Seagrass meadow
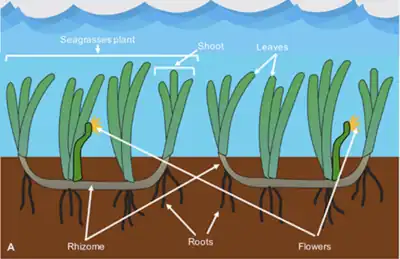
A seagrass meadow or seagrass bed is an underwater ecosystem formed by seagrasses. Seagrasses are marine (saltwater) plants found in shallow coastal waters and in the brackish waters of estuaries. Seagrasses are flowering plants with stems and long green, grass-like leaves. They produce seeds and pollen and have roots and rhizomes which anchor them in seafloor sand.
| Marine habitats |
|---|
 |
Open ocean |
Seagrasses form dense underwater meadows which are among the most productive ecosystems in the world. They provide habitats and food for a diversity of marine life comparable to that of coral reefs. This includes invertebrates like shrimp and crabs, cod and flatfish, marine mammals and birds. They provide refuges for endangered species such as seahorses, turtles, and dugongs. They function as nursery habitats for shrimps, scallops and many commercial fish species. Seagrass meadows provide coastal storm protection by the way their leaves absorb energy from waves as they hit the coast. They keep coastal waters healthy by absorbing bacteria and nutrients, and slow the speed of climate change by sequestering carbon dioxide into the sediment of the ocean floor.
Seagrasses evolved from marine algae which colonized land and became land plants, and then returned to the ocean about 100 million years ago. However, today seagrass meadows are being damaged by human activities such as pollution from land runoff, fishing boats that drag dredges or trawls across the meadows uprooting the grass, and overfishing which unbalances the ecosystem. Seagrass meadows are currently being destroyed at a rate of about two football fields every hour.
Background

Seagrasses are flowering plants (angiosperms) which grow in marine environments. They evolved from terrestrial plants which migrated back into the ocean about 75 to 100 million years ago.[1][2] These days they occupy the sea bottom in shallow and sheltered coastal waters anchored in sand or mud bottoms.[3]
There are four lineages of seagrasses [4] containing relatively few species (all in a single order of monocotyledon). They occupy shallow environments on all continents except Antarctica:[5] their distribution also extends to the High Seas, such as on the Mascarene Plateau.
Seagrasses are formed by a polyphyletic group of monocotyledons (order Alismatales), which recolonised marine environments about 80 million years ago.[4] Seagrasses are habitat-forming species because they are a source of food and shelter for a wide variety of fish and invertebrates, and they perform relevant ecosystem services.[6][7]
There are about 60 species of fully marine seagrasses belonging to four families (Posidoniaceae, Zosteraceae, Hydrocharitaceae and Cymodoceaceae), all in the order Alismatales (in the class of monocotyledons).[8] Seagrasses beds or meadows can be either made up of a single species (monospecific) or mixed. In temperate areas one or a few species usually dominate (like the eelgrass Zostera marina in the North Atlantic), whereas tropical beds are usually more diverse, with up to thirteen species recorded in the Philippines. Like all autotrophic plants, seagrasses photosynthesize, in the submerged photic zone. Most species undergo submarine pollination and complete their life cycle underwater.[9]
Seagrass meadows are found in depths up to about 50m, depending on water quality and light availability.[10] These seagrass meadows are highly productive habitats that provide many ecosystem services, including protecting the coast from storms and big waves, stabilising sediment, providing safe habitats for other species and encouraging biodiversity, enhancing water quality, and sequestering carbon and nutrients.[11][3]
Seagrass meadows are sometimes called prairies of the sea. They are diverse and productive ecosystems sheltering to and harbouring species from all phyla, such as juvenile and adult fish, epiphytic and free-living macroalgae and microalgae, mollusks, bristle worms, and nematodes. Few species were originally considered to feed directly on seagrass leaves (partly because of their low nutritional content), but scientific reviews and improved working methods have shown that seagrass herbivory is an important link in the food chain, feeding hundreds of species, including green turtles, dugongs, manatees, fish, geese, swans, sea urchins and crabs. Some fish species that visit or feed on seagrasses raise their young in adjacent mangroves or coral reefs.[9]

Seagrass meadows are rich biodiverse ecosystems that occur all over the globe, in both tropical and temperate seas.[14] They contain complex food webs that provide trophic subsidy to species and habitats way beyond the extent of their distribution.[15] Given the wide variety of food sources provided by this productive habitat, it is no surprise that seagrass meadows support an equally wide array of grazers and predators. However, despite its importance for sustaining biodiversity and many other ecosystem services,[16] the global distribution of seagrass is a fraction of what was historically present.[17][18] Recent estimates from where records exist indicate that at least 20% of the world's seagrass has been lost.[18] Seagrasses also provide other services in the coastal zone such as preventing coastal erosion, storing and trapping carbon [19] and filtering the water column.[20][21]
The true ecosystem-level consequences of such decline and the benefits that can be afforded through habitat restoration are poorly understood. Given the relatively high-per-unit area costs of marine habitat restoration,[22] making the case for such work requires a thorough examination of the ecosystem service benefits of such new habitat creation.[21]
Global distribution
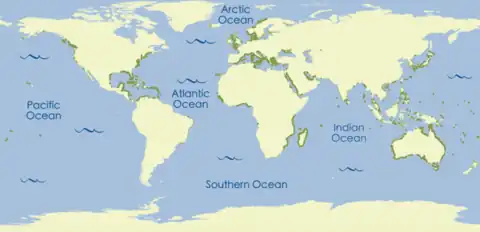
Seagrass meadows are found in the shallow seas of the continental shelves of all continents except Antarctica. Continental shelves are underwater areas of land surrounding each continent, creating areas of relatively shallow water known as shelf seas.[12] The grasses live in areas with soft sediment that are either intertidal (uncovered daily by seawater, as the tide goes in and out) or subtidal (always under the water). They prefer sheltered places, such as shallow bays, lagoons, and estuaries (sheltered areas where rivers flow in to the sea), where waves are limited and light and nutrient levels are high.
Seagrasses can survive to maximum depths of about 60 metres. However, this depends on the availability of light, because, like plants on the land, seagrass meadows need sunlight if photosynthesis is to occur. Tides, wave action, water clarity, and low salinity (low amounts of salt in the water) control where seagrasses can live at their shallow edge nearest the shore;[23] all of these things must be right for seagrass to survive and grow.[12]
The current documented seagrass area is 177,000 km2 (68,000 sq mi), but is thought to underestimate the total area since many areas with large seagrass meadows have not been thoroughly documented.[10] Most common estimates are 300,000 to 600,000 km2, with up to 4,320,000 km2 suitable seagrass habitat worldwide.[24]
Ecosystem services
Seagrass meadows provide coastal zones with significant ecosystem goods and services. They enhance water quality by stabilizing heavy metals and other toxic pollutants, as well as cleansing the water of excess nutrients,[25][2][1] and lowering acidity levels in coastal waters.[26][27] Further, because seagrasses are underwater plants, they produce significant amounts of oxygen which oxygenate the water column. Their root systems also assist in oxygenating the sediment, providing hospitable environments for sediment-dwelling organisms.[28] Additionally, the conservation of seagrass meadows contributes to 16 of the 17 UN Sustainable Development Goals.[29]
_(South_Pigeon_Creek_estuary%252C_San_Salvador_Island%252C_Bahamas)_2_(16043820341).jpg.webp)
.jpg.webp)
As shown in the image above on the left, many epiphytes can grow on the leaf blades of seagrasses, and algae, diatoms and bacterial films can cover the surface. The grass is eaten by turtles, herbivorous parrotfish, surgeonfish, and sea urchins, while the leaf surface films are a food source for many small invertebrates.[30]
Blue carbon
The meadows also account for more than 10% of the ocean's total carbon storage. Per hectare, they hold twice as much carbon dioxide as rain forests and can sequester about 27 million tons of CO2 annually.[31] This ability to store carbon is important as atmospheric carbon levels continue to rise.
