Multiregional origin of modern humans
The multiregional hypothesis, multiregional evolution (MRE), or polycentric hypothesis, is a scientific model that provides an alternative explanation to the more widely accepted "Out of Africa" model of monogenesis for the pattern of human evolution.
Multiregional evolution holds that the human species first arose around two million years ago and subsequent human evolution has been within a single, continuous human species. This species encompasses all archaic human forms such as H. erectus and Neanderthals as well as modern forms, and evolved worldwide to the diverse populations of anatomically modern humans (Homo sapiens).
The hypothesis contends that the mechanism of clinal variation through a model of "Centre and Edge" allowed for the necessary balance between genetic drift, gene flow and selection throughout the Pleistocene, as well as overall evolution as a global species, but while retaining regional differences in certain morphological features.[1] Proponents of multiregionalism point to fossil and genomic data and continuity of archaeological cultures as support for their hypothesis.
The multiregional hypothesis was first proposed in 1984, and then revised in 2003. In its revised form, it is similar to the Assimilation Model, which holds that modern humans originated in Africa and today share a predominant recent African origin, but have also absorbed small, geographically variable, degrees of admixture from other regional (archaic) hominin species.[2]
The multiregional hypothesis is not currently the most accepted theory of modern human origin among scientists. "The African replacement model has gained the widest acceptance owing mainly to genetic data (particularly mitochondrial DNA) from existing populations. This model is consistent with the realization that modern humans cannot be classified into subspecies or races, and it recognizes that all populations of present-day humans share the same potential."[3] The African replacement model is also known as the "Out of Africa" theory. See also, "The 'out of Africa' model is currently the most widely accepted model. It proposes that Homo sapiens evolved in Africa before migrating across the world."[4] And: "The primary competing scientific hypothesis is currently recent African origin of modern humans, which proposes that modern humans arose as a new species in Africa around 100-200,000 years ago, moving out of Africa around 50-60,000 years ago to replace existing human species such as Homo erectus and the Neanderthals without interbreeding. [5] [6] [7] [8] This differs from the multiregional hypothesis in that the multiregional model predicts interbreeding with preexisting local human populations in any such migration." [9] [10]
History
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Overview
The Multiregional hypothesis was proposed in 1984 by Milford H. Wolpoff, Alan Thorne and Xinzhi Wu.[11][12][1] Wolpoff credits Franz Weidenreich's "Polycentric" hypothesis of human origins as a major influence, but cautions that this should not be confused with polygenism, or Carleton Coon's model that minimized gene flow.[12][13][14] According to Wolpoff, multiregionalism was misinterpreted by William W. Howells, who confused Weidenreich's hypothesis with a polygenic "candelabra model" in his publications spanning five decades:
How did Multiregional evolution get stigmatized as polygeny? We believe it comes from the confusion of Weidenreich's ideas, and ultimately of our own, with Coon's. The historic reason for linking Coon's and Weidenreich's ideas came from the mischaracterizations of Weidenreich's Polycentric model as a candelabra (Howells, 1942, 1944, 1959, 1993), that made his Polycentric model appear much more similar to Coon's than it actually was.[15]
Through the influence of Howells, many other anthropologists and biologists have confused multiregionalism with polygenism i.e. separate or multiple origins for different populations. Alan Templeton for example notes that this confusion has led to the error that gene flow between different populations was added to the Multiregional hypothesis as a "special pleading in response to recent difficulties", despite the fact: "parallel evolution was never part of the multiregional model, much less its core, whereas gene flow was not a recent addition, but rather was present in the model from the very beginning"[16] (emphasis in original). Despite this, multiregionalism is still confused with polygenism, or Coon's model of racial origins, from which Wolpoff and his colleagues have distanced themselves.[17][18] Wolpoff has also defended Wiedenreich's Polycentric hypothesis from being labeled polyphyletic. Weidenreich himself in 1949 wrote: "I may run the risk of being misunderstood, namely that I believe in polyphyletic evolution of man".[19]
In 1998, Wu founded a China-specific Multiregional model called "Continuity with [Incidental] Hybridization".[20][21] Wu's variant only applies the Multiregional hypothesis to the East Asian fossil record, and is popular among Chinese scientists.[22] However, James Leibold, a political historian of modern China, has argued the support for Wu's model is largely rooted in Chinese nationalism.[23] Outside of China, the Multiregional hypothesis has limited support, held only by a small number of paleoanthropologists.[24]
"Classic" vs "weak" multiregionalism
Chris Stringer, a leading proponent of the more mainstream recent African origin theory, debated Multiregionalists such as Wolpoff and Thorne in a series of publications throughout the late 1980s and 1990s.[25][26][27][28] Stringer describes how he considers the original Multiregional hypothesis to have been modified over time into a weaker variant that now allows a much greater role for Africa in human evolution, including anatomical modernity (and subsequently less regional continuity than was first proposed).[29]
Stringer distinguishes the original or "classic" Multiregional model as having existed from 1984 (its formulation) until 2003, to a "weak" post-2003 variant that has "shifted close to that of the Assimilation Model".[30][31]
Genetic studies
The finding that "Mitochondrial Eve" was relatively recent and African seemed to give the upper hand to the proponents of the Out of Africa hypothesis. But in 2002, Alan Templeton published a genetic analysis involving other loci in the genome as well, and this showed that some variants that are present in modern populations existed already in Asia hundreds of thousands of years ago.[32] This meant that even if our male line (Y chromosome) and our female line (mitochondrial DNA) came out of Africa in the last 100,000 years or so, we have inherited other genes from populations that were already outside of Africa. Since this study other studies have been done using much more data (see Phylogeography).
Fossil evidence
Morphological clades


Proponents of the multiregional hypothesis see regional continuity of certain morphological traits spanning the Pleistocene in different regions across the globe as evidence against a single replacement model from Africa. In general, three major regions are recognized: Europe, China, and Indonesia (often including Australia).[33][34][35] Wolpoff cautions that the continuity in certain skeletal features in these regions should not be seen in a racial context, instead calling them morphological clades; defined as sets of traits that "uniquely characterise a geographic region".[36] According to Wolpoff and Thorne (1981): "We do not regard a morphological clade as a unique lineage, nor do we believe it necessary to imply a particular taxonomic status for it".[37] Critics of multiregionalism have pointed out that no single human trait is unique to a geographical region (i.e. confined to one population and not found in any other) but Wolpoff et al. (2000) note that regional continuity only recognizes combinations of features, not traits if individually accessed, a point they elsewhere compare to the forensic identification of a human skeleton:
Regional continuity... is not the claim that such features do not appear elsewhere; the genetic structure of the human species makes such a possibility unlikely to the extreme. There may be uniqueness in combinations of traits, but no single trait is likely to have been unique in a particular part of the world although it might appear to be so because of the incomplete sampling provided by the spotty human fossil record.
