Steroid Delta-isomerase
In enzymology, a steroid Δ5-isomerase (EC 5.3.3.1) is an enzyme that catalyzes the chemical reaction
- a 3-oxo-Δ5-steroid a 3-oxo-Δ4-steroid

| Steroid Δ-isomerase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
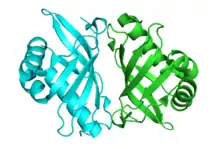 Crystallographic structure of Pseudomonas putida steroid Δ5-isomerase homodimer.[1] | |||||||||
| Identifiers | |||||||||
| EC no. | 5.3.3.1 | ||||||||
| CAS no. | 9031-36-1 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
Hence, this enzyme has one substrate, a 3-oxo-Δ5-steroid, and one product, a 3-oxo-Δ4-steroid.
Introduction
This enzyme belongs to the family of isomerases, specifically those intramolecular oxidoreductases transposing C=C bonds. The systematic name of this enzyme class is 3-oxosteroid Δ5-Δ4-isomerase. Other names in common use include ketosteroid isomerase (KSI), hydroxysteroid isomerase, steroid isomerase, Δ5-ketosteroid isomerase, Δ5(or Δ4)-3-keto steroid isomerase, Δ5-steroid isomerase, 3-oxosteroid isomerase, Δ5-3-keto steroid isomerase, and Δ5-3-oxosteroid isomerase.
KSI has been studied extensively from the bacteria Comamonas testosteroni (TI), formerly referred to as Pseudomonas testosteroni, and Pseudomonas putida (PI).[2] The enzymes from these two sources are 34% homologous, and structural studies have shown that the placement of the catalytic groups in the active sites is virtually identical.[3] Mammalian KSI has been studied from bovine adrenal cortex[4] and rat liver.[5] This enzyme participates in c21-steroid hormone metabolism and androgen and estrogen metabolism. An example substrate is Δ5-androstene-3,17-dione, which KSI converts to Δ4-androstene-3,17-dione.[6] The above reaction in the absence of enzyme takes 7 weeks to complete in aqueous solution.[7] KSI performs this reaction on an order of 1011 times faster, ranking it among the most proficient enzymes known.[7] Bacterial KSI also serves as a model protein for studying enzyme catalysis[8] and protein folding.[9]
Structural studies
KSI exists as a homodimer with two identical halves.[9] The interface between the two monomers is narrow and well defined, consisting of neutral or apolar amino acids, suggesting the hydrophobic interaction is important for dimerization.[9] Results show that the dimerization is essential to function.[9] The active site is highly apolar and folds around the substrate in a manner similar to other enzymes with hydrophobic substrates, suggesting this fold is characteristic for binding hydrophobic substrates.[10]
No complete atomic structure of KSI appeared until 1997, when an NMR structure of TI KSI was reported.[11] This structure showed that the active site is a deep hydrophobic pit with Asp-38 and Tyr-14 located at the bottom of this pit.[11] The structure is thus entirely consistent with the proposed mechanistic roles of Asp-38 and Tyr-14.
| Residue Role | Comamonas testosteroni (PDB: 8CHO) | Pseudomonas putida (PDB: 1OH0) |
|---|---|---|
| Oxyanion H-Bond Donor(s) | Asp-99 | Asp-103 |
| Tyr-14 | Tyr-16 | |
| General Acid/Base | Asp-38 | Asp-40 |
As of late 2007, 25 structures have been solved for this class of enzymes, with PDB accession codes 1BUQ, 1C7H, 1CQS, 1DMM, 1DMN, 1DMQ, 1E97, 1GS3, 1ISK, 1K41, 1OCV, 1OGX, 1OGZ, 1OH0, 1OHO, 1OHP, 1OHS, 1OPY, 1VZZ, 1W00, 1W01, 1W02, 1W6Y, 2PZV, and 8CHO.
Mechanism
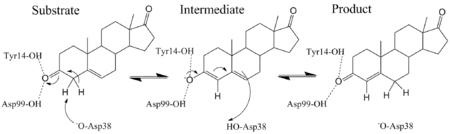
KSI catalyzes the rearrangement of a carbon-carbon double bond in ketosteroids through an enolate intermediate at a diffusion-limited rate.[2] There have been conflicting results on the ionization state of the intermediate, whether it exists as the enolate[12] or enol.[13] Pollack uses a thermodynamic argument to suggest the intermediate exists as the enolate.[2] The general base Asp-38 abstracts a proton from position 4 (alpha to the carbonyl, next to the double bond) of the steroid ring to form an enolate (the rate-limiting step)[14] that is stabilized by the hydrogen bond donating Tyr-14 and Asp-99.[2] Tyr-14 and Asp-99 are positioned deep within the hydrophobic active site and form a so-called oxanion hole.[15] Protonated Asp-38 then transfers its proton to position 6 of the steroid ring to complete the reaction.[2]
Although the mechanistic steps of the reaction are not disputed, the contributions of various factors to catalysis such as electrostatics, hydrogen bonding of the oxyanion hole, and distal binding effects are discussed below and still debated.