Blue carbon refers to carbon dioxide removed from the atmosphere by the world's coastal marine ecosystems, mostly mangroves, salt marshes, seagrasses and potentially macroalgae, through plant growth and the accumulation and burial of organic matter in the sediment.[32][33]
Although seagrass meadows occupy only 0.1% of the area of the ocean floor, they account for 10–18% of the total oceanic carbon burial.[34] Currently global seagrass meadows are estimated to store as much as 19.9 Pg (petagrams or gigatons, equals a billion tons) of organic carbon.[34] Carbon primarily accumulates in marine sediments, which are anoxic and thus continually preserve organic carbon from decadal-millennial time scales. High accumulation rates, low oxygen, low sediment conductivity and slower microbial decomposition rates all encourage carbon burial and carbon accumulation in these coastal sediments.[10] Compared to terrestrial habitats that lose carbon stocks as CO2 during decomposition or by disturbances like fires or deforestation, marine carbon sinks can retain C for much longer time periods. Carbon sequestration rates in seagrass meadows vary depending on the species, characteristics of the sediment, and depth of the habitats, but on average the carbon burial rate is about 140 g C m−2 yr−1.[25][35]
Coastal protection
Seagrasses are also ecosystem engineers, which means they alter the ecosystem around them, adjusting their surroundings in both physical and chemical ways.[2][1] The long blades of seagrasses slow the movement of water which reduces wave energy and offers further protection against coastal erosion and storm surge. Many seagrass species produce an extensive underground network of roots and rhizome which stabilizes sediment and reduces coastal erosion.[36] Seagrasses are not only affected by water in motion; they also affect the currents, waves and turbulence environment.[37]
Seagrasses prevent erosion of the seafloor to the point that their presence can raise the seafloor. They contribute to coast protection by trapping rock debris transported by the sea. Seagrasses reduce erosion of the coast and protect houses and cities from both the force of the sea and from sea-level rise caused by global warming. They do this by softening the force of the waves with their leaves, and helping sediment transported in the seawater to accumulate on the seafloor. Seagrass leaves act as baffles in turbulent water that slow down water movement and encourage particulate matter to settle out. Seagrass meadows are one of the most effective barriers against erosion, because they trap sediment amongst their leaves.[3]

The diagram on the left above illustrates how seagrasses help trap sediment particles transported by sea currents. The leaves, extending toward the sea surface, slow down the water currents. The slower current is not able to carry the particles of sediment, so the particles drop down and become part of the seafloor, eventually building it up. When seagrasses are not present, the sea current has no obstacles and carries the sediment particles away, lifting them and eroding the seafloor.[3]
Archaeologists have learned from seagrasses how to protect underwater archaeological sites, like a site in Denmark where dozens of ancient Roman and Viking shipwrecks have been discovered. The archaeologists use seagrass-like covers as sediment traps, to build up sediment so that it buries the ships. Burial creates low-oxygen conditions and keeps the wood from rotting.[39][3]
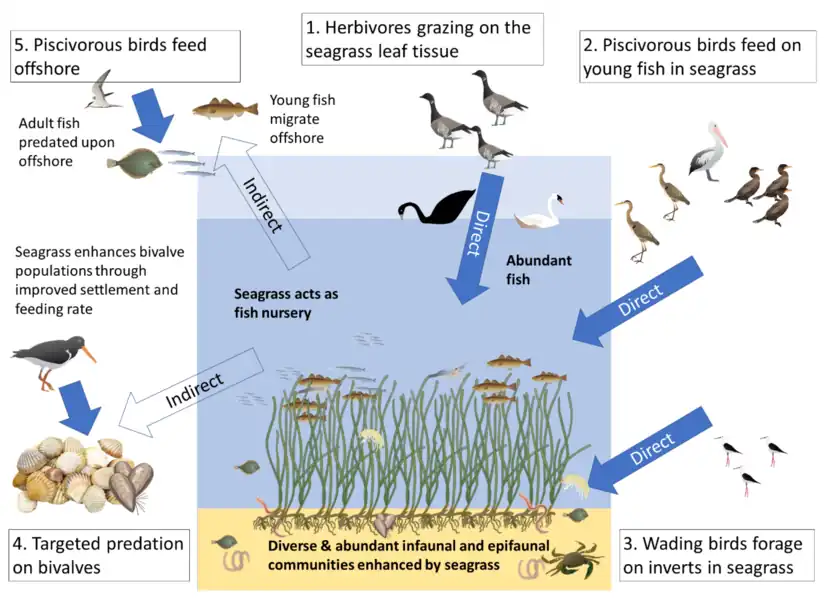 Hypothesised links in NE Atlantic coastal seas
Hypothesised links in NE Atlantic coastal seas
(1) direct consumption of seagrass (2,3) seagrass-associated fauna
(4,5) indirect links from consumption of populations benefitting from seagrass [21]
Links to seabirds
Birds are an often-overlooked part of marine ecosystems, not only are they crucial to the health of marine ecosystems, but their populations are also supported by the productivity and biodiversity of marine and coastal ecosystems.[40][41][42] The links of birds to specific habitat types such as seagrass meadows are largely not considered except in the context of direct herbivorous consumption by wildfowl.[43] This is despite the fact that both bottom-up and top-down processes have been considered as pathways for the population maintenance of some coastal birds.[44][21]
Given the long-term decline in the population of many coastal and seabirds, the known response of many seabird populations to fluctuations in their prey, and the need for compensatory restorative actions to enhance their populations, there is a need for understanding the role of key marine habitats such as seagrass in supporting coastal and seabirds.[21]

Nursery habitats for fisheries
Seagrass meadows provide nursery habitats for many commercially important fish species. It's estimated that about half of the global fisheries get their start because they are supported by seagrass habitats. If the seagrass habitats are lost, then the fisheries are lost as well. According to a 2019 paper by Unsworth et al,[45] the significant role seagrass meadows play in supporting fisheries productivity and food security across the globe is not adequately reflected in the decisions made by authorities with statutory responsibility for their management. They argue that: (1) Seagrass meadows provide valuable nursery habitat to over 1/5th of the world's largest 25 fisheries, including walleye pollock, the most landed species on the planet. (2) In complex small‐scale fisheries from around the world (poorly represented in fisheries statistics), there is evidence that many of those in proximity to seagrass are supported to a large degree by these habitats. (3) Intertidal fishing activity in seagrass is a global phenomenon, often directly supporting human livelihoods. According to the study, seagrasses should be recognized and managed to maintain and maximize their role in global fisheries production.[45] In 2022, Jones et al [46] showed seagrass associated small-scale fisheries can provide a safety net for the poor, and are used more commonly than reef-associated fisheries across the Indo-Pacific. Nearly half the people interviewed in the study preferred fishing in seagrass, since their function as a nursery habitat could result in large and reliable catches of fish.[46]
 Seagrass meadows support global food security by (1) providing nursery habitat for fish stocks in adjacent and deep water habitats, (2) creating expansive fishery habitat rich in fauna, and (3) by providing trophic support to adjacent fisheries. They also provide support by promoting the health of fisheries associated with connected habitats, such as coral reefs.[45]
Seagrass meadows support global food security by (1) providing nursery habitat for fish stocks in adjacent and deep water habitats, (2) creating expansive fishery habitat rich in fauna, and (3) by providing trophic support to adjacent fisheries. They also provide support by promoting the health of fisheries associated with connected habitats, such as coral reefs.[45]

In the oceans, gleaning can be defined as fishing with basic gear, including bare hands, in shallow water not deeper than that one can stand.[48] Invertebrate gleaning (walking) fisheries are common within intertidal seagrass meadows globally, contributing to the food supply of hundreds of millions of people, but understanding of these fisheries and their ecological drivers are extremely limited. A 2019 study by Nessa et al. analysed these fisheries using a combined social and ecological approach. Catches were dominated by bivalves, sea urchins and gastropods. The catch per unit effort (CPUE) in all sites varied from 0.05 to 3 kg per gleaner per hour, with the majority of fishers being women and children. Landings were of major significance for local food supply and livelihoods at all sites. Local ecological knowledge suggests seagrass meadows are declining in line with other regional trends. Increasing seagrass density significantly and positively correlated with CPUE of the invertebrate gleaning highlighting the importance of conserving these threatened habitats.[47]
Habitats for other species



_(southeastern_Graham's_Harbour%252C_San_Salvador_Island%252C_Bahamas)_2_(15428049493).jpg.webp)

_(South_Pigeon_Creek_estuary%252C_San_Salvador_Island%252C_Bahamas)_3_(15859724719).jpg.webp)

 Stingray in seagrass
Stingray in seagrass Peppered moray in seagrass
Peppered moray in seagrass
.jpg.webp) Blackspot emperor in seagrass
Blackspot emperor in seagrass
Other services
Historically, seagrasses were collected as fertilizer for sandy soil. This was an important use in the Aveiro Lagoon, Portugal, where the plants collected were known as moliço. In the early 20th century, in France and, to a lesser extent, the Channel Islands, dried seagrasses were used as a mattress (paillasse) filling – such mattresses were in high demand by French forces during World War I. It was also used for bandages and other purposes.
