxidation of specifically glyceraldehyde-3-phosphate. GAPOR only functions under anaerobic conditions, as with many enzymes in P. furiosus.[13] Another oxidoreductase is formaldehyde ferredoxin oxidoreductase, or FOR, which catalyzes the oxidation of aldehydes into carboxylic acids. This enzyme utilizes four types of cofactors: tungsten, iron, sulfur, and calcium.[14] The next oxidoreductase, WOR4, does not help oxidize aldehydes, but rather has a role in the reduction of elemental sulfur (S0) into H2S. This uses the same cofactors as FOR, and is only found in P. furiosus cells that are grown in the presence of elemental sulfur.[15] The fifth and final oxidoreductase is named WOR5, and it has a broad specificity for aromatic and aliphatic aldehyde species.[16]
An oxidoreductase species in P. furiosus that does not contain tungsten is pyruvate ferredoxin oxidoreductase, or POR, which catalyzes the final step of the glycolytic pathway. It is possible that POR is an ancestor of mesophilic pyruvate oxidoreductases.[17] There is also the indolepyruvate ferredoxin oxidoreductase, or IOR, which utilizes iron and sulfur to catalyze the "oxidative decarboxylation of aryl pyruvates."[18]
Uses
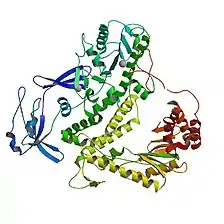
In DNA amplification
A DNA polymerase was discovered in P. furiosus that was thought to be unrelated to other known DNA polymerases, as no significant sequence homology was found between its two proteins and those of other known DNA polymerases. This DNA polymerase has strong 3'-to-5' exonucleolytic activity and a template-primer preference which is characteristic of a replicative DNA polymerase, leading scientists to believe that this enzyme may be the replicative DNA polymerase of P. furiosus.[19] It has since been placed in the family B of polymerases, the same family as DNA polymerase II. Its structure, which appears quite typical for polymerase B, has been solved as well.[20][21]
Since the enzymes of P. furiosus are extremely thermostable, the DNA polymerase from P. furiosus (also known as Pfu DNA polymerase) can be used in the polymerase chain reaction (PCR) DNA amplification process. The PCR process must use a thermostable DNA polymerase for automated in vitro amplification and originally used Taq DNA polymerase.[22] However, since purified Taq DNA polymerase lacks exonuclease (proofreading) activity, it cannot excise mismatched nucleotides. Researchers discovered in the early 1990's that the Pfu DNA polymerase of P. furiosus does actually possess a requisite 3’-to-5’ exonuclease activity allowing for the removal of errors. Subsequent tests utilizing Pfu DNA polymerase in the PCR process revealed a more than tenfold improvement over the accuracy of using Taq DNA polymerase.[2]
In production of diols
One practical application of P. furiosus is in the production of diols for various industrial processes. It may be possible to use the enzymes of P. furiosus for applications in such industries as food, pharmaceuticals, and fine-chemicals in which alcohol dehydrogenases are necessary in the production of enantio- and diastereomerically pure diols. Enzymes from hyperthermophiles such as P. furiosus can perform well in laboratory processes because they are relatively resistant: they generally function well at high temperatures and high pressures, as well as in high concentrations of chemicals.
In order to make naturally derived enzymes useful in the laboratory, it is often necessary to alter their genetic makeup. Otherwise, the naturally occurring enzymes may not be efficient in an artificially induced procedure. Although the enzymes of P. furiosus function optimally at a high temperature, scientists may not necessarily want to carry out a procedure at 100 °C (212 °F). Consequently, in this case, the specific enzyme AdhA was taken from P. furiosus and put through various mutations in a laboratory in order to obtain a suitable alcohol dehydrogenase for use in artificial processes. This allowed scientists to obtain a mutant enzyme that could function efficiently at lower temperatures and maintain productivity.[23]
In plants
The expression of a certain gene found in P. furiosus in plants can also render them more durable by increasing their tolerance for heat. In response to environmental stresses such as heat exposure, plants produce reactive oxygen species which can result in cell death. If these free radicals are removed, cell death can be delayed. Enzymes in plants called superoxide dismutases remove superoxide anion radicals from cells, but increasing the amount and activity of these enzymes is difficult and not the most efficient way to go about improving the durability of plants.[24]
By introducing the superoxide reductases of P. furiosus into plants, the levels of O2 can be rapidly reduced. Scientists tested this method using the Arabidopsis thaliana plant. As a result of this procedure, cell death in plants occurs less often, therefore resulting in a reduction in the severity of responses to environmental stress. This enhances the survival of plants, making them more resistant to light, chemical, and heat stress.