Combinations of features are "unique" in the sense of being found in only one region, or more weakly limited to one region at high frequency (very rarely in another). Wolpoff stresses that regional continuity works in conjunction with genetic exchanges between populations. Long-term regional continuity in certain morphological traits is explained by Alan Thorne's "Centre and Edge"[38] population genetics model which resolves Weidenreich's paradox of "how did populations retain geographical distinctions and yet evolve together?". For example, in 2001 Wolpoff and colleagues published an analysis of character traits of the skulls of early modern human fossils in Australia and central Europe. They concluded that the diversity of these recent humans could not "result exclusively from a single late Pleistocene dispersal", and implied dual ancestry for each region, involving interbreeding with Africans.[39]
Indonesia, Australia
Thorne held that there was regional continuity in Indonesia and Australia for a morphological clade.[40][41] This sequence is said to consist of the earliest fossils from Sangiran, Java, that can be traced through Ngandong and found in prehistoric and recent Aboriginal Australians. In 1991, Andrew Kramer tested 17 proposed morphological clade features. He found that: "a plurality (eight) of the seventeen non-metric features link Sangiran to modern Australians" and that these "are suggestive of morphological continuity, which implies the presence of a genetic continuum in Australasia dating back at least one million years"[42] but Colin Groves has criticized Kramer's methodology, pointing out that the polarity of characters was not tested and that the study is actually inconclusive.[43] Dr. Phillip Habgood discovered that the characters said to be unique to the Australasian region by Thorne are plesiomorphic:
...it is evident that all of the characters proposed... to be 'clade features' linking Indonesian Homo erectus material with Australian Aboriginal crania are retained primitive features present on Homo erectus and archaic Homo sapiens crania in general. Many are also commonly found on the crania and mandibles of anatomically-modern Homo sapiens from other geographical locations, being especially prevalent on the robust Mesolithic skeletal material from North Africa."[44]
Yet, regardless of these criticisms Habgood (2003) allows for limited regional continuity in Indonesia and Australia, recognizing four plesiomorphic features which do not appear in such a unique combination on fossils in any other region: a sagittally flat frontal bone, with a posterior position of minimum frontal breadth, great facial prognathism, and zygomaxillary tuberosities.[45] This combination, Habgood says, has a "certain Australianness about it".
Wolpoff, initially skeptical of Thorne's claims, became convinced when reconstructing the Sangiran 17 Homo erectus skull from Indonesia, when he was surprised that the skull's face to vault angle matched that of the Australian modern human Kow Swamp 1 skull in excessive prognathism. Durband (2007) in contrast states that "features cited as showing continuity between Sangiran 17 and the Kow Swamp sample disappeared in the new, more orthognathic reconstruction of that fossil that was recently completed".[46] Baba et al. who newly restored the face of Sangiran 17 concluded: "regional continuity in Australasia is far less evident than Thorne and Wolpoff argued".[47]
China
_presented_at_Paleozoological_Museum_of_China.jpg.webp)
Xinzhi Wu has argued for a morphological clade in China spanning the Pleistocene, characterized by a combination of 10 features.[48][49] The sequence is said to start with Lantian and Peking Man, traced to Dali, to Late Pleistocene specimens (e.g. Liujiang) and recent Chinese. Habgood in 1992 criticized Wu's list, pointing out that most of the 10 features in combination appear regularly on fossils outside China.[50] He did though note that three combined: a non-depressed nasal root, non-projecting perpendicularly oriented nasal bones and facial flatness are unique to the Chinese region in the fossil record and may be evidence for limited regional continuity. However, according to Chris Stringer, Habgood's study suffered from not including enough fossil samples from North Africa, many of which exhibit the small combination he considered to be region-specific to China.[28]
Facial flatness as a morphological clade feature has been rejected by many anthropologists since it is found on many early African Homo erectus fossils, and is therefore considered plesiomorphic,[51] but Wu has responded that the form of facial flatness in the Chinese fossil record appears distinct to other (i.e. primitive) forms. Toetik Koesbardiati in her PhD thesis "On the Relevance of the Regional Continuity Features of the Face in East Asia" also found that a form of facial flatness is unique to China (i.e. only appears there at high frequency, very rarely elsewhere) but cautions that this is the only available evidence for regional continuity: "Only two features appear to show a tendency as suggested by the Multiregional model: flatness at the upper face expressed by an obtuse nasio-frontal angle and flatness at the middle part of the face expressed by an obtuse zygomaxillay angle".
Shovel-shaped incisors are commonly cited as evidence for regional continuity in China.[52][53] Stringer (1992) however found that shovel-shaped incisors are present on >70% of the early Holocene Wadi Halfa fossil sample from North Africa, and common elsewhere.[54] Frayer et al. (1993) have criticized Stringer's method of scoring shovel-shaped incisor teeth. They discuss the fact that there are different degrees of "shovelled" e.g. trace (+), semi (++), and marked (+++), but that Stringer misleadingly lumped all these together: "...combining shoveling categories in this manner is biologically meaningless and misleading, as the statistic cannot be validly compared with the very high frequencies for the marked shoveling category reported for East Asians."[34] Palaeoanthropologist Fred H. Smith (2009) also emphasizes that: "It is the pattern of shoveling that identities as an East Asian regional feature, not just the occurrence of shoveling of any sort".[2] Multiregionalists argue that marked (+++) shovel-shaped incisors only appear in China at a high frequency, and have <10% occurrence elsewhere.