The Warshel group applied statistical mechanical computational methods and empirical valence bond theory to previous experimental data. It was determined that electrostatic preorganization-including ionic residues and fixed dipoles within the active site-contributes most to KSI catalysis.[16] More specifically, Tyr-14 and Asp-99 dipoles work to stabilize the growing charge which accumulates on the enolate oxygen (O-3) throughout catalysis. In a similar way, the charge on Asp38 is stabilized by surrounding residues and a water molecule during the course of the reaction.[16] The Boxer group used experimental Stark spectroscopy methods to identify the presence of H-bond-mediated electric fields within the KSI active site. These measurements quantified the electrostatic contribution to KSI catalysis (70%).[17]
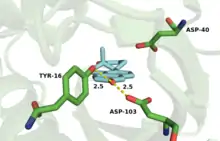
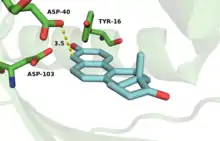
The active site is lined with hydrophobic residues to accommodate the substrate, but Asp-99 and Tyr-14 are within hydrogen bonding distance of O-3.[18] The hydrogen bonds from Tyr-14 and Asp-99 are known to significantly affect the rate of catalysis in KSI.[2] Mutagenesis of this residue to alanine (D99A) or asparagine (D99N) results in a loss in activity at pH 7 of 3000-fold and 27-fold, respectively,[11][19] implicating Asp-99 as important for enzymatic activity. Wu et al.[11] proposed a mechanism that involves both Tyr-14 and Asp-99 forming hydrogen bonds directly to O-3 of the steroid. This mechanism was challenged by Zhao et al.,[20] who postulated a hydrogen bonding network with Asp-99 hydrogen bonding to Tyr-14, which in turn forms a hydrogen bond to O-3. More recently, the Herschlag group utilized unnatural amino acid incorporation to assay the importance of Tyr-14 to KSI catalysis.[21] The natural tyrosine residue was substituted with unnatural halogenated amino acids surveying a range of pKa's. There was very little difference in KSI catalytic turnover with decreasing pKa, suggesting, in contrast to the electrostatic studies outlined above, that oxyanion hole stabilization is not primarily important for catalysis.[21]
| Wild-Type KSI Reaction Kinetics on 5-Androstenedione[22] | |
|---|---|
| kcat (s−1) | 3.0 x 104 |
| Km (μM) | 123 |
| kcat/Km (M−1s−1) | 2.4 x 108 |
Asp-38 general acidic/basic activity and effective molarity was probed by the Herschlag group through site-directed mutagenesis and exogenous base rescue.[23] Asp-38 was mutated to Gly, nullifying catalytic activity, and exogenous rescue was attempted with carboxylates of varying size and molarity. By calculating the concentration of base needed for full rescue, the Herschlag group determined the effective molarity of Asp-38 in KSI (6400 M). Thus, Asp-38 is critical for KSI catalysis.[23]
Sigala et al. found that solvent exclusion and replacement by the remote hydrophobic steroid rings negligibly alter the electrostatic environment within the KSI oxyanion hole.[24] In addition, ligand binding does not grossly alter the conformations of backbone and side chain groups observed in X-ray structures of PI KSI. However, NMR and UV studies suggest that steroid binding restricts the motions of several active-site groups, including Tyr-16.[25][26] Recently, the Herschlag group proposed that remote binding of hydrophobic regions of the substrate to distal portions of the active site contribute to KSI catalysis (>5 kcal/mol).[27] A 4-ring substrate reacted 27,000 times faster than a single ring substrate indicating the importance of distal active site binding motifs. This activity ratio persists throughout mutagenesis of residues important to oxyanion hole stabilization, implying that distal binding is what accounts for the large aforementioned reactivity difference.[27]
Numerous physical changes occur upon steroid binding within the KSI active site. In the free enzyme an ordered water molecule is positioned within hydrogen-bonding distance of Tyr-16 (the PI equivalent of TI KSI Tyr-14) and Asp-103 (the PI equivalent of TI KSI Asp-99).[28] This and additional disordered water molecules present within the unliganded active site are displaced upon steroid binding and are substantially excluded by the dense constellation of hydrophobic residues that pack around the bound, hydrophobic steroid skeleton.[28][25]
As stated above, the degree to which various factors contribute to KSI catalysis is still debated.
Function
KSI occurs in animal tissues concerned with steroid hormone biosynthesis, such as the adrenal, testis, and ovary.[29] KSI in Comamomas testosteroni is used in the degradation pathway of steroids, allowing this bacteria to utilize steroids containing a double bond at Δ5, such as testosterone, as its sole source of carbon.[30] In mammals, transfer of a double bond at Δ5 to Δ4 is catalyzed by 3-β-hydroxy-Δ5-steroid dehydrogenase at the same time as the dehydroxylation of 3-β-hydroxyl group to ketone group,[31] while in C. testosteroni and P. putida, Δ5,3-ketosteroid isomerase just transfers a double bond at Δ5 of 3-ketosteroid to Δ4.[32]
A Δ5-3-ketosteroid isomerase-disrupted mutant of strain TA441 can grow on dehydroepiandrosterone, which has a double bond at Δ5, but cannot grow on epiandrosterone, which lacks a double bond at Δ5, indicating that C. testosteroni KSI is responsible for transfer of the double bond from Δ5 to Δ4 and transfer of the double bond by hydrogenation at Δ5 and following dehydrogenation at Δ4 is not possible.[33]
Model enzyme
KSI has been used as a model system to test different theories to explain how enzymes achieve their catalytic efficiency. Low-barrier hydrogen bonds and unusual pKa values for the catalytic residues have been proposed as the basis for the fast action of KSI.[10][15] Gerlt and Gassman proposed the formation of unusually short, strong hydrogen bonds between KSI oxanion hole and the reaction intermediate as a means of catalytic rate enhancement.[34][35] In their model, high-energy states along the reaction coordinate are specifically stabilized by the formation of these bonds. Since then, the catalytic role of short, strong hydrogen bonds has been debated.[36][37] Another proposal explaining enzyme catalysis tested through KSI is the geometrical complementarity of the active site to the transition state, which proposes the active site electrostatics