This study could potentially be used as a starting point to creating plants that could survive in more extreme climates on other planets such as Mars. By introducing more enzymes from extremophiles like P. furiosus into other species of plants, it may be possible to create incredibly resistant species.[3]
In researching amino acids
By comparing P. furiosus with a related species of archaea, Pyrococcus abyssi, scientists have tried to determine the correlation between certain amino acids and affinity for certain pressures in different species. P. furiosus is not barophilic, while P. abyssi is, meaning that it functions optimally at very high pressures. Using two hyperthermophilic species of archaea lessens the possibility of deviations having to do with temperature of the environment, essentially reducing the variables in the experimental design.[25]
Besides yielding information about the barophilicity of certain amino acids, the experiment also provided valuable insight into the origin of the genetic code and its organizational influences. It was found that most of the amino acids that determined barophilicity were also found to be important in the organization of the genetic code. It was also found that more polar amino acids and smaller amino acids were more likely to be barophilic. Through the comparison of these two archaea, the conclusion was reached that the genetic code was likely structured under high hydrostatic pressure, and that hydrostatic pressure was a more influential factor in determining genetic code than temperature.[25]
History
Pyrococcus furiosus was originally isolated anaerobically from geothermal marine sediments with temperatures between 90 °C (194 °F) and 100 °C (212 °F) collected at the beach of Porto Levante, Vulcano Island, Italy. It was first described by Karl Stetter of the University of Regensburg in Germany, and a colleague, Gerhard Fiala. Pyrococcus furiosus actually originated a new genus of archaea with its relatively recent discovery in 1986.[1]
The name Pyrococcus means "fireball" in Greek, to refer to the extremophile's round shape and ability to grow in temperatures of around 100 degrees Celsius. The species name furiosus means 'rushing' in Latin, and refers to the extremophile's doubling time and rapid swimming.[1]
References
- Fiala, Gerhard; Stetter, Karl O (1986). "Pyrococcus furiosus sp. nov. Represents a novel genus of marine heterotrophic archaebacteria growing optimally at 100°C". Archives of Microbiology. 145 (1): 56–61. doi:10.1007/BF00413027. S2CID 41589578.
- Lundberg, Kelly S.; Shoemaker, Dan D.; Adams, Michael W.W.; Short, Jay M.; Sorge, Joseph A.; Mathur, Eric J. (December 1991). "High-fidelity amplification using a thermostable DNA polymerase isolated from Pyrococcus furiosus". Gene. 108 (1): 1–6. doi:10.1016/0378-1119(91)90480-y. ISSN 0378-1119. PMID 1761218.
- Karen Miller (August 5, 2005). "Prozac for Plants". National Space Science Data Center. NASA. Archived from the original on August 8, 2005.
- Silva, Pedro J.; Ban, Eyke C. D. van den; Wassink, Hans; Haaker, Huub; Castro, Baltazar de; Robb, Frank T.; Hagen, Wilfred R. (2000). "Enzymes of hydrogen metabolism in Pyrococcus furiosus". European Journal of Biochemistry. 267 (22): 6541–6551. doi:10.1046/j.1432-1327.2000.01745.x. ISSN 0014-2956. PMID 11054105.
- Sapra, R; Bagramyan, K; Adams, M. W. W (2003). "A simple energy-conserving system: Proton reduction coupled to proton translocation". Proceedings of the National Academy of Sciences. 100 (13): 7545–50. Bibcode:2003PNAS..100.7545S. doi:10.1073/pnas.1331436100. PMC 164623. PMID 12792025.
- Robb, Frank T; Maeder, Dennis L; Brown, James R; DiRuggiero, Jocelyne; Stump, Mark D; Yeh, Raymond K; Weiss, Robert B; Dunn, Dianne M (2001), Genomic sequence of hyperthermophile, Pyrococcus furiosus: Implications for physiology and enzymology, Methods in Enzymology, vol. 330, Elsevier, pp. 134–157, doi:10.1016/s0076-6879(01)30372-5, ISBN 9780121822316, PMID 11210495, retrieved 2022-10-06
- W., Poole, Farris L. Gerwe, Brian A. Hopkins, Robert C. Schut, Gerrit J. Weinberg, Michael V. Jenney, Francis E. Adams, Michael W. Defining Genes in the Genome of the Hyperthermophilic Archaeon Pyrococcus furiosus: Implications for All Microbial Genomes†. American Society for Microbiology. OCLC 678564723.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Bridger, Stephanie L.; Lancaster, W. Andrew; Poole, Farris L.; Schut, Gerrit J.; Adams, Michael W. W. (August 2012). "Genome Sequencing of a Genetically Tractable Pyrococcus furiosus Strain Reveals a Highly Dynamic Genome". Journal of Bacteriology. 194 (15): 4097–4106. doi:10.1128/jb.00439-12. ISSN 0021-9193. PMC 3416535. PMID 22636780.