In February 2017, researchers found that seagrass meadows may be able to remove various pathogens from seawater. On small islands without wastewater treatment facilities in central Indonesia, levels of pathogenic marine bacteria – such as Enterococcus – that affect humans, fish and invertebrates were reduced by 50 percent when seagrass meadows were present, compared to paired sites without seagrass,[49] although this could be a detriment to their survival.[50]
Movement ecology
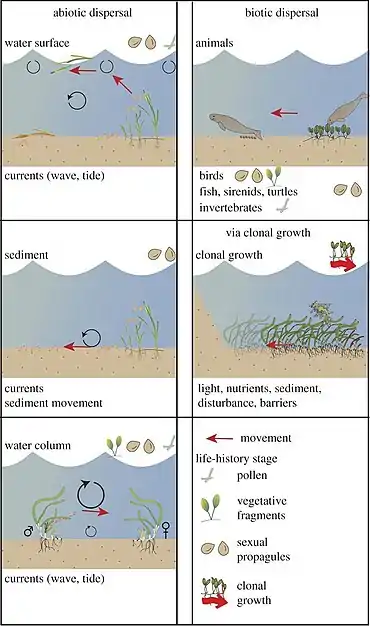
Understanding the movement ecology of seagrasses provides a way to assess the capacity of populations to recover from impacts associated with existing and future pressures. These include the (re)-colonization of altered or fragmented landscapes, and movement associated with climate change.[51]
The marine environment acts as an abiotic dispersal vector and its physical properties significantly influence movement, presenting both challenges and opportunities that differ from terrestrial environments. Typical flow speeds in the ocean are around 0.1 m s−1, generally one to two orders of magnitude weaker than typical atmospheric flows (1–10 m s−1), that can limit dispersal.[52] However, as seawater density is approximately 1000 times greater than air, momentum of a moving mass of water at the same speed is three orders of magnitude greater than in air. Therefore, drag forces acting on individuals (proportional to density) are also three orders of magnitude higher, enabling relatively larger-sized propagules to be mobilized. But most importantly, buoyancy forces (proportional to the density difference between seawater and the propagule) significantly reduce the effective weight of submerged propagules.[53] Within seagrasses, propagules can weakly settle (negatively buoyant), remain effectively suspended in the interior of the water column (neutrally buoyant), or float at the surface (positively buoyant).[54][51]
With positive buoyancy (e.g. floating fruit), ocean surface currents freely move propagules, and dispersal distances are only limited by the viability time of the fruit,[55][56] leading to exceptionally long single dispersal events (more than 100 km),[57] which is rare for passive abiotic movement of terrestrial fruit and seeds.[58][51]
There are a variety of biotic dispersal vectors for seagrasses, as they feed on or live in seagrass habitat. These include dugongs, manatees, turtles, waterfowl, fish and invertebrates.[59][60][61][62] Each biotic vector has its own internal state, motion capacity, navigation capacity and external factors influencing its movement. These interact with plant movement ecology to determine the ultimate movement path of the plant.[63][64][51]
For example, if a waterbird feeds on a seagrass containing fruit with seeds that are viable after defecation, then the bird has the potential to transport the seeds from one feeding ground to another. Therefore, the movement path of the bird determines the potential movement path of the seed. Particular traits of the animal, such as its digestive passage time, directly influence the plant's movement path.[51]
Biogeochemistry

The primary nutrients determining seagrass growth are carbon (C), nitrogen (N), phosphorus (P), and light for photosynthesis. Nitrogen and phosphorus can be acquired from sediment pore water or from the water column, and sea grasses can uptake N in both ammonium (NH4+) and nitrate (NO3−) form.[66]
A number of studies from around the world have found that there is a wide range in the concentrations of C, N, and P in seagrasses depending on their species and environmental factors. For instance, plants collected from high-nutrient environments had lower C:N and C:P ratios than plants collected from low-nutrient environments. Seagrass stoichiometry does not follow the Redfield ratio commonly used as an indicator of nutrient availability for phytoplankton growth. In fact, a number of studies from around the world have found that the proportion of C:N:P in seagrasses can vary significantly depending on their species, nutrient availability, or other environmental factors. Depending on environmental conditions, seagrasses can be either P-limited or N-limited.[67]
An early study of seagrass stoichiometry suggested that the Redfield balanced ratio between N and P for seagrasses is approximately 30:1.[68] However, N and P concentrations are strictly not correlated, suggesting that seagrasses can adapt their nutrient uptake based on what is available in the environment. For example, seagrasses from meadows fertilized with bird excrement have shown a higher proportion of phosphate than unfertilized meadows. Alternately, seagrasses in environments with higher loading rates and organic matter diagenesis supply more P, leading to N-limitation. P availability in Thalassia testudinum is the limiting nutrient. The nutrient distribution in Thalassia testudinum ranges from 29.4 to 43.3% C, 0.88-3.96% N, and 0.048-0.243% P. This equates to a mean ratio of 24.6 C:N, 937.4 C:P, and 40.2 N:P. This information can also be used to characterize the nutrient availability of a bay or other water body (which is difficult to measure directly) by sampling the seagrasses living there.[69]
Light availability is another factor that can affect the nutrient stoichiometry of seagrasses. Nutrient limitation can only occur when photosynthetic energy causes grasses to grow faster than the influx of new nutrients. For example, low light environments tend to have a lower C:N ratio.[69] Alternately, high-N environments can have an indirect negative effect to seagrass growth by promoting growth of algae that reduce the total amount of available light.[70]
Nutrient variability in seagrasses can have potential implications for wastewater management in coastal environments. High amounts of anthropogenic nitrogen discharge could cause eutrophication in previously N-limited environments, leading to hypoxic conditions in the seagrass meadow and affecting the carrying capacity of that ecosystem.[69]
A study of annual deposition of C, N, and P from Posidonia oceanica seagrass meadows in northeast Spain found that the meadow sequestered 198 g C m−2 yr−1, 13.4 g N m−2 yr−1, and 2.01 g P m−2 yr−1 into the sediment. Subsequent remineralization of carbon from the sediments due to respiration returned approximately 8% of the sequestered carbon, or 15.6 g C m−2 yr −1.[71]
 Seagrass lagoon, Chek Jawa, Singapore
Seagrass lagoon, Chek Jawa, Singapore
Threats
Seagrasses are in global decline, with some 30,000 km2 (12,000 sq mi) lost during recent decades. Seagrass loss has accelerated over the past few decades, from 0.9% per year prior to 1940 to 7% per year in 1990.[72]
Natural disturbances, such as grazing, storms, ice-scouring and desiccation, are an inherent part of seagrass ecosystem dynamics. Seagrasses display a high degree of phenotypic plasticity, adapting rapidly to changing environmental conditions. Human activities, on the other hand, have caused significant disturbance and are accountable for the majority of the losses.
The seagrass can be damaged from direct mechanical destruction of habitat through fishing methods that rely on heavy nets that are dragged across the sea floor, putting this important ecosystem at serious risk.[3] When humans drive motor boats over shallow seagrass areas, the propeller blade can also damage the seagrass.
Seagrass habitats are threatened by coastal eutrophication, which is caused by excessive input of nutrients (nitrogen, phosphorus). That excessive input is directly toxic to seagrasses, but most importantly, it stimulates the growth of epiphytic and free-floating macro- and micro-algae. Known as nuisance species, macroalgae grow in filamentous and sheet-like forms and form thick unattached mats over seagrass, occurring as epiphytes on seagrass leaves. Eutrophication leads to the forming of algal blooms, causing the attenuation of light in the water column, which eventually leads to anoxic conditions for the seagrass and organisms living in/around the plant(s). In addition to the direct blockage of light to the plant, benthic macroalgae have low carbon/nitrogen content, causing their decomposition to stimulate bacterial activity, leading to sediment resuspension, an increase in water turbidity, and further light attenuation.[73][74] When the seagrass does not get enough sunlight, it reduces the photosynthesis that nourishes the seagrass and the primary production results, and then decaying seagrass leaves and algae fuel algal blooms even further, resulting in a positive feedback loop. This can cause the decline and eradication of seagrasses to algal dominance.
Accumulating evidence also suggests that overfishing of top predators (large predatory fish) could indirectly increase algal growth by reducing grazing control performed by mesograzers, such as crustaceans and gastropods, through a trophic cascade.