Europe
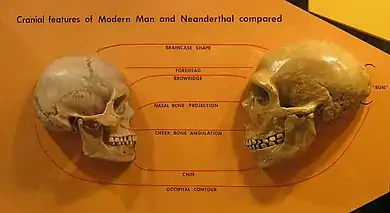
Since the early 1990s, David W. Frayer has described what he regards as a morphological clade in Europe.[55][56][57] The sequence starts with the earliest dated Neanderthal specimens (Krapina and Saccopastore skulls) traced through the mid-Late Pleistocene (e.g. La Ferrassie 1) to Vindija Cave, and late Upper Palaeolithic Cro-Magnons or recent Europeans. Although many anthropologists consider Neanderthals and Cro Magnons morphologically distinct,[58][59] Frayer maintains quite the opposite and points to their similarities, which he argues is evidence for regional continuity:
"Contrary to Brauer's recent pronouncement that there is a large and generally recognized morphological gap between the Neanderthals and the early moderns, the actual evidence provided by the extensive fossil record of late Pleistocene Europe shows considerable continuity between Neanderthals and subsequent Europeans."[34]
Frayer et al. (1993) consider there to be at least four features in combination that are unique to the European fossil record: a horizontal-oval shaped mandibular foramen, anterior mastoid tubercle, suprainiac fossa and narrowing of the nasal breadth associated with tooth-size reduction. Regarding the latter, Frayer observes a sequence of nasal narrowing in Neanderthals, following through to late Upper Palaeolithic and Holocene (Mesolithic) crania. His claims are disputed by others,[60] but have received support from Wolpoff, who regards late Neanderthal specimens to be "transitional" in nasal form between earlier Neanderthals and later Cro Magnons.[61] Based on other cranial similarities, Wolpoff et al. (2004) argue for a sizable Neanderthal contribution to modern Europeans.[62]
More recent claims regarding continuity in skeletal morphology in Europe focus on fossils with both Neanderthal and modern anatomical traits, to provide evidence of interbreeding rather than replacement.[63][64][65] Examples include the Lapedo child found in Portugal[66] and the Oase 1 mandible from Peștera cu Oase, Romania,[67] though the Lapedo child is disputed by some.[68]
Genetic evidence
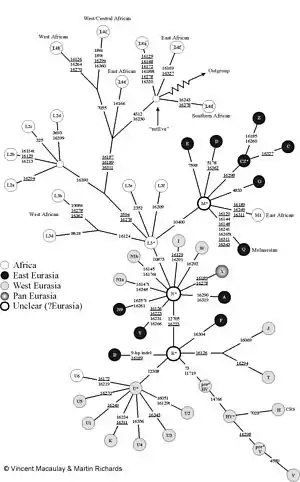
Mitochondrial Eve
A 1987 analysis of mitochondrial DNA from 147 people by Cann et al. from around the world indicated that their mitochondrial lineages all coalesced in a common ancestor from Africa between 140,000 and 290,000 years ago.[69] The analysis suggested that this reflected the worldwide expansion of modern humans as a new species, replacing, rather than mixing with, local archaic humans outside of Africa. Such a recent replacement scenario is not compatible with the Multiregional hypothesis and the mtDNA results led to increased popularity for the alternative single replacement theory.[70][71][72] According to Wolpoff and colleagues:
"When they were first published, the Mitochondrial Eve results were clearly incongruous with Multiregional evolution, and we wondered how the two could be reconciled."[73]
Multiregionalists have responded to what they see as flaws in the Eve theory,[74] and have offered contrary genetic evidences.[75][76][77] Wu and Thorne have questioned the reliability of the molecular clock used to date Eve.[78][79] Multiregionalists point out that Mitochondrial DNA alone can not rule out interbreeding between early modern and archaic humans, since archaic human mitochondrial strains from such interbreeding could have been lost due to genetic drift or a selective sweep.[80][81] Wolpoff for example states that Eve is "not the most recent common ancestor of all living people" since "Mitochondrial history is not population history".[82]
Neanderthal mtDNA
Neanderthal mitochondrial DNA (mtDNA) sequences from Feldhofer and Vindija Cave are substantially different from modern human mtDNA.[83][84][85] Multiregionalists however have discussed the fact that the average difference between the Feldhofer sequence and living humans is less than that found between chimpanzee subspecies,[86][87] and therefore that while Neanderthals were different subspecies, they were still human and part of the same lineage.
Nuclear DNA
Initial analysis of Y chromosome DNA, which like mitochondrial DNA, is inherited from only one parent, was consistent with a recent African replacement model. However, the mitochondrial and Y chromosome data could not be explained by the same modern human expansion out of Africa; the Y chromosome expansion would have involved genetic mixing that retained regionally local mitochondrial lines. In addition, the Y chromosome data indicated a later expansion back into Africa from Asia, demonstrating that gene flow between regions was not unidirectional.[88]
An early analysis of 15 noncoding sites on the X chromosome found additional inconsistencies with the recent African replacement hypothesis. The analysis found a multimodal distribution of coalescence times to the most recent common ancestor for those sites, contrary to the predictions for recent African replacement; in particular, there were more coalescence times near 2 million years ago (mya) than expected, suggesting an ancient population split around the time humans first emerged from Africa as Homo erectus, rather than more recently as suggested by the mitochondrial data. While most of these X chromosome sites showed greater diversity in Africa, consistent with African origins, a few of the sites showed greater diversity in Asia rather than Africa. For four of the 15 gene sites that did show greater diversity in Africa, the sites' varying diversity by region could not be explained by simple expansion from Africa, as would be required by the recent African replacement hypothesis.[89]
Later analyses of X chromosome and autosomal DNA continued to find sites with deep coalescence times inconsistent with a single origin of modern humans,[90][91][92][93][94] diversity patterns inconsistent with a recent expansion from Africa,[95] or both.[96][97] For example, analyses of a region of RRM2P4 (ribonucleotide reductase M2 subunit pseudogene 4) showed a coalescence time of about 2 Mya, with a clear root in Asia,[98][99] while the MAPT locus at 17q21.31 is split into two deep genetic lineages, one of which is common in and largely confined to the present European population, suggesting inheritance from Neanderthals.[100][101][102][103] In the case of the Microcephalin D allele, evidence for rapid recent expansion indicated introgression from an archaic population.[104][105][106][107] However, later analysis, including of the genomes of Neanderthals, did not find the Microcephalin D allele (in the proposed archaic species), nor evidence that it had introgressed from an archaic lineage as previously suggested.[108][109][110]
In 2001, a DNA study of more than 12,000 men from 163 East Asian regions showed that all of them carry a mutation that originated in Africa about 35,000 to 89,000 years ago and these "data do not support even a minimal in situ hominid contribution in the origin of anatomically modern humans in East Asia".[111]
In a 2005 review and analysis of the genetic lineages of 25 chromosomal regions, Alan Templeton found evidence of more than 34 occurrences of gene flow between Africa and Eurasia. Of these occurrences, 19 were associated with continuous restricted gene exchange through at least 1.46 million years ago; only 5 were associated with a recent expansion from Africa to Eurasia. Three were associated with the original expansion of Homo erectus out of Africa around 2 million years ago, 7 with an intermediate expansion out of Africa at a date consistent with the expansion of Acheulean tool technology, and a few others with other gene flows such as an expansion out of Eurasia and back into Africa subsequent to the most recent expansion out of Africa. Templeton rejected a hypothesis of complete recent African replacement with greater than 99% certainty (p < 10−17).[112]
Ancient DNA
Recent analyses of DNA taken directly from Neanderthal specimens indicates that they or their ancestors contributed to the genome of all humans outside of Africa, indicating there was some degree of interbreeding with Neanderthals before their replacement.[113] It has also been shown that Denisova hominins contributed to the DNA of Melanesians and Australians through interbreeding.[114]
By 2006, extraction of DNA directly from some archaic human samples was becoming possible. The earliest analyses were of Neanderthal DNA, and indicated that the Neanderthal contribution to modern human genetic diversity was no more than 20%, with a most likely value of 0%.[115] By 2010, however, detailed DNA sequencing of the Neanderthal specimens from Europe indicated that the contribution was nonzero, with Neanderthals sharing 1-4% more genetic variants with living non-Africans than with living humans in sub-Saharan Africa.[116][117] In late 2010, a recently discovered non-Neanderthal archaic human, the Denisova hominin from south-western Siberia, was found to share 4–6% more of its genome with living Melanesian humans than with any other living group, supporting admixture between two regions outside of Africa.[118][119] In August 2011, human leukocyte antigen (HLA) alleles from the archaic Denisovan and Neanderthal genomes were found to show patterns in the modern human population demonstrating origins from these non-African populations; the ancestry from these archaic alleles at the HLA-A site was more than 50% for modern Europeans, 70% for Asians, and 95% for Papua New Guineans.[120] Proponents of the multiregional hypothesis believe the combination of regional continuity inside and outside of Africa and lateral gene transfer between various regions around the world supports the multiregional hypothesis. However, "Out of Africa" Theory proponents also explain this with the fact that genetic changes occur on a regional basis rather than a continental basis, and populations close to each other are likely to share certain specific regional SNPs while sharing most other genes in common.[121][122] Migration Matrix theory (A=Mt) indicates that dependent upon the potential contribution of Neanderthal ancestry, we would be able to calculate the percentage of Neanderthal mtDNA contribution to the human species. As we do not know the specific migration matrix, we are unable to input the exact data, which would answer these questions irrefutably.[86]
See also
References
- Wolpoff, M. H.; Spuhler, J. N.; Smith, F. H.; Radovcic, J.; Pope, G.; Frayer, D. W.; Eckhardt, R.; Clark, G. (1988). "Modern Human Origins". Science. 241 (4867): 772–74. Bibcode:1988Sci...241..772W. doi:10.1126/science.3136545. PMID 3136545. S2CID 5223638.