- van der Oost, John; Voorhorst, Wilfried G. B.; Kengen, Servé W. M.; Geerling, Ans C. M.; Wittenhorst, Vincent; Gueguen, Yannick; de Vos, Willem M. (2001-05-15). "Genetic and biochemical characterization of a short-chain alcohol dehydrogenase from the hyperthermophilic archaeonPyrococcus furiosus". European Journal of Biochemistry. 268 (10): 3062–3068. doi:10.1046/j.1432-1327.2001.02201.x. ISSN 0014-2956. PMID 11358525.
- Kube, Jürgen; Brokamp, Christian; Machielsen, Ronnie; van der Oost, John; Märkl, Herbert (2006-02-07). "Influence of temperature on the production of an archaeal thermoactive alcohol dehydrogenase from Pyrococcus furiosus with recombinant Escherichia coli". Extremophiles. 10 (3): 221–227. doi:10.1007/s00792-005-0490-z. ISSN 1431-0651. PMID 16463078. S2CID 28865345.
- Ma, Kesen, Adams, Michael W. W. (15 February 1999). "An Unusual Oxygen-Sensitive, Iron- and Zinc-Containing Alcohol Dehydrogenase from the Hyperthermophilic Archaeon Pyrococcus furiosus". Journal of Bacteriology. 181 (4): 1163–1170. doi:10.1128/JB.181.4.1163-1170.1999. PMC 93493. PMID 9973342.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Makund, S.; Adams, M.W.W. (August 1991). "The novel tungsten-iron-sulfur protein of the hyperthermophilic archaebacterium, Pyrococcus furiosus, is an aldehyde ferredoxin oxidoreductase: Evidence for its participation in a unique glycolytic pathway". Journal of Inorganic Biochemistry. 43 (2–3): 257. doi:10.1016/0162-0134(91)84247-7. ISSN 0162-0134.
- Mukund, Swarnalatha; Adams, Michael W.W. (April 1995). "Glyceraldehyde-3-phosphate Ferredoxin Oxidoreductase, a Novel Tungsten-containing Enzyme with a Potential Glycolytic Role in the Hyperthermophilic Archaeon Pyrococcus furiosus". Journal of Biological Chemistry. 270 (15): 8389–8392. doi:10.1074/jbc.270.15.8389. ISSN 0021-9258. PMID 7721730.
- Hu, Yonglin; Faham, Salem; Roy, Roopali; Adams, Michael W.W; Rees, Douglas C (February 1999). "Formaldehyde ferredoxin oxidoreductase from Pyrococcus furiosus: the 1.85 Å resolution crystal structure and its mechanistic implications 1 1Edited by I. A. Wilson". Journal of Molecular Biology. 286 (3): 899–914. doi:10.1006/jmbi.1998.2488. PMID 10024458.
- Roy, Roopali; Adams, Michael W. W. (2002-12-15). "Characterization of a Fourth Tungsten-Containing Enzyme from the Hyperthermophilic Archaeon Pyrococcus furiosus". Journal of Bacteriology. 184 (24): 6952–6956. doi:10.1128/JB.184.24.6952-6956.2002. ISSN 0021-9193. PMC 135473. PMID 12446645.
- Bevers, Loes E.; Bol, Emile; Hagedoorn, Peter-Leon; Hagen, Wilfred R. (2005-10-15). "WOR5, a Novel Tungsten-Containing Aldehyde Oxidoreductase from Pyrococcus furiosus with a Broad Substrate Specificity". Journal of Bacteriology. 187 (20): 7056–7061. doi:10.1128/JB.187.20.7056-7061.2005. ISSN 0021-9193. PMC 1251609. PMID 16199576.
- Blamey, Jenny M.; Adams, Michael W.W. (January 1993). "Purification and characterization of pyruvate ferredoxin oxidoreductase from the hyperthermophilic archaeon Pyrococcus furiosus". Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology. 1161 (1): 19–27. doi:10.1016/0167-4838(93)90190-3. PMID 8380721.
- Mai, X.; Adams, M.W. (June 1994). "Indolepyruvate ferredoxin oxidoreductase from the hyperthermophilic archaeon Pyrococcus furiosus. A new enzyme involved in peptide fermentation". Journal of Biological Chemistry. 269 (24): 16726–16732. doi:10.1016/S0021-9258(19)89451-6. PMID 8206994.