Increased seawater temperatures,[10] increased sedimentation, and coastal development have also had a significant impact in the decline of seagrasses.[25]
The most-used methods to protect and restore seagrass meadows include nutrient and pollution reduction, marine protected areas, and restoration using seagrass transplanting. Seagrass is not seen as resilient to the impacts of future environmental change.[75]
Ocean deoxygenation
Globally, seagrass has been declining rapidly. Hypoxia that leads to eutrophication caused from ocean deoxygenation is one of the main underlying factors of these die-offs. Eutrophication causes enhanced nutrient enrichment which can result in seagrass productivity, but with continual nutrient enrichment in seagrass meadows, it can cause excessive growth of microalgae, epiphytes and phytoplankton resulting in hypoxic conditions.[76]
Seagrass is both a source and a sink for oxygen in the surrounding water column and sediments. At night, the inner part of seagrass oxygen pressure is linearly related to the oxygen concentration in the water column, so low water column oxygen concentrations often result in hypoxic seagrass tissues, which can eventually kill off the seagrass. Normally, seagrass sediments must supply oxygen to the below-ground tissue through either photosynthesis or by diffusing oxygen from the water column through leaves to rhizomes and roots. However, with the change in seagrass oxygen balances, it can often result in hypoxic seagrass tissues. Seagrass exposed to this hypoxic water column show increased respiration, reduced rates of photosynthesis, smaller leaves, and reduced number of leaves per shoot. This causes insufficient supply of oxygen to the belowground tissues for aerobic respiration, so seagrass must rely on the less-efficient anaerobic respiration. Seagrass die-offs create a positive feedback loop in which the mortality events cause more death as higher oxygen demands are created when dead plant material decomposes.[76]
Because hypoxia increases the invasion of sulfides in seagrass, this negatively affects seagrass through photosynthesis, metabolism and growth. Generally, seagrass is able to combat the sulfides by supplying enough oxygen to the roots. However, deoxygenation causes the seagrass to be unable to supply this oxygen, thus killing it off.[76] Deoxygenation reduces the diversity of organisms inhabiting seagrass beds by eliminating species that cannot tolerate the low oxygen conditions. Indirectly, the loss and degradation of seagrass threatens numerous species that rely on seagrass for either shelter or food. The loss of seagrass also effects the physical characteristics and resilience of seagrass ecosystems. Seagrass beds provide nursery grounds and habitat to many harvested commercial, recreational, and subsistence fish and shellfish. In many tropical regions, local people are dependent on seagrass associated fisheries as a source of food and income.[76]
Diminishing meadows
The storage of carbon is an essential ecosystem service as we move into a period of elevated atmospheric carbon levels. However, some climate change models suggest that some seagrasses will go extinct – Posidonia oceanica is expected to go extinct, or nearly so, by 2050.[77]
The UNESCO World Heritage Site around the Balearic islands of Mallorca and Formentera includes about 55,000 hectares (140,000 acres) of Posidonia oceanica, which has global significance because of the amount of carbon dioxide it absorbs. However, the meadows are being threatened by rising temperatures, which slows down its growth, as well as damage from anchors.[78]
Restoration

Using propagules
Seagrass propagules are materials that help propagate seagrass. Seagrasses pollinate by hydrophily, that is, by dispersing in the water. Sexually and asexually produced propagules are important for this dispersal.[79]
Species from the genera Amphibolis and Thalassodendron produce viviparous seedlings.[80] Most others produce seeds, although their characteristics vary widely;[81] some species produce seeds or fruit that are positively buoyant and have potential for long-distance dispersal (e.g., Enhalus, Posidonia, and Thalassia). Others produce seeds that are negatively buoyant with limited dispersal potential (e.g., Zostera and Halophila).[56][81] although long-distance dispersal can still occur via transport of detached fragments carrying spathes (modified leaves which enclose the flower cluster); e.g., Zostera spp.[82] Nearly all species are also capable of asexual reproduction through rhizome elongation [83] or the production of asexual fragments (e.g., rhizome fragments, pseudoviviparous plantlets).[84][85] Sexually derived propagules of some species lack the ability to be dormant (e.g., Amphibolis and Posidonia), while others can remain dormant for long periods.[86][87] These differences in biology and ecology of propagules strongly influence patterns of recruitment and dispersal, and the way they can be used effectively in restoration.[79]
Seagrass restoration has primarily involved using asexual material (e.g., cuttings, rhizome fragments or cores) collected from donor meadows. Relatively few seagrass restoration efforts have used sexually derived propagules.[88][89] The infrequent use of sexually derived propagules is probably in part due to the temporal and spatial variability of seed availability,[90] as well as the perception that survival rates of seeds and seedlings are poor.[91][92] Although survival rates are often low, recent reviews of seed-based research highlight that this is probably because of limited knowledge about availability and collection of quality seed, skills in seed handling and delivery, and suitability of restoration sites.[88][89][83][79]
Methods for collecting and preparing propagules vary according to their characteristics and typically harness their natural dispersal mechanisms. For example, for viviparous taxa such as Amphibolis, recently detached seedlings can be collected by placing fibrous and weighted material, such as sand-filled hessian bags, which the seedlings' grappling structures attach to as they drift past. In this way thousands of seedlings can be captured in less than a square meter.[93] Typically, sandbags are deployed in locations where restoration is required, and are not collected and re-deployed elsewhere.[79]
For species which have seeds contained within spathes (e.g., Zostera spp.), these can be harvested using divers or mechanical harvesters.[94] In Chesapeake Bay several million Zostera marina seeds have been collected each year during the peak reproductive season using a mechanical harvester.[94] Seeds are extracted from spathes after harvesting, but the methods of extraction and delivery vary. For example, some methods involve keeping the spathes within large holding tanks where they eventually split open and release the (negatively buoyant) seeds, which are then collected from the tank bottom.[94] The seeds are then placed in a flume to determine seed quality based on settling velocity, after which they are scattered by hand from boats over recipient habitats.[94] Alternatively, using buoys anchored in place, Z. marina spathes can be suspended over restoration sites in mesh bags; the spathes release and deliver the seeds to the seafloor.[95][79]
For species that release seeds from fruits that float (Posidonia spp., Halophila spp.), fruits can be detached from the parent plant by shaking; they then float to the surface where they are collected in nets.[96][97] Seeds are then extracted from the fruit via vigorous aeration and water movement from pumps at stable temperatures (25 °C) within tanks. The negatively buoyant seeds are then collected from the tank bottom and scattered by-hand over recipient habitats. Other methods have been trialed with limited success, including direct planting of seeds by hand, injecting seeds using machinery, or planting and deploying within hessian sandbags.[79]
Restoration using seagrass propagules has so far demonstrated low and variable outcomes, with more than 90% of propagules failing to survive.[98][99][93] For propagules to be successfully incorporated within seagrass restoration programs, there will need to be a reduction in propagule wastage (which includes mortality, but also failure to germinate or dispersal away from the restoration site), to facilitate higher rates of germination and survival. A major barrier to effective use of seeds in seagrass restoration is knowledge about seed quality. Seed quality includes aspects such as viability, size (which can confer energy reserves available for initial growth and establishment), damage to the seed coat or seedling, bacterial infection, genetic diversity and ecotype (which may influence a seeds ability to respond to the restoration environment).[79] Nevertheless, the diversity of propagules and species used in restoration is increasing and understanding of seagrass seed biology and ecology is advancing.[93][97][100] To improve chances of propagule establishment, better understanding is needed about the steps that precede seed delivery to restoration sites, including seed quality,[87] as well as the environmental and social barriers that influence survival and growth.[79]
Other efforts

In various locations, communities are attempting to restore seagrass beds that were lost to human action, including in the US states of Virginia,[101] Florida[102] and Hawaii,[103] as well as the United Kingdom.[104] Such reintroductions have been shown to improve ecosystem services.[105]
Dr. Fred Short of the University of New Hampshire developed a specialized transplant methodology known as "Transplanting Eelgrass Remotely with Frames" (TERF). This method involves using clusters of plants which are temporarily tied with degradable crepe paper unto a weighted frame of wire mesh. The method has already been tried out by Save The Bay.[106]
In 2001, Steve Granger, from the University of Rhode Island Graduate School of Oceanography used a boat-pulled sled that is able to deposit seeds below the sediment surface. Together with colleague Mike Traber (who developed a Knox gelatin matrix to encase the seeds in), they conducted a test planting at Narragansett Bay. They were able to plant a 400 m2 (480 sq yd) area in less than two hours.[106]
As of 2019 the Coastal Marine Ecosystems Research Centre of Central Queensland University has been growing seagrass for six years and has been producing seagrass seeds. They have been running trials in germination and sowing techniques.[107]
References
- Orth; et al. (2006). "A global crisis for seagrass ecosystems". BioScience. 56 (12): 987–996. doi:10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2. hdl:10261/88476. S2CID 4936412.
