Reverse Krebs cycle
The reverse Krebs cycle (also known as the reverse tricarboxylic acid cycle, the reverse TCA cycle, or the reverse citric acid cycle, or the reductive tricarboxylic acid cycle, or the reductive TCA cycle) is a sequence of chemical reactions that are used by some bacteria to produce carbon compounds from carbon dioxide and water by the use of energy-rich reducing agents as electron donors.
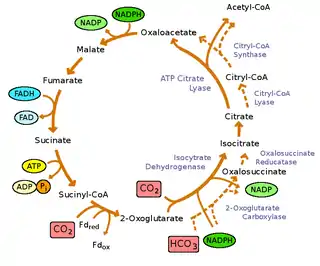
The reaction is the citric acid cycle run in reverse. Where the Krebs cycle takes carbohydrates and oxidizes them to CO2 and water, the reverse cycle takes CO2 and H2O to make carbon compounds. This process is used by some bacteria (such as Aquificota) to synthesize carbon compounds, sometimes using hydrogen, sulfide, or thiosulfate as electron donors.[1][2] This process can be seen as an alternative to the fixation of inorganic carbon in the reductive pentose phosphate cycle which occurs in a wide variety of microbes and higher organisms.
Differences from Krebs cycle
In contrast to the oxidative citric acid cycle, the reverse or reductive cycle has a few key differences. There are three enzymes specific to the reductive citric acid cycle – citrate lyase, fumarate reductase, and α-ketoglutarate synthase.
The splitting of citric acid to oxaloacetate and acetate is in catalyzed by citrate lyase, rather than the reverse reaction of citrate synthase.[3] Succinate dehydrogenase is replaced by fumarate reductase and α-ketoglutarate synthase replaces α-ketoglutarate dehydrogenase.
The conversion of succinate to 2-oxoglutarate is also different. In the oxidative reaction this step is coupled to the reduction of NADH. However, the oxidation of 2-oxoglutarate to succinate is so energetically favorable, that NADH lacks the reductive power to drive the reverse reaction. In the rTCA cycle, this reaction has to use a reduced low potential ferredoxin.[4]
Relevance to early life
The reaction is a possible candidate for prebiotic early-earth conditions and, therefore, is of interest in the research of the origin of life. It has been found that some non-consecutive steps of the cycle can be catalyzed by minerals through photochemistry,[5] while entire two and three-step sequences can be promoted by metal ions such as iron (as reducing agents) under acidic conditions. In addition, these organisms that undergo photochemistry can and do utilize the citric acid cycle.[1] However, the conditions are extremely harsh and require 1 M hydrochloric or 1 M sulfuric acid and strong heating at 80–140 °C.[6]
Along with the these possibilities of the rTCA cycle contributing to early life and biomolecules, it is thought that the rTCA cycle could not have been completed without the use of enzymes. The kinetic and thermodynamic parameters of the reduction of highly oxidized species to push the rTCA cycle are seemingly unlikely without the necessary action of biological catalysts known as enzymes. The rate of some of the reactions in the rTCA cycle likely would have been too slow to contribute significantly to the formation of life on earth without enzymes. Considering the thermodynamics of the rTCA cycle, the increase in Gibbs free energy going from product to reactant would make pyrophosphate an unlikely energy source for the conversion of pyruvate to oxaloacetate as the reaction is too endoergic.[7] However, it is suggested that a nonenzymatic precursor to the Krebs cycle, glyoxylate cycle, and reverse Krebs cycle might have originated, where oxidation and reduction reactions cooperated. The later use of carboxylation utilizing ATP could have given rise to parts of reverse Krebs cycle.[8]
It is suggested that the reverse Krebs cycle was incomplete, even in the last universal common ancestor.[9][10] Many reactions of the reverse Krebs cycle, including thioesterification and hydrolysis, could have been catalyzed by iron-sulfide minerals at deep sea alkaline hydrothermal vent cavities.[11] More recently, aqueous microdroplets have been shown to promote reductive carboxylation reactions in the reverse Krebs cycle.[12]
Medical relevance
The reverse Krebs cycle is proposed to be a major role in the pathophysiology of melanoma. Melanoma tumors are known to alter normal metabolic pathways in order to utilize waste products. These metabolic adaptations help the tumor adapt to its metabolic needs. The most well known adaptation is the Warburg effect where tumors increase their uptake and utilization of glucose. Glutamine is one of the known substances to be utilized in the reverse Krebs cycle in order to produce acetyl-CoA.[13] This type of mitochondrial activity could provide a new way to identify and target cancer causing cells.[14]
Microbial use of the reverse Krebs cycle
Thiomicrospira denitrificans, Candidatus Arcobacter, and Chlorobaculum tepidum have been shown to utilize the rTCA cycle to turn CO2 into carbon compounds. The ability of these bacteria, among others, to use the rTCA cycle, supports the idea that they are derived from an ancestral proteobacterium, and that other organisms using this cycle are much more abundant than previously believed.[15]
See also
References
- Evans MC, Buchanan BB, Arnon DI (April 1966). "A new ferredoxin-dependent carbon reduction cycle in a photosynthetic bacterium". Proceedings of the National Academy of Sciences of the United States of America. 55 (4): 928–934. Bibcode:1966PNAS...55..928E. doi:10.1073/pnas.55.4.928. PMC 224252. PMID 5219700.