- Cartmill, M & Smith, F. H. (2009). The Human Lineage. Wiley-Blackwell. p. 450.
- https://www.britannica.com/science/human-evolution/The-emergence-of-Homo-sapiens
- https://www.yourgenome.org/stories/evolution-of-modern-humans/#:~:text=The%20'out%20of%20Africa'%20model%20is%20currently%20the%20most%20widely,a%20long%20period%20of%20time.
- Hua Liu, et al. (2006). "A Geographically Explicit Genetic Model of Worldwide Human-Settlement History". American Journal of Human Genetics 79 (2): 230–237. doi:10.1086/505436. PMID 16826514. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1559480
- Weaver, Timothy D; Charles C. Roseman (2008). "New developments in the genetic evidence for modern human origins". Evolutionary Anthropology: Issues, News, and Reviews (Wiley-Liss) 17 (1): 69–80. doi:10.1002/evan.20161. http://www3.interscience.wiley.com/journal/117921411/abstract.
- Fagundes, NJ; Ray N, Beaumont M, Neuenschwander S, Salzano FM, Bonatto SL, Excoffier L. (2007). "Statistical evaluation of alternatihttps://encyclopedia.pub/entry/33820ve models of human evolution". Proc Natl Acad Sci USA 104 (45): 17614–9. doi:10.1073/pnas.0708280104. PMID 17978179. PMC 2077041. Bibcode: 2007PNAS..10417614F. http://www.pnas.org/content/104/45/17614.long.
- Wolpoff, Milford, and Caspari, Rachel (1997). Race and Human Evolution. Simon & Schuster. p. 42.
- Wolpoff, Milford, and Caspari, Rachel (1997). Race and Human Evolution. Simon & Schuster. p. 42.
- Scholarly Community Encyclopeda.
- Wolpoff, M. H., Wu, X. Z., & Alan, G. (86). G. Thorne: 1984, "Modern Homo Sapiens Origins: A General Theory of Hominid Evolution Involving the Fossil Evidence from east Asia". The Origins of Modern Humans, Liss, New York, 411–83.
- Wolpoff, MH; Hawks, J; Caspari, R (2000). "Multiregional, not multiple origins" (PDF). American Journal of Physical Anthropology. 112 (1): 129–36. doi:10.1002/(SICI)1096-8644(200005)112:1<129::AID-AJPA11>3.0.CO;2-K. hdl:2027.42/34270. PMID 10766948.
- Hawks, J.; Wolpoff, M. H. (2003). "Sixty years of modern human origins in the American Anthropological Association" (PDF). American Anthropologist. 105 (1): 89–100. doi:10.1525/aa.2003.105.1.89. hdl:2027.42/65197.
- Eckhardt, R. B.; Wolpoff, M. H.; Thorne, A. G. (1993). "Multiregional Evolution". Science. 262 (5136): 974. doi:10.1126/science.262.5136.973-b.
- Caspari, R.; Wolpoff, M. H. (1996). "Weidenreich, Coon, and multiregional evolution". Human Evolution. 11 (3–4): 261–68. doi:10.1007/bf02436629. S2CID 84805412.
- Templeton, A. R. (2007). "Genetics and recent human evolution". Evolution. 61 (7): 1507–19. doi:10.1111/j.1558-5646.2007.00164.x. PMID 17598736.
- Wolpoff, M. H. and R. Caspari. 1997. Race and human evolution: A fatal attraction. New York: Simon and Schuster.
- Wolpoff, M.H., and R. Caspari. (2000). "The Many Species of Humanity". Przegląd Antropologiczny (Anthropological Review) 63(1): 3–17.
- Weidenreich, F. (1949). "Interpretations of the fossil material". In: Early Man in the Far East: Studies in Physical Anthropology. Howells, W. W (ed). Studies in Physical Anthropology; No. 1, American Association of Physical Anthropologists, Detroit. pp. 149–57.
- Wu, X. (1998). "Origin of modern humans of China viewed from cranio-dental characteristics of late Homo sapiens". Acta Anthropologica Sinica. 17: 276–82.
- Rosenberg, K. R., Wu, X. (2013). "A River Runs through It: Modern Human Origins in East Asia". In: The Origins of Modern Humans: Biology Reconsidered. Smith, F. H (ed). Wiley-Blackwell. pp. 89–122.
- Liu, L., Chen, X. (2012). The Archaeology of China: From the Late Paleolithic to the Early Bronze Age. Cambridge University Press. p. 14: "the majority of Chinese archaeologists and palaeontologists support[s] the multiregional development model, proposing a hypothesis of regional continuity with hybridization between immigrants and indigenous populations in the evolution from H. erectus to H. sapiens in East Asia."
- Leibold, J. (2012). "'Filling in the Nation: The Spatial Trajectory of Prehistoric Archaeology in Twentieth-Century China," in Transforming History: The Making of a Modern Academic Discipline in Twentieth Century China, eds. Brian Moloughney and Peter Zarrow, pp. 333–71 (Hong Kong: Chinese University Press).
- Begun, DR. (2013). "The Past, Present and Future of Palaeoanthropology". In: A Companion to Paleoanthropology. Wiley-Blackwell. p. 8: "It needs to be noted, however, that this [Multiregional model] is a minority view among paleoanthropologists, most of whom support the African replacement model."
- Stringer, C. B.; Andrews, P. (1988). "Genetic and fossil evidence for the origin of modern humans". Science. 239 (4845): 1263–68. Bibcode:1988Sci...239.1263S. doi:10.1126/science.3125610. PMID 3125610.
- Stringer, C.; Bräuer, G. (1994). "Methods, misreading, and bias". American Anthropologist. 96 (2): 416–24. doi:10.1525/aa.1994.96.2.02a00080.