- Papenbrock, J (2012). "Highlights in seagrass' phylogeny, physiology, and metabolism: what makes them so species?". International Scholarly Research Network: 1–15.
- Fusi M and Daffonchio D (2019) "How Seagrasses Secure Our Coastlines". Frontiers for Young Minds. 7: 114. doi:10.3389/frym.2019.00114.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - Les, Donald H.; Cleland, Maryke A.; Waycott, Michelle (1997). "Phylogenetic Studies in Alismatidae, II: Evolution of Marine Angiosperms (Seagrasses) and Hydrophily". Systematic Botany. 22 (3): 443–463. doi:10.2307/2419820. JSTOR 2419820.
- Short, F.; Carruthers, T.; Dennison, W.; Waycott, M. (2007). "Global seagrass distribution and diversity: A bioregional model". Journal of Experimental Marine Biology and Ecology. 350 (1–2): 3–20. doi:10.1016/j.jembe.2007.06.012.
- Hughes, A Randall; Williams, Susan L.; Duarte, Carlos M.; Heck, Kenneth L.; Waycott, Michelle (2009). "Associations of concern: Declining seagrasses and threatened dependent species". Frontiers in Ecology and the Environment. 7 (5): 242–246. doi:10.1890/080041.
- Heck Hay, KL; Hays, G.; Orth, RJ (2003). "Critical evaluation of the nursery role hypothesis for seagrass meadows". Marine Ecology Progress Series. 253: 123–136. Bibcode:2003MEPS..253..123H. doi:10.3354/meps253123. hdl:10536/DRO/DU:30058231.
- Tomlinson and Vargo (1966). "On the morphology and anatomy of turtle grass, Thalassia testudinum (Hydrocharitaceae). I. Vegetative Morphology". Bulletin of Marine Science. 16: 748–761.
- Hogarth, Peter J. (2015). The Biology of Mangroves and Seagrasses. Oxford University Press. ISBN 9780198716556.
- Duarte, CM (2011). "Assessing the capacity of seagrass meadows for carbon burial: current limitations and future strategies". Ocean Coastal Management.
- Greiner, Jill (2013). "Seagrass restoration enhances "blue carbon" sequestration in coastal waters". PLOS ONE. 8 (8): e72469. Bibcode:2013PLoSO...872469G. doi:10.1371/journal.pone.0072469. PMC 3743776. PMID 23967303.
- Cullen-Unsworth, L.C., Jones, B.L., Lilley, R. and Unsworth, R.K. (2018) "Secret gardens under the sea: What are seagrass meadows and why are they important?" Frontiers for Young Minds, 6(2): 1–10. doi:10.3389/frym.2018.00002. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Reynolds PL (2018) "Seagrass and Seagrass Beds" Smithsonian Ocean Portal.
- McKenzie, Len J.; Nordlund, Lina M.; Jones, Benjamin L.; Cullen-Unsworth, Leanne C.; Roelfsema, Chris; Unsworth, Richard K F. (2020). "The global distribution of seagrass meadows". Environmental Research Letters. 15 (7): 074041. Bibcode:2020ERL....15g4041M. doi:10.1088/1748-9326/ab7d06. S2CID 216481192. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Heck, Kenneth L.; Carruthers, Tim J. B.; Duarte, Carlos M.; Hughes, A. Randall; Kendrick, Gary; Orth, Robert J.; Williams, Susan W. (2008). "Trophic Transfers from Seagrass Meadows Subsidize Diverse Marine and Terrestrial Consumers". Ecosystems. 11 (7): 1198–1210. doi:10.1007/s10021-008-9155-y. S2CID 7289464.
- Nordlund, Lina Mtwana; Koch, Evamaria W.; Barbier, Edward B.; Creed, Joel C. (2017). "Correction: Seagrass Ecosystem Services and Their Variability across Genera and Geographical Regions". PLOS ONE. 12 (1): e0169942. Bibcode:2017PLoSO..1269942N. doi:10.1371/journal.pone.0169942. PMC 5215874. PMID 28056075.
- Waycott, M.; Duarte, C. M.; Carruthers, T. J. B.; Orth, R. J.; Dennison, W. C.; Olyarnik, S.; Calladine, A.; Fourqurean, J. W.; Heck, K. L.; Hughes, A. R.; Kendrick, G. A.; Kenworthy, W. J.; Short, F. T.; Williams, S. L. (2009). "Accelerating loss of seagrasses across the globe threatens coastal ecosystems". Proceedings of the National Academy of Sciences. 106 (30): 12377–12381. Bibcode:2009PNAS..10612377W. doi:10.1073/pnas.0905620106. PMC 2707273. PMID 19587236.
- Dunic, Jillian C.; Brown, Christopher J.; Connolly, Rod M.; Turschwell, Mischa P.; Côté, Isabelle M. (2021). "Long‐term declines and recovery of meadow area across the world's seagrass bioregions". Global Change Biology. 27 (17): 4096–4109. doi:10.1111/gcb.15684. hdl:10072/404960. PMID 33993580. S2CID 234746753. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Röhr, Maria Emilia; et al. (2018). "Blue Carbon Storage Capacity of Temperate Eelgrass (Zostera marina) Meadows" (PDF). Global Biogeochemical Cycles. 32 (10): 1457–1475. Bibcode:2018GBioC..32.1457R. doi:10.1029/2018GB005941. S2CID 53481706.
- Maxwell, Paul S.; Eklöf, Johan S.; Van Katwijk, Marieke M.; O'Brien, Katherine R.; de la Torre-Castro, Maricela; Boström, Christoffer; Bouma, Tjeerd J.; Krause-Jensen, Dorte; Unsworth, Richard K. F.; Van Tussenbroek, Brigitta I.; Van Der Heide, Tjisse (2017). "The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems - a review". Biological Reviews. 92 (3): 1521–1538. doi:10.1111/brv.12294. PMID 27581168. S2CID 8693669.
- Unsworth, Richard K. F.; Butterworth, Emma G. (2021). "Seagrass Meadows Provide a Significant Resource in Support of Avifauna". Diversity. 13 (8): 363. doi:10.3390/d13080363. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License
- Bayraktarov, Elisa; Saunders, Megan I.; Abdullah, Sabah; Mills, Morena; Beher, Jutta; Possingham, Hugh P.; Mumby, Peter J.; Lovelock, Catherine E. (2016). "The cost and feasibility of marine coastal restoration". Ecological Applications. 26 (4): 1055–1074. doi:10.1890/15-1077. PMID 27509748.
- Hemminga, M. A., and Duarte, C. M. (2000) Seagrass Ecology, first edition, Cambridge University Press. ISBN 9780521661843.
- Gattuso, J. (2006). "Light availability in the coastal ocean: impact on the distribution of benthic photosynthetic organisms and their contribution to primary production". Biogeosciences. 3 (4): 489–513. Bibcode:2006BGeo....3..489G. doi:10.5194/bg-3-489-2006. hdl:20.500.11937/23744.
- Darnell, Kelly; Dunton, Kenneth (2016). "Reproductive phenology of the subtropical seagrasses Thalassia testudinum (Turtle grass) and Halodule wrightii (Shoal grass) in the northwest Gulf of Mexico". Botanica Marina. 59 (6): 473–483. doi:10.1515/bot-2016-0080. S2CID 88685282.
- Harvey, Chelsea (5 April 2021). "Seagrass Forests Counteract Ocean Acidification". Scientific American. Retrieved 23 November 2021.
- Ricart, Aurora M.; Ward, Melissa; Hill, Tessa M.; Sanford, Eric; Kroeker, Kristy J.; Takeshita, Yuichiro; Merolla, Sarah; Shukla, Priya; Ninokawa, Aaron T.; Elsmore, Kristen; Gaylord, Brian (2021). "Coast-wide evidence of low pH amelioration by seagrass ecosystems". Global Change Biology. 27 (11): 2580–2591. Bibcode:2021GCBio..27.2580R. doi:10.1111/gcb.15594. ISSN 1365-2486. PMC 8252054. PMID 33788362.
- Jones, Clive G.; Lawton, John H.; Shachak, Moshe (1994). "Organisms as ecosystem engineers". Oikos. 69 (3): 373–386. doi:10.2307/3545850. JSTOR 3545850.