- Buchanan BB, Arnon DI (1990). "A reverse KREBS cycle in photosynthesis: consensus at last". Photosynthesis Research. 24: 47–53. doi:10.1007/BF00032643. PMID 11540925. S2CID 2753977.
- Bar-Even A, Noor E, Milo R (March 2012). "A survey of carbon fixation pathways through a quantitative lens". Journal of Experimental Botany. 63 (6): 2325–2342. doi:10.1093/jxb/err417. PMID 22200662.
- Bar-Even A, Noor E, Milo R (March 2012). "A survey of carbon fixation pathways through a quantitative lens". Journal of Experimental Botany. 63 (6): 2325–2342. doi:10.1093/jxb/err417. PMID 22200662.
- Zhang XV, Martin ST (December 2006). "Driving parts of Krebs cycle in reverse through mineral photochemistry". Journal of the American Chemical Society. 128 (50): 16032–16033. doi:10.1021/ja066103k. PMID 17165745.
- Muchowska KB, Varma SJ, Chevallot-Beroux E, Lethuillier-Karl L, Li G, Moran J (November 2017). "Metals promote sequences of the reverse Krebs cycle". Nature Ecology & Evolution. 1 (11): 1716–1721. doi:10.1038/s41559-017-0311-7. PMC 5659384. PMID 28970480.
- Ross DS (February 2007). "The viability of a nonenzymatic reductive citric acid cycle--kinetics and thermochemistry". Origins of Life and Evolution of the Biosphere. 37 (1): 61–65. Bibcode:2007OLEB...37...61R. doi:10.1007/s11084-006-9017-6. PMID 17136437. S2CID 2208326.
- Muchowska, Kamila B.; Varma, Sreejith J.; Moran, Joseph (1 May 2019). "Synthesis and breakdown of universal metabolic precursors promoted by iron". Nature. 569 (7754): 104–107. Bibcode:2019Natur.569..104M. doi:10.1038/s41586-019-1151-1. ISSN 1476-4687. PMC 6517266. PMID 31043728.
- Harrison SA, Palmeira RN, Halpern A, Lane N (November 2022). "A biophysical basis for the emergence of the genetic code in protocells". Biochimica et Biophysica Acta. Bioenergetics. 1863 (8): 148597. doi:10.1016/j.bbabio.2022.148597. PMID 35868450.
- Muchowska KB, Varma SJ, Moran J (August 2020). "Nonenzymatic Metabolic Reactions and Life's Origins" (PDF). Chemical Reviews. 120 (15): 7708–7744. doi:10.1021/acs.chemrev.0c00191. PMID 32687326. S2CID 220671580.
- Akbari A, Palsson BO (May 2023). "Metabolic homeostasis and growth in abiotic cells". Proceedings of the National Academy of Sciences of the United States of America. 120 (19): e2300687120. Bibcode:2023PNAS..12000687A. doi:10.1073/pnas.2300687120. PMC 10175716. PMID 37126695.
- Ju, Yun; Zhang, Hong; Jiang, Yanxiao; Wang, Wenxin; Kan, Guangfeng; Yu, Kai; Wang, Xiaofei; Liu, Jilin; Jiang, Jie (2023-09-07). "Aqueous microdroplets promote C–C bond formation and sequences in the reverse tricarboxylic acid cycle". Nature Ecology & Evolution: 1–11. doi:10.1038/s41559-023-02193-8. ISSN 2397-334X. PMID 37679455. S2CID 261609019.
- Filipp FV, Scott DA, Ronai ZA, Osterman AL, Smith JW (May 2012). "Reverse TCA cycle flux through isocitrate dehydrogenases 1 and 2 is required for lipogenesis in hypoxic melanoma cells". Pigment Cell & Melanoma Research. 25 (3): 375–383. doi:10.1111/j.1755-148X.2012.00989.x. PMC 3329592. PMID 22360810.
- Wise DR, Ward PS, Shay JE, Cross JR, Gruber JJ, Sachdeva UM, et al. (December 2011). "Hypoxia promotes isocitrate dehydrogenase-dependent carboxylation of α-ketoglutarate to citrate to support cell growth and viability". Proceedings of the National Academy of Sciences of the United States of America. 108 (49): 19611–19616. Bibcode:2011PNAS..10819611W. doi:10.1073/pnas.1117773108. PMC 3241793. PMID 22106302.
- Hügler M, Wirsen CO, Fuchs G, Taylor CD, Sievert SM (May 2005). "Evidence for autotrophic CO2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria". Journal of Bacteriology. 187 (9): 3020–3027. doi:10.1128/JB.187.9.3020-3027.2005. PMC 1082812. PMID 15838028.