- Stringer, C. B. (1992). "Replacement, continuity and the origin of Homo sapiens". In: Continuity or replacement? Controversies in Homo sapiens evolution. F. H. Smith (ed). Rotterdam: Balkema. pp. 9–24.
- Bräuer, G., & Stringer, C. (1997). "Models, polarization, and perspectives on modern human origins". In: Conceptual issues in modern human origins research. New York: Aldine de Gruyter. pp. 191–201.
- Stringer, C. (2001). "Modern human origins – distinguishing the models". Afr. Archaeol. Rev. 18 (2): 67–75. doi:10.1023/A:1011079908461. S2CID 161991922.
- Stringer, C (2002). "Modern human origins: progress and prospects". Philosophical Transactions of the Royal Society of London. 357 (1420): 563–79. doi:10.1098/rstb.2001.1057. PMC 1692961. PMID 12028792.
- Stringer, C. (2014). "Why we are not all multiregionalists now". Trends in Ecology & Evolution. 29 (5): 248–51. doi:10.1016/j.tree.2014.03.001. PMID 24702983.
- Templeton, A. R. (2002). "Out of Africa again and again" (PDF). Nature. 416 (6876): 45–51. Bibcode:2002Natur.416...45T. doi:10.1038/416045a. PMID 11882887. S2CID 4397398.
- Wolpoff, M. H. (1985). Human evolution at the peripheries: the pattern at the eastern edge. Hominid Evolution: past, present and future, 355–65.
- Frayer, D.W.; Wolpoff, M. H.; Thorne, A.G.; Smith, F. H.; Pope, G.G. (1993). "Theories of modern human origins: the paleontological test". American Anthropologist. 95 (1): 14–50. doi:10.1525/aa.1993.95.1.02a00020.
- Wolpoff, M.H., A.G. Thorne, F.H. Smith, D.W. Frayer, and G.G. Pope: Multiregional Evolution: A World-Wide Source for Modern Human Populations. In: Origins of Anatomically Modern Humans, edited by M.H. Nitecki and D.V. Nitecki. Plenum Press, New York. pp. 175–99.
- Wolpoff, M. H. (1989). "Multiregional evolution: the fossil alternative to Eden". The Human Revolution: Behavioural and Biological Perspectives on the Origins of Modern Humans. 1: 62–108.
- Thorne, A. G.; Wolpoff, M. H. (1981). "Regional continuity in Australasian Pleistocene hominid evolution". American Journal of Physical Anthropology. 55 (3): 337–49. doi:10.1002/ajpa.1330550308. PMID 6791505.
- Thorne, A.G. (1981). "The Centre and the Edge: The significance of Australian hominids to African Palaeoanthropology". Proceedings of the 8th Pan-African Congress of Prehistory (Nairobi), pp. 180–81. Nairobi: National Museums of Kenya.
- Wolpoff, Milford H.; John Hawks; David W. Frayer; Keith Hunley (2001). "Modern Human Ancestry at the Peripheries: A Test of the Replacement Theory". Science. 291 (5502): 293–97. Bibcode:2001Sci...291..293W. doi:10.1126/science.291.5502.293. PMID 11209077.
- Thorne, A.G. (1984). "Australia's human origins – how many sources?". American Journal of Physical Anthropology. 63 (2): 133–242. doi:10.1002/ajpa.1330630203. PMID 6711682.
- Thorne, A.G.; Wolpoff, M.H. (1992). "The multiregional evolution of humans". Scientific American. 266 (4): 76–83. Bibcode:1992SciAm.266d..76T. doi:10.1038/scientificamerican0492-76. PMID 1566033.
- Kramer, A. (1991). "Modern human origins in Australasia: replacement or evolution?". American Journal of Physical Anthropology. 86 (4): 455–73. doi:10.1002/ajpa.1330860403. PMID 1776654.
- Groves, C.P. (1997). "Thinking about Evolutionary Change The Polarity of Our Ancestors". In: Conceptual issues in modern human origins research. Transaction Publishers.
- Habgood, P.J. (1989) "The origin of anatomically modern humans in Australasia. In: P. Mellars and C.B. Stringer (eds.) The Human Revolution: Behavioural and Biological Perspectives in the Origins of Modern Humans, pp. 245–73.
- Habgood, P.J. (2003). A Morphometric Investigation into the Origins of Anatomically Modern Humans. British Archaeological Reports, International Series 1176. Oxford England: Archaeopress.
- Durband, A. (2007). "The view from down under: a test of the multiregional hypothesis of modern human origins using the basicranial evidence from Australasia". Collegium Antropologicum. 31 (3): 651–59. PMID 18041369.
- Baba, H.; Aziz, F.; Narasaki, S. (2000). "Restoration of the face of Javanese Homo erectus Sangiran 17 and re-evaluation of regional continuity in Australasia". Acta Anthropologica Sinica. 19: 34–40.
- Wu, X. (1990). "The evolution of humankind in China". Acta Anthropologica Sinica. 9 (4): 312–21.
- Wu, X., Poirier, F. E. (1995).Human evolution in China: a metric description of the fossils and a review of the sites. New York: Oxford University Press.
- Habgood, P.J. (1992). "The origin of anatomically modern humans in east Asia". In: G. Bräuer, and F.H. Smith (eds.) Continuity or Replacement: Controversies in Homo sapiens evolution. pp. 273–88.
- Groves, C. P. (1989). "A regional approach to the problem of the origin of modern humans in Australasia". In: P. Mellars & CB. Stringer (eds). In: The Human Revolution. Princeton: Princeton University Press. pp. 274–85.
- Woo, R (1986). "Chinese human fossils and the origin of Mongoloid racial group". Anthropos (Brno). 23: 151–55.
- Wu, X. (2006). "Evidence of Multiregional Human Evolution Hypothesis from China". Quaternary Sciences. 26 (5): 702–70.
- Stringer, C. B. (1992). "Replacement, continuity and the origin of Homo sapiens". In: Continuity or replacement? Controversies in Homo sapiens evolution. Rotterdam: A. A. Balkema. pp. 9–24.
- Frayer, D. W. (1992). "The persistence of Neanderthal features in post-Neanderthal Europeans". In: Continuity or Replacement: Controversies in Homo sapiens Evolution. Rotterdam: Balkema, pp. 179–88.
- Frayer, D. W. (1992)."Evolution at the European edge: Neanderthal and the Upper Paleolithic relationships. Préhistoire Européene. 2:9–69.
- Frayer, D. W. (1997)."Perspectives on Neanderthals as ancestors". In: Conceptual Issues in Modern Human Origins Research. New York: Aldine de Gruyter. pp. 220–35.
- Harvati, Katerina; Stephen R. Frost; Kieran P McNulty (2004). "Neanderthal taxonomy reconsidered: Implications of 3D primate models of intra- and interspecific differences". PNAS. 101 (5): 1147–52. Bibcode:2004PNAS..101.1147H. doi:10.1073/pnas.0308085100. PMC 337021. PMID 14745010.