- Unsworth, Richard K. F.; Cullen-Unsworth, Leanne C.; Jones, Benjamin L.; Lilley, Richard J. (5 August 2022). "The planetary role of seagrass conservation". Science. 377 (6606): 609–613. Bibcode:2022Sci...377..609U. doi:10.1126/science.abq6923. PMID 35926055. S2CID 251347987.
- Dineen, J. (25 July 2001). "Thalassia testudinum (Turtle grass)". Smithsonian Marine Station at Fort Pierce. Retrieved 7 November 2012.
- Macreadie, P. I.; Baird, M. E.; Trevathan-Tackett, S. M.; Larkum, A. W. D.; Ralph, P. J. (2013). "Quantifying and modelling the carbon sequestration capacity of seagrass meadows". Marine Pollution Bulletin. 83 (2): 430–439. doi:10.1016/j.marpolbul.2013.07.038. PMID 23948090.
- Nellemann, Christian et al. (2009): Blue Carbon. The Role of Healthy Oceans in Binding Carbon. A Rapid Response Assessment. Arendal, Norway: UNEP/GRID-Arendal
- National Academies Of Sciences, Engineering (2019). Negative Emissions Technologies and Reliable Sequestration: A Research Agenda. Washington, D.C.: National Academies of Sciences, Engineering, and Medicine. p. 45. doi:10.17226/25259. ISBN 978-0-309-48452-7. PMID 31120708. S2CID 134196575.
- Fourqurean, James W. (2012). "Seagrass ecosystems as a globally significant carbon stock". Nature Geoscience. 5 (7): 505–509. Bibcode:2012NatGe...5..505F. doi:10.1038/ngeo1477.
- Oreska, Matthew P. J.; McGlathery, Karen J.; Aoki, Lillian R.; Berger, Amélie C.; Berg, Peter; Mullins, Lindsay (30 April 2020). "The greenhouse gas offset potential from seagrass restoration". Scientific Reports. 10 (1): 7325. Bibcode:2020NatSR..10.7325O. doi:10.1038/s41598-020-64094-1. ISSN 2045-2322. PMC 7193639. PMID 32355280.
- Grey, William; Moffler, Mark (1987). "Flowering of the seagrass Thalassia testudinum (Hydrocharitacea) in the Tampa Bay, Florida area". Aquatic Botany. 5: 251–259. doi:10.1016/0304-3770(78)90068-2.
- Koch, E.W., Ackerman, J.D., Verduin, J. and van Keulen, M. (2007) "Fluid dynamics in seagrass ecology—from molecules to ecosystems". In" Seagrasses: biology, ecology and conservation, pages 193–225, Springer, Dordrecht. doi:10.1007/978-1-4020-2983-7_8.
- Van Veelen, Thomas J.; Karunarathna, Harshinie; Reeve, Dominic E. (2021). "Modelling wave attenuation by quasi-flexible coastal vegetation". Coastal Engineering. 164: 103820. doi:10.1016/j.coastaleng.2020.103820. S2CID 229402284. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Gregory, D., Jensen, P. and Strætkvern, K. (2012) "Conservation and in situ preservation of wooden shipwrecks from marine environments". Journal of Cultural Heritage, 13(3): S139–S148. doi:10.1016/j.culher.2012.03.005.
- Green, Andy J.; Elmberg, Johan (2014). "Ecosystem services provided by waterbirds". Biological Reviews. 89 (1): 105–122. doi:10.1111/brv.12045. PMID 23786594. S2CID 17234490.
- Kollars, Nicole M.; Henry, Amy K.; Whalen, Matthew A.; Boyer, Katharyn E.; Cusson, Mathieu; Eklöf, Johan S.; Hereu, Clara M.; Jorgensen, Pablo; Kiriakopolos, Stephanie L.; Reynolds, Pamela L.; Tomas, Fiona; Turner, Mo S.; Ruesink, Jennifer L. (2017). "Meta-Analysis of Reciprocal Linkages between Temperate Seagrasses and Waterfowl with Implications for Conservation". Frontiers in Plant Science. 8: 2119. doi:10.3389/fpls.2017.02119. PMC 5744074. PMID 29312384.
- Cury, Philippe M.; Boyd, Ian L.; Bonhommeau, Sylvain; Anker-Nilssen, Tycho; Crawford, Robert J. M.; Furness, Robert W.; Mills, James A.; Murphy, Eugene J.; Österblom, Henrik; Paleczny, Michelle; Piatt, John F.; Roux, Jean-Paul; Shannon, Lynne; Sydeman, William J. (2011). "Global Seabird Response to Forage Fish Depletion—One-Third for the Birds". Science. 334 (6063): 1703–1706. Bibcode:2011Sci...334.1703C. doi:10.1126/science.1212928. PMID 22194577. S2CID 1855657.
- Percival, S.M.; Sutherland, W.J.; Evans, P.R. (1998). "Intertidal habitat loss and wildfowl numbers: Applications of a spatial depletion model". Journal of Applied Ecology. 35: 57–63. doi:10.1046/j.1365-2664.1998.00273.x.
- Frederiksen, Morten; Edwards, Martin; Richardson, Anthony J.; Halliday, Nicholas C.; Wanless, Sarah (2006). "From plankton to top predators: Bottom-up control of a marine food web across four trophic levels". Journal of Animal Ecology. 75 (6): 1259–1268. doi:10.1111/j.1365-2656.2006.01148.x. PMID 17032358.
- Unsworth, R.K., Nordlund, L.M. and Cullen‐Unsworth, L.C. (2019) "Seagrass meadows support global fisheries production". Conservation Letters, 12(1): e12566. doi:10.1111/conl.12566. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Jones, B.L., Unsworth, R.K.F., Nordlund, L.M., Eklöf, J.S., Ambo-Rappe, R., Carly, F., Jiddawi, N.S., La Nafie, Y.A., Udagedara, S. and Cullen-Unsworth, L.C. (2022) "Dependence on seagrass fisheries governed by household income and adaptive capacity". Ocean & Coastal Management, 225. doi:10.1016/j.ocecoaman.2022.106247.
- Nessa, N., Ambo-Rappe, R., Cullen-Unsworth, L.C. and Unsworth, R.K.F. (2019) "Social-ecological drivers and dynamics of seagrass gleaning fisheries". Ambio, pages 1–11. doi:10.1007/s13280-019-01267-x. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Nordlund, L.M., Unsworth, R.K., Gullström, M. and Cullen‐Unsworth, L.C. (2018) "Global significance of seagrass fishery activity. Fish and Fisheries", 19(3): 399–412. doi:10.1111/faf.12259. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Byington, Cara (17 February 2017). "New Science Shows Seagrass Meadows Suppress Pathogens". Nature.org. NatureNet Fellows for Cool Green Science. Retrieved 17 February 2017.
- Jones, BJ; Cullen-Unsworth, L. C.; Unsworth, R. K. F. (2018). "Tracking Nitrogen Source Using δ15N Reveals Human and Agricultural Drivers of Seagrass Degradation across the British Isles". Frontiers in Plant Science. 9: 133. doi:10.3389/fpls.2018.00133. PMC 5808166. PMID 29467789.
- McMahon, Kathryn; Van Dijk, Kor-Jent; Ruiz-Montoya, Leonardo; Kendrick, Gary A.; Krauss, Siegfried L.; Waycott, Michelle; Verduin, Jennifer; Lowe, Ryan; Statton, John; Brown, Eloise; Duarte, Carlos (2014). "The movement ecology of seagrasses". Proceedings of the Royal Society B: Biological Sciences. 281 (1795). doi:10.1098/rspb.2014.0878. PMC 4213608. PMID 25297859. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Denny, Mark (1993). Air and Water: The Biology and Physics of Life's Media. Princeton University Press. ISBN 0691025185.
- Vogel, Steven (5 May 2020). Life in Moving Fluids: The Physical Biology of Flow - Revised and Expanded Second Edition. Princeton University Press. ISBN 9780691212975.
- Larkum, Anthony W. D.; Orth, Robert J.; Duarte, Carlos (16 May 2007). Seagrasses: Biology, Ecology and Conservation. Springer. ISBN 9781402029837.
- Van Dijk, JK; Van Tussenbroek, BI; Jiménez-Durán, K.; Márquez-Guzmán, GJ; Ouborg, J. (2009). "High levels of gene flow and low population genetic structure related to high dispersal potential of a tropical marine angiosperm". Marine Ecology Progress Series. 390: 67–77. Bibcode:2009MEPS..390...67V. doi:10.3354/meps08190. hdl:2066/75120.