- Uemori, Takashi; Sato, Yoshimi; Kato, Ikunoshin; Doi, Hirofumi; Ishino, Yoshizumi (1997). "A novel DNA polymerase in the hyperthermophilic archaeon,Pyrococcus furiosus: Gene cloning, expression, and characterization". Genes to Cells. 2 (8): 499–512. doi:10.1046/j.1365-2443.1997.1380336.x. PMID 9348040.
- Ishino, S; Ishino, Y (2014). "DNA polymerases as useful reagents for biotechnology - the history of developmental research in the field". Frontiers in Microbiology. 5: 465. doi:10.3389/fmicb.2014.00465. PMC 4148896. PMID 25221550.
- Kim, Suhng Wook; Kim, Dong-Uk; Kim, Jin Kwang; Kang, Lin-Woo; Cho, Hyun-Soo (May 2008). "Crystal structure of Pfu, the high fidelity DNA polymerase from Pyrococcus furiosus". International Journal of Biological Macromolecules. 42 (4): 356–361. doi:10.1016/j.ijbiomac.2008.01.010. PMID 18355915.
- Saiki, RK; Gelfand, DH; Stoffel, S; Scharf, SJ; Higuchi, R; Horn, GT; Mullis, KB; Erlich, HA (1988-01-29). "Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase". Science. 239 (4839): 487–491. doi:10.1126/science.239.4839.487. ISSN 0036-8075. PMID 2448875.
- Machielsen, Ronnie; Leferink, Nicole G. H; Hendriks, Annemarie; Brouns, Stan J. J; Hennemann, Hans-Georg; Dauβmann, Thomas; Van Der Oost, John (2008). "Laboratory evolution of Pyrococcus furiosus alcohol dehydrogenase to improve the production of (2S,5S)-hexanediol at moderate temperatures". Extremophiles. 12 (4): 587–94. doi:10.1007/s00792-008-0164-8. PMC 2467505. PMID 18452026.
- Im, Y. J; Ji, M; Lee, A; Killens, R; Grunden, A. M; Boss, W. F (2009). "Expression of Pyrococcus furiosus Superoxide Reductase in Arabidopsis Enhances Heat Tolerance". Plant Physiology. 151 (2): 893–904. doi:10.1104/pp.109.145409. PMC 2754621. PMID 19684226.
- Di Giulio, Massimo (2005). "A comparison of proteins from Pyrococcus furiosus and Pyrococcus abyssi: Barophily in the physicochemical properties of amino acids and in the genetic code". Gene. 346: 1–6. doi:10.1016/j.gene.2004.10.008. PMID 15716096.
Further reading
- McTernan, Patrick M; Chandrayan, Sanjeev K; Wu, Chang-Hao; Vaccaro, Brian J; Lancaster, W. Andrew; Yang, Qingyuan; Fu, Dax; Hura, Greg L; Tainer, John A; Adams, Michael W. W (2014). "Intact Functional Fourteen-subunit Respiratory Membrane-bound [Ni Fe]-Hydrogenase Complex of the Hyperthermophilic Archaeon Pyrococcus furiosus". Journal of Biological Chemistry. 289 (28): 19364–72. doi:10.1074/jbc.M114.567255. PMC 4094048. PMID 24860091.
- Dong, Qing; Yan, Xufan; Zheng, Minhui; Yang, Ziwen (2014). "Characterization of an extremely thermostable but cold-adaptive β-galactosidase from the hyperthermophilic archaeon Pyrococcus furiosus for use as a recombinant aggregation for batch lactose degradation at high temperature". Journal of Bioscience and Bioengineering. 117 (6): 706–10. doi:10.1016/j.jbiosc.2013.12.002. PMID 24462527.
- Elshawadfy, Ashraf M; Keith, Brian J; Ee Ooi, H'Ng; Kinsman, Thomas; Heslop, Pauline; Connolly, Bernard A (2014). "DNA polymerase hybrids derived from the family-B enzymes of Pyrococcus furiosus and Thermococcus kodakarensis: Improving performance in the polymerase chain reaction". Frontiers in Microbiology. 5: 224. doi:10.3389/fmicb.2014.00224. PMC 4034419. PMID 24904539.
- Herzog, B; Wirth, R (2012). "Swimming Behavior of Selected Species of Archaea". Applied and Environmental Microbiology. 78 (6): 1670–4. doi:10.1128/AEM.06723-11. PMC 3298134. PMID 22247169.