- Pearson, Osbjorn M. (2004). "Has the Combination of Genetic and Fossil Evidence Solved the Riddle of Modern Human Origins?". Evolutionary Anthropology. 13 (4): 145–59. doi:10.1002/evan.20017. S2CID 31478877.
- Holton, N. E.; Franciscus, R. G. (2008). "The paradox of a wide nasal aperture in cold-adapted Neandertals: a causal assessment". Journal of Human Evolution. 55 (6): 942–51. doi:10.1016/j.jhevol.2008.07.001. PMID 18842288.
- Wolpoff, M. H. (1989). "The place of Neanderthals in human evolution". In: The Emergence of Modern Humans: Biocultural Adaptations in the Later Pleistocene. Trinkaus, E (ed.). Cambridge University Press. pp. 97–41.
- Wolpoff, Milford; Bruce Mannheim; Alan Mann; John Hawks; Rachel Caspari; Karen R. Rosenberg; David W. Frayer; George W. Gill; Geoffrey Clark (2004). "Why not the Neandertals?". World Archaeology. 36 (4): 527–46. doi:10.1080/0043824042000303700. S2CID 2507757.
- Trinkaus, E. (May 2007). "European early modern humans and the fate of the Neandertals". Proceedings of the National Academy of Sciences of the United States of America. 104 (18): 7367–72. Bibcode:2007PNAS..104.7367T. doi:10.1073/pnas.0702214104. ISSN 0027-8424. PMC 1863481. PMID 17452632.
- The Emerging Fate Of The Neandertals
- Smith, F. H.; Janković, I.; Karavanić, I. (2005). "The assimilation model, modern human origins in Europe, and the extinction of Neandertals". Quaternary International. 137 (1): 7–19. Bibcode:2005QuInt.137....7S. doi:10.1016/j.quaint.2004.11.016.
- Duarte C.; Maurício J.; Pettitt P.; Souto P.; Trinkaus E.; van der Plicht H.; Zilhão J. (1999). "The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia". Proc Natl Acad Sci USA. 96 (13): 7604–09. Bibcode:1999PNAS...96.7604D. doi:10.1073/pnas.96.13.7604. PMC 22133. PMID 10377462.
- Trinkaus, E.; Moldovan, O.; Milota, S.; Bîlgăr, A.; Sarcina, L.; Athreya, S.; Bailey, S.E.; Rodrigo, R.; Mircea, G.; Higham, T.; Ramsey, C.B.; van der Plicht, J. (September 2003). "An early modern human from the Peştera cu Oase, Romania". Proceedings of the National Academy of Sciences of the United States of America. 100 (20): 11231–36. Bibcode:2003PNAS..10011231T. doi:10.1073/pnas.2035108100. ISSN 0027-8424. PMC 208740. PMID 14504393.
When multiple measurements are undertaken, the mean result can be determined through averaging the activity ratios. For Oase 1, this provides a weighted average activity ratio of 〈14a〉 = 1.29 ± 0.15%, resulting in a combined OxA-GrA 14C age of 34,950, +990, and −890 B.P.
- Tattersall, Ian & Schwartz, Jeffrey H. (1999). "Hominids and hybrids: The place of Neanderthals in human evolution". Proceedings of the National Academy of Sciences of the United States of America. 96 (13): 7117–19. Bibcode:1999PNAS...96.7117T. doi:10.1073/pnas.96.13.7117. PMC 33580. PMID 10377375.
- Cann, Rebecca L.; Stoneking, Mark; Wilson, Allan C. (1987-01-01). "Mitochondrial DNA and human evolution". Nature. 325 (6099): 31–36. Bibcode:1987Natur.325...31C. doi:10.1038/325031a0. PMID 3025745. S2CID 4285418. Archived from the original on 2010-08-13.
- McBride B, Haviland WE, Prins HE, Walrath D (2009). The Essence of Anthropology. Belmont, CA: Wadsworth Publishing. p. 90. ISBN 978-0-495-59981-4.
- Reid GR, Hetherington R (2010). The climate connection: climate change and modern human evolution. Cambridge, UK: Cambridge University Press. p. 64. ISBN 978-0-521-14723-1.
- Meredith M (2011). Born in Africa: The Quest for the Origins of Human Life. New York: PublicAffairs. ISBN 978-1-58648-663-1.
- Wolpoff, M., Caspari, R. (1997). Race and human evolution: A fatal attraction. New York: Simon and Schuster. p. 213.
- Wolpoff, M.; Thorne, A. (1991) "The case against Eve" New Scientist 130(1774): 37–41
- Curnoe, D.; Thorne, A. (2003). "Number of ancestral human species: a molecular perspective". HOMO: Journal of Comparative Human Biology. 53 (3): 201–24. doi:10.1078/0018-442x-00051. PMID 12733395.
- Wu, X. (2004). "Discussion on the results of some molecular studies concerning the origin of modern Chinese". Acta Anthropologica Sinica. 24 (4): 259–69.
- Thorne, A.G.; Wolpoff, M. H.; Eckhardt, R.B. (1993). "Genetic variation in Africa". Science. 261 (5128): 1507–1508. Bibcode:1993Sci...261.1507T. doi:10.1126/science.8372344. PMID 8372344.
- Wu, Xinzhi; Gao, Xing; Zhang, X.; Yang, D.; Shen, C. (2010). "Revisiting the origin of modern humans in China and its implications for global human evolution". Science China Earth Sciences. 53 (12): 1927–40. Bibcode:2010ScChD..53.1927G. doi:10.1007/s11430-010-4099-4. S2CID 195307737.
- Thorne, A.G.; Curnoe, D. (2006). "What is the real age of Adam and Eve? Proceedings of the Australian Society of Human Biology". HOMO: Journal of Comparative Human Biology. 57: 240.
- Relethford, J.H. (2008-03-05). "Genetic evidence and the modern human origins debate". Heredity. 100 (6): 555–63. doi:10.1038/hdy.2008.14. PMID 18322457.
- "Selection, nuclear genetic variation, and mtDNA". john hawks weblog. 5 September 2005. Retrieved 2011-01-05.
- Thorne, A.G.; Wolpoff, M.H. (2003). "The Multiregional Evolution of Humans". Scientific American. 13 (2): 46–53.
- Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S (July 1997). "Neandertal DNA sequences and the origin of modern humans". Cell. 90 (1): 19–30. doi:10.1016/S0092-8674(00)80310-4. hdl:11858/00-001M-0000-0025-0960-8. PMID 9230299. S2CID 13581775.
- Krings, M.; Capelli, C.; Tschentscher, F.; Geisert, H.; Meyer, S.; von Haeseler, A.; et al. (2000). "A view of Neandertal genetic diversity". Nature Genetics. 26 (2): 144–46. doi:10.1038/79855. PMID 11017066. S2CID 10426584.
- Wang, C.C.; Farina, S.E.; Li, H. (2013). "Neanderthal DNA and modern human origins". Quaternary International. 295: 126–29. Bibcode:2013QuInt.295..126W. doi:10.1016/j.quaint.2012.02.027.