- Lacap, CDA; Vermaat, JE; Rollon, RN; Nacorda, HM (2002). "Propagule dispersal of the SE Asian seagrasses Enhalus acoroides and Thalassia hemprichii". Marine Ecology Progress Series. 235: 75–80. Bibcode:2002MEPS..235...75L. doi:10.3354/meps235075.
- Ruiz-Montoya, L.; Lowe, R. J.; Van Niel, K. P.; Kendrick, G. A. (2012). "The role of hydrodynamics on seed dispersal in seagrasses". Limnology and Oceanography. 57 (5): 1257–1265. Bibcode:2012LimOc..57.1257R. doi:10.4319/lo.2012.57.5.1257.
- Maurer, Kyle D.; Bohrer, Gil; Medvigy, David; Wright, S. Joseph (2013). "The timing of abscission affects dispersal distance in a wind-dispersed tropical tree". Functional Ecology. 27: 208–218. doi:10.1111/1365-2435.12028.
- Van Tussenbroek, BI; Monroy-Velazquez, LV; Solis-Weiss, V. (2012). "Meso-fauna foraging on seagrass pollen may serve in marine zoophilous pollination". Marine Ecology Progress Series. 469: 1–6. Bibcode:2012MEPS..469....1V. doi:10.3354/meps10072.
- Figuerola, Jordi; Green, Andy J.; Santamaria, Luis (2002). "Comparative dispersal effectiveness of wigeongrass seeds by waterfowl wintering in south-west Spain: Quantitative and qualitative aspects". Journal of Ecology. 90 (6): 989–1001. doi:10.1046/j.1365-2745.2002.00734.x. hdl:10261/43047. S2CID 55550198.
- Sumoski, SE; Orth, RJ (2012). "Biotic dispersal in eelgrass Zostera marina". Marine Ecology Progress Series. 471: 1–10. Bibcode:2012MEPS..471....1S. doi:10.3354/meps10145.
- Sheppard, James K.; Marsh, Helene; Jones, Rhondda E.; Lawler, Ivan R. (2010). "Dugong habitat use in relation to seagrass nutrients, tides, and diel cycles". Marine Mammal Science. 26 (4): 855–879. doi:10.1111/j.1748-7692.2010.00374.x.
- Damschen, E. I.; Brudvig, L. A.; Haddad, N. M.; Levey, D. J.; Orrock, J. L.; Tewksbury, J. J. (2008). "The movement ecology and dynamics of plant communities in fragmented landscapes". Proceedings of the National Academy of Sciences. 105 (49): 19078–19083. doi:10.1073/pnas.0802037105. PMC 2614718. PMID 19060187.
- Tsoar A, Shohami D and Nathan R (2011). "A movement ecology approach to study seed dispersal and plant invasion: an overview and application of seed dispersal by fruit bats". Fifty years of invasion ecology, the legacy of Charles Elton (Ed. D M Richardson), pp. 103–120. Centre for Invasion Biology, Department of Botany and Zoology, Stellenbosch University, Wiley Blackwell.
- Brodersen, K.E., Koren, K., Moßhammer, M., Ralph, P.J., Kühl, M. and Santner, J. (2017) "Seagrass-mediated phosphorus and iron solubilization in tropical sediments". Environmental science & technology, 51(24): 14155–14163. doi:10.1021/acs.est.7b03878. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Chung, I. K.; Beardall, J.; Mehta, S.; Sahoo, D.; Stojkovic, S. (2011). "Using marine macroalgae for carbon sequestration: a critical appraisal". Journal of Applied Phycology. 23 (5): 877–886. doi:10.1007/s10811-010-9604-9. S2CID 45039472.
- Fourqurean, James W.; Zieman, Joseph C.; Powell, George V. N. (1992). "Phosphorus Limitation of Primary Production in Florida Bay: Evidence from C:N:P Ratios of the Dominant Seagrass Thalassia Testudinum". Limnology and Oceanography. 37 (1): 162–71. Bibcode:1992LimOc..37..162F. doi:10.4319/lo.1992.37.1.0162.
- Mcleod, E.; Chmura, G. L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C. M.; Silliman, B. R. (2011). "A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2" (PDF). Frontiers in Ecology and the Environment. 9 (10): 552–560. doi:10.1890/110004.
- Fourqurean, James W.; Zieman, Joseph C. (2002). "Nutrient Content of the Seagrass Thalassia Testudinum Reveals Regional Patterns of Relative Availability of Nitrogen and Phosphorus in the Florida Keys USA". Biogeochemistry. 61 (3): 229–45. doi:10.1023/A:1020293503405. S2CID 4948302.
- Chmura, Gail; Anisfield, Shimon (2003). "Global carbon sequestration in tidal, saline wetland soils". Global Biogeochemical Cycles. 17 (4): n/a. Bibcode:2003GBioC..17.1111C. doi:10.1029/2002GB001917.
- Chi, Z.; Xie, Y.; Elloy, F.; Zheng, Y.; Hu, Y.; Chen, S. (2013). "Bicarbonate-based integrated carbon capture and algae production system with alkalihalophilic cyanobacterium". Bioresource Technology. 133: 513–521. doi:10.1016/j.biortech.2013.01.150. PMID 23455223.
- Waycott, M (2009). "Accelerating loss of seagrasses across the globe threatens coastal ecosystems". Proceedings of the National Academy of Sciences of the USA. 106 (30): 12377–12381. Bibcode:2009PNAS..10612377W. doi:10.1073/pnas.0905620106. PMC 2707273. PMID 19587236.
- McGlathery, KJ (2001). "Macroalgal blooms contribute to the decline of seagrass in nutrient‐enriched coastal waters" (PDF). Journal of Phycology. 37 (4): 453–456. doi:10.1046/j.1529-8817.2001.037004453.x. S2CID 38983997.
- Fox SE, YS Olsen and AC Spivak (2010) "Effects of bottom-up and top-down controls and climate change on estuarine macrophyte communities and the ecosystem services they provide" In: PF Kemp (Ed) Eco-DAS Symposium Proceedings, ALSO, Chapter 8: 129–145.
- Unsworth, Richard K.F.; Collier, Catherine J.; Waycott, Michelle; McKenzie, Len J.; Cullen-Unsworth, Leanne C. (2015). "A framework for the resilience of seagrass ecosystems". Marine Pollution Bulletin. 100 (1): 34–46. Bibcode:2015MarPB.100...34U. doi:10.1016/j.marpolbul.2015.08.016. PMID 26342389.
- Laffoley, D. & Baxter, J.M. (eds.) (2019). Ocean deoxygenation: Everyone’s problem - Causes, impacts, consequences and solutions. IUCN, Switzerland.
- Jordà, Gabriel; Marbà, Núria; Duarte, Carlos M. (2012). "Mediterranean seagrass vulnerable to regional climate warming". Nature Climate Change. 2 (11): 821–824. Bibcode:2012NatCC...2..821J. doi:10.1038/nclimate1533. ISSN 1758-6798.
- McGrath, Matt (13 March 2021). "Climate change: 'Forever plant' seagrass faces uncertain future". BBC News. Retrieved 20 April 2021.
- Vanderklift, Mathew A.; Doropoulos, Christopher; Gorman, Daniel; Leal, Inês; Minne, Antoine J. P.; Statton, John; Steven, Andrew D. L.; Wernberg, Thomas (2020). "Using Propagules to Restore Coastal Marine Ecosystems". Frontiers in Marine Science. 7. doi:10.3389/fmars.2020.00724. S2CID 221667138. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Kuo, J.; Hartog, C. den (2006). "Seagrass Morphology, Anatomy, and Ultrastructure". Seagrasses: Biology, Ecologyand Conservation. pp. 51–87. doi:10.1007/978-1-4020-2983-7_3. ISBN 978-1-4020-2942-4.
- Kendrick, Gary A.; Waycott, Michelle; Carruthers, Tim J. B.; Cambridge, Marion L.; Hovey, Renae; Krauss, Siegfried L.; Lavery, Paul S.; Les, Donald H.; Lowe, Ryan J.; Vidal, Oriol Mascaró i.; Ooi, Jillian L. S.; Orth, Robert J.; Rivers, David O.; Ruiz-Montoya, Leonardo; Sinclair, Elizabeth A.; Statton, John; Van Dijk, Jent Kornelis; Verduin, Jennifer J. (2012). "The Central Role of Dispersal in the Maintenance and Persistence of Seagrass Populations". BioScience. 62: 56–65. doi:10.1525/bio.2012.62.1.10. S2CID 4632119.