- Relethford. John. H. (2001-04-12). "Absence of Regional Affinities of Neandertal DNA with Living Humans Does Not Reject Multiregional Evolution". American Journal of Physical Anthropology. 115 (1): 95–98. doi:10.1002/ajpa.1060. PMID 11309754.
- Wolpoff, M. (1998). "Concocting a Divisive Theory". Evolutionary Anthropology. 7 (1): 1–3. doi:10.1002/(sici)1520-6505(1998)7:1<1::aid-evan1>3.3.co;2-w.
- Hammer, M.F.; et al. (1998). "Out of Africa and Back Again: Nested Cladistic Analysis of Human Y Chromosome Variation". Molecular Biology and Evolution. 15 (4): 427–41. doi:10.1093/oxfordjournals.molbev.a025939. PMID 9549093.
- Hammer, M.F.; Garrigan, D.; Wood, E.; Wilder, J.A.; Mobasher, Z.; Bigham, A.; Krenz, J.G.; Nachman, M.W. (August 2004). "Heterogeneous patterns of variation among multiple human x-linked Loci: the possible role of diversity-reducing selection in non-africans" (Free full text). Genetics. 167 (4): 1841–53. doi:10.1534/genetics.103.025361. ISSN 0016-6731. PMC 1470985. PMID 15342522. Additional discussion of these results is available in a video of a presentation given by Hammer at https://www.youtube.com/watch?v=Ff0jwWaPlnU (video) from about 40 to 50 minutes into the video.
- The CMP-N-acetylneuraminic acid hydroxylase CMAH pseudogene shows 2.9 Mya coalescence time. Hayakawa, T; Aki, I; Varki, A; Satta, Y; Takahata, N (February 2006). "Fixation of the Human-Specific CMP-N-Acetylneuraminic Acid Hydroxylase Pseudogene and Implications of Haplotype Diversity for Human Evolution". Genetics. 172 (2): 1139–46. doi:10.1534/genetics.105.046995. ISSN 0016-6731. PMC 1456212. PMID 16272417.
- The PDHA1 (pyruvate dehydrogenase) locus on the X chromosome has an estimated coalescence time of 1.86 Mya, inconsistent with a recent species origin, although the worldwide lineage pattern is unlike other autosomal sites and may be consistent with recent dispersal from Africa. Rosalind M. Harding (March 16, 1999). "More on the X files". Proceedings of the National Academy of Sciences. 96 (6): 2582–84. Bibcode:1999PNAS...96.2582H. doi:10.1073/pnas.96.6.2582. PMC 33533. PMID 10077551.
- A second group finds the same ancient origin for PDHA1, but finds no evidence of a recent expansion, consistent with other autosomal and X chromosome sites and contrary to mitochondrial data. Harris, E. E.; Jody Hey (1999). "X chromosome evidence for ancient human histories". Proceedings of the National Academy of Sciences. 96 (6): 3320–24. Bibcode:1999PNAS...96.3320H. doi:10.1073/pnas.96.6.3320. PMC 15940. PMID 10077682.
- The ASAH1 gene has two recently differentiated lineages with a coalescence time 2.4±.4 Mya not explainable by balancing selection. The V lineage shows evidence of recent positive selection. The lineage pattern may be the result of hybridization during a recent range expansion from Africa with the V lineage tracing to archaic humans from outside Africa, though it is also consistent with a mixture of two long isolated groups within Africa; it is not consistent with a recent origination of a modern human species that replaced archaic forms without interbreeding. Kim, Hl; Satta, Y (March 2008). "Population Genetic Analysis of the N-Acylsphingosine Amidohydrolase Gene Associated With Mental Activity in Humans". Genetics. 178 (3): 1505–15. doi:10.1534/genetics.107.083691. ISSN 0016-6731. PMC 2278054. PMID 18245333.
It is speculated that, when modern humans dispersed from Africa, admixture of the distinct V and M lineages occurred and the V lineage has since spread in the entire population by possible positive selection.
- Daniel Garrigan; Zahra Mobasher; Sarah B. Kingan; Jason A. Wilder; Michael F. Hammer (August 2005). "Deep Haplotype Divergence and Long-Range Linkage Disequilibrium at Xp21.1 Provide Evidence That Humans Descend From a Structured Ancestral Population". Genetics. 170 (4): 1849–56. doi:10.1534/genetics.105.041095. PMC 1449746. PMID 15937130.
- NAT2 SNPs lineages cluster in sub-Saharan Africa, Europe, and East Asia, with genetic distances scaling with geographic distances. Sabbagh, A.; Langaney, A.; Darlu, P.; Gérard, N.; Krishnamoorthy, R.; Poloni, E. S. (February 2008). "Worldwide distribution of NAT2 diversity: Implications for NAT2 evolutionary history". BMC Genetics. 9: 21. doi:10.1186/1471-2156-9-21. PMC 2292740. PMID 18304320. Also see map; may resize browser window.
- The NAT1 lineage tree is rooted in Eurasia with a coalescence time of 2.0 Mya that cannot be explained by balancing selection and with the NAT1*11A haplotype absent from subsaharan Africa. Patin, E.; Barreiro, L.B.; Sabeti, P.C.; et al. (March 2006). "Deciphering the Ancient and Complex Evolutionary History of Human Arylamine N-Acetyltransferase Genes". American Journal of Human Genetics. 78 (3): 423–36. doi:10.1086/500614. ISSN 0002-9297. PMC 1380286. PMID 16416399.
- "Variation in NAT1 and NAT2". john hawks weblog. 15 January 2006. Retrieved 2011-01-04.
- Garrigan, D.; Mobasher, Z.; Severson, T.; Wilder, J. A.; Hammer, M.F. (February 2005). "Evidence for archaic Asian ancestry on the human X chromosome" (Free full text). Molecular Biology and Evolution. 22 (2): 189–92. doi:10.1093/molbev/msi013. ISSN 0737-4038. PMID 15483323.
- Cox, M.P.; Mendez, F.L.; Karafet, T.M.; Pilkington, M.M.; Kingan, S.B.; Destro-Bisol, G.; Strassmann, B.I.; Hammer, M.F. (January 2008). "Testing for Archaic Hominin Admixture on the X Chromosome: Model Likelihoods for the Modern Human RRM2P4 Region From Summaries of Genealogical Topology Under the Structured Coalescent" (Free full text). Genetics. 178 (1): 427–37. doi:10.1534/genetics.107.080432. ISSN 0016-6731. PMC 2206091. PMID 18202385.
- J. Hardy; A. Pittman; A. Myers; K. Gwinn-Hardy; H. C. Fung; R. de Silva; M. Hutton; J. Duckworth (2005). "Evidence suggesting that Homo neanderthalensis contributed the H2 MAPT haplotype to Homo sapiens". Biochemical Society Transactions. 33 (4): 582–85. doi:10.1042/bst0330582. PMID 16042549.
- Zody, M. C.; Jiang, Z.; Fung, H. C.; et al. (August 2008). "Evolutionary Toggling of the MAPT 17q21.31 Inversion Region". Nature Genetics. 40 (9): 1076–83. doi:10.1038/ng.193. ISSN 1061-4036. PMC 2684794. PMID 19165922.