- Harwell, Matthew C.; Orth, Robert J. (2002). "Long-Distance Dispersal Potential in a Marine Macrophyte". Ecology. 83 (12): 3319–3330. doi:10.1890/0012-9658(2002)083[3319:lddpia]2.0.co;2.
- Kendrick, Gary A.; Orth, Robert J.; Statton, John; Hovey, Renae; Ruiz Montoya, Leonardo; Lowe, Ryan J.; Krauss, Siegfried L.; Sinclair, Elizabeth A. (2017). "Demographic and genetic connectivity: The role and consequences of reproduction, dispersal and recruitment in seagrasses". Biological Reviews. 92 (2): 921–938. doi:10.1111/brv.12261. PMID 27010433. S2CID 3470879.
- Ballesteros, Enric; Cebrian, Emma; Garcia-Rubies, Antoni; Alcoverro, Teresa; Romero, Javier; Font, Xavier (2005). "Pseudovivipary, a new form of asexual reproduction in the seagrass Posidonia oceanica". Botanica Marina. 48 (2). doi:10.1515/BOT.2005.026. hdl:10261/2993. S2CID 84864719.
- Sinclair, Elizabeth A.; Statton, John; Hovey, Renae; Anthony, Janet M.; Dixon, Kingsley W.; Kendrick, Gary A. (2015). "Reproduction at the extremes: Pseudovivipary, hybridization and genetic mosaicism in Posidonia australis(Posidoniaceae)". Annals of Botany. 117 (2): 237–47. doi:10.1093/aob/mcv162. PMC 4724040. PMID 26578720.
- Inglis, Graeme J. (2000). "Disturbance-related heterogeneity in the seed banks of a marine angiosperm". Journal of Ecology. 88: 88–99. doi:10.1046/j.1365-2745.2000.00433.x. S2CID 84928065.
- Orth, RJ; Harwell, MC; Bailey, EM; Bartholomew, A.; Jawad, JT; Lombana, AV; Moore, KA; Rhode, JM; Woods, HE (2000). "A review of issues in seagrass seed dormancy and germination:implications for conservation and restoration". Marine Ecology Progress Series. 200: 277–288. Bibcode:2000MEPS..200..277O. doi:10.3354/meps200277.
- Statton, John; Dixon, Kingsley W.; Hovey, Renae K.; Kendrick, Gary A. (2012). "A comparative assessment of approaches and outcomes for seagrass revegetation in Shark Bay and Florida Bay". Marine and Freshwater Research. 63 (11): 984. doi:10.1071/mf12032.
- Van Katwijk, Marieke M.; Thorhaug, Anitra; Marbà, Núria; Orth, Robert J.; Duarte, Carlos M.; Kendrick, Gary A.; Althuizen, Inge H. J.; Balestri, Elena; Bernard, Guillaume; Cambridge, Marion L.; Cunha, Alexandra; Durance, Cynthia; Giesen, Wim; Han, Qiuying; Hosokawa, Shinya; Kiswara, Wawan; Komatsu, Teruhisa; Lardicci, Claudio; Lee, Kun-Seop; Meinesz, Alexandre; Nakaoka, Masahiro; O'Brien, Katherine R.; Paling, Erik I.; Pickerell, Chris; Ransijn, Aryan M. A.; Verduin, Jennifer J. (2016). "Global analysis of seagrass restoration: The importance of large-scale planting". Journal of Applied Ecology. 53 (2): 567–578. doi:10.1111/1365-2664.12562.
- Orth, Robert J.; Luckenbach, Mark L.; Marion, Scott R.; Moore, Kenneth A.; Wilcox, David J. (2006). "Seagrass recovery in the Delmarva Coastal Bays, USA". Aquatic Botany. 84: 26–36. doi:10.1016/j.aquabot.2005.07.007.
- Fonseca, M. S. (1998). Guidelines for the Conservation and Restoration of Seagrasses in the United States and Adjacent Waters, US Department of Commerce, Washington, DC.
- Kirkman, Hugh (1999). "Pilot Experiments on Planting Seedlings and Small Seagrass Propagules in Western Australia". Marine Pollution Bulletin. 37 (8–12): 460–467. Bibcode:1999MarPB..37..460K. doi:10.1016/s0025-326x(99)00146-0.
- Tanner, Jason E. (2015). "Restoration of the Seagrass Amphibolis antarctica—Temporal Variability and Long-Term Success". Estuaries and Coasts. 38 (2): 668–678. doi:10.1007/s12237-014-9823-4. S2CID 88784518.
- Orth, RJ; Moore, KA; Marion, SR; Wilcox, DJ; Parrish, DB (2012). "Seed addition facilitates eelgrass recovery in a coastal bay system". Marine Ecology Progress Series. 448: 177–195. Bibcode:2012MEPS..448..177O. doi:10.3354/meps09522.
- Pickerell, Christopher H.; Schott, Stephen; Wyllie-Echeverria, Sandy (2005). "Buoy-deployed seeding: Demonstration of a new eelgrass (Zostera marina L.) planting method". Ecological Engineering. 25 (2): 127–136. doi:10.1016/j.ecoleng.2005.03.005.
- Statton, John; Cambridge, Marion L.; Dixon, Kingsley W.; Kendrick, Gary A. (2013). "Aquaculture of Posidonia australis Seedlings for Seagrass Restoration Programs: Effect of Sediment Type and Organic Enrichment on Growth". Restoration Ecology. 21 (2): 250–259. doi:10.1111/j.1526-100x.2012.00873.x. S2CID 84428852.
- Statton, John; Montoya, Leonardo R.; Orth, Robert J.; Dixon, Kingsley W.; Kendrick, Gary A. (2017). "Identifying critical recruitment bottlenecks limiting seedling establishment in a degraded seagrass ecosystem". Scientific Reports. 7 (1): 14786. Bibcode:2017NatSR...714786S. doi:10.1038/s41598-017-13833-y. PMC 5665928. PMID 29093460.
- Orth, RJ; Fishman, JR; Harwell, MC; Marion, SR (2003). "Seed-density effects on germination and initial seedling establishment in eelgrass Zostera marina in the Chesapeake Bay region". Marine Ecology Progress Series. 250: 71–79. Bibcode:2003MEPS..250...71O. doi:10.3354/meps250071.
- Orth, R. J.; Marion, S. R.; Moore, K. A. (2007). "A Summary of Eelgrass (Zostera marina) Reproductive Biology with an Emphasis on Seed Biology and Ecology from the Chesapeake Bay Region" (PDF). U.S. Army Corps of Engineers.
{{cite journal}}: Cite journal requires|journal=(help) - Strydom, Simone; McMahon, Kathryn; Lavery, Paul S. (2017). "Response of the seagrass Halophila ovalis to altered light quality in a simulated dredge plume". Marine Pollution Bulletin. 121 (1–2): 323–330. Bibcode:2017MarPB.121..323S. doi:10.1016/j.marpolbul.2017.05.060. PMID 28625617.
- "Eelgrass Restoration | The Nature Conservancy in Virginia". www.nature.org. Retrieved 6 August 2018.
- "Seagrass Restoration". myfwc.com. Retrieved 6 August 2018.
- "Seagrass Restoration Initiative – Malama Maunalua". www.malamamaunalua.org. Retrieved 6 August 2018.
- Unsworth, Richard K. F.; McKenzie, Len J.; Collier, Catherine J.; Cullen-Unsworth, Leanne C.; Duarte, Carlos M.; Eklöf, Johan S.; Jarvis, Jessie C.; Jones, Benjamin L.; Nordlund, Lina M. (1 August 2019). "Global challenges for seagrass conservation". Ambio. 48 (8): 801–815. doi:10.1007/s13280-018-1115-y. ISSN 1654-7209. PMC 6541581. PMID 30456457.
- van Katwijk, Marieke M.; Thorhaug, Anitra; Marbà, Núria; Orth, Robert J.; Duarte, Carlos M.; Kendrick, Gary A.; Althuizen, Inge H. J.; Balestri, Elena; Bernard, Guillaume (25 November 2015). "Global analysis of seagrass restoration: the importance of large-scale planting". Journal of Applied Ecology. 53 (2): 567–578. doi:10.1111/1365-2664.12562. ISSN 0021-8901.
- Restoration Methods
- Mackay, Jacquie; Stünzner, Inga (24 October 2019). "Seagrass nursery in central Queensland could offset carbon emissions". ABC News. Australian Broadcasting Corporation. Retrieved 24 October 2019.
One flower can produce 15 seeds, and one seed planted in the right conditions can create a hectare of seagrass.