- Almos, P. Z.; Horváth, S.; Czibula, A.; Raskó, I.; Sipos, B.; Bihari, P.; Béres, J.; Juhász, A.; Janka, Z.; Kálmán, J. (November 2008). "H1 tau haplotype-related genomic variation at 17q21.3 as an Asian heritage of the European Gypsy population". Heredity. 101 (5): 416–19. doi:10.1038/hdy.2008.70. ISSN 0018-067X. PMID 18648385.
- Stefansson H.; Helgason A.; Thorleifsson G.; et al. (2005-01-16). "A common inversion under selection in Europeans". Nature Genetics. 37 (2): 129–37. doi:10.1038/ng1508. PMID 15654335. S2CID 120515.
- Evans, P. D.; Mekel-Bobrov, N.; Vallender, E. J.; Hudson, R. R.; Lahn, B. T. (November 2006). "Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage". Proceedings of the National Academy of Sciences of the United States of America. 103 (48): 18178–83. Bibcode:2006PNAS..10318178E. doi:10.1073/pnas.0606966103. ISSN 0027-8424. PMC 1635020. PMID 17090677.
- Trinkaus, E. (May 2007). "European early modern humans and the fate of the Neandertals". Proceedings of the National Academy of Sciences of the United States of America. 104 (18): 7367–72. Bibcode:2007PNAS..104.7367T. doi:10.1073/pnas.0702214104. ISSN 0027-8424. PMC 1863481. PMID 17452632.
- Evans, P. D.; Gilbert, S. L.; Mekel-Bobrov, N.; Vallender, E. J.; Anderson, J. R.; Vaez-Azizi, L. M.; Tishkoff, S. A.; Hudson, R. R.; Lahn, B. T. (September 2005). "Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans". Science. 309 (5741): 1717–20. Bibcode:2005Sci...309.1717E. doi:10.1126/science.1113722. ISSN 0036-8075. PMID 16151009. S2CID 85864492.
- "Introgression and microcephalin FAQ". john hawks weblog. 8 November 2006. Retrieved 2011-01-05.
- Pennisi E (February 2009). "Neandertal genomics. Tales of a prehistoric human genome". Science. 323 (5916): 866–71. doi:10.1126/science.323.5916.866. PMID 19213888. S2CID 206584252.
- Green RE, Krause J, Briggs AW, Maricic T, Stenzel U, Kircher M, et al. (May 2010). "A draft sequence of the Neandertal genome". Science. 328 (5979): 710–722. Bibcode:2010Sci...328..710G. doi:10.1126/science.1188021. PMC 5100745. PMID 20448178.
- Lari M, Rizzi E, Milani L, Corti G, Balsamo C, Vai S, Catalano G, Pilli E, Longo L, Condemi S, Giunti P, Hänni C, De Bellis G, Orlando L, Barbujani G, Caramelli D (May 2010). "The microcephalin ancestral allele in a Neanderthal individual". PLOS ONE. 5 (5): e10648. Bibcode:2010PLoSO...510648L. doi:10.1371/journal.pone.0010648. PMC 2871044. PMID 20498832.
- Ke, Yuehai; et al. (2001). "African Origin of Modern Humans in East Asia: A Tale of 12,000 Y Chromosomes". Science. 292 (5519): 1151–53. Bibcode:2001Sci...292.1151K. doi:10.1126/science.1060011. PMID 11349147. S2CID 32685801.
- Templeton, Alan R. (2005). "Haplotype Trees and Modern Human Origins" (PDF). Yearbook of Physical Anthropology. 48 (S41): 33–59. doi:10.1002/ajpa.20351. PMID 16369961.
- Yotova, Vania; Lefebvre, Jean-Francois; et al. (Jul 2011). "An X-Linked Haplotype of Neandertal Origin Is Present Among All Non-African Populations". Molecular Biology and Evolution. 28 (7): 1957–62. doi:10.1093/molbev/msr024. PMID 21266489.
- Reich; et al. (2011). "Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania" (PDF). The American Journal of Human Genetics. 89 (4): 516–528. doi:10.1016/j.ajhg.2011.09.005. PMC 3188841. PMID 21944045. Archived from the original (PDF) on 2014-10-09. Retrieved 2013-09-07.
- Noonan, James P.; et al. (2006-11-17). "Sequencing and Analysis of Neanderthal Genomic DNA". Science. 314 (5802): 1113–18. Bibcode:2006Sci...314.1113N. doi:10.1126/science.1131412. PMC 2583069. PMID 17110569.
- Green, Richard E.; et al. (2010-05-07). "A Draft Sequence of the Neandertal Genome". Science. 328 (5979): 710–22. Bibcode:2010Sci...328..710G. doi:10.1126/science.1188021. PMC 5100745. PMID 20448178.
- "Neandertals Live!". john hawks weblog. 6 May 2010. Retrieved 2010-12-31.
- Reich, David; et al. (2010-12-23). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature. 468 (7327): 1053–60. Bibcode:2010Natur.468.1053R. doi:10.1038/nature09710. PMC 4306417. PMID 21179161.
- "The Denisova genome FAQ". john hawks weblog. 22 December 2010. Retrieved 2010-12-31.
- Laurent Abi-Rached; et al. (2011-08-25). "The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans". Science. 334 (6052): 89–94. Bibcode:2011Sci...334...89A. doi:10.1126/science.1209202. PMC 3677943. PMID 21868630.
-
- Witherspoon, D. J.; Wooding, S.; Rogers, A. R.; Marchani, E. E.; Watkins, W. S.; Batzer, M. A.; Jorde, L. B. (2007). "Genetic Similarities Within and Between Human Populations". Genetics. 176 (1): 351–59. doi:10.1534/genetics.106.067355. PMC 1893020. PMID 17339205.
- Witherspoon DJ, Wooding S, Rogers AR, et al. (May 2007). "Genetic Similarities Within and Between Human Populations". Genetics. 176 (1): 351–59. doi:10.1534/genetics.106.067355. PMC 1893020. PMID 17339205.
Further reading
- Biogeography of Middle Pleistocene hominins in mainland Southeast Asia: A review of current evidence, Academia.edu, 23 July 2008. retrieved on 4, April 2014
External links
- From www.conrante.com: Templeton's lattice diagram showing major gene flows graphically.
- From darwin.eeb.uconn.edu: notes on drift and migration with equations for calculating the effects on allele frequencies of different populations.
- Human Evolution. (2011). In Encyclopædia Britannica. Retrieved from http://www.britannica.com/EBchecked/topic/275670/human-evolution
- Plural Lineages in the Human mtDNA Genome
- Human Timeline (Interactive) – Smithsonian, National Museum of Natural History (August 2016).
{kind=link}
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines |
| ||||||||||||||||||||||||||
| Others |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Sub-topics | |
|---|---|
| Genetic history by region | |
| Population genetics by group |
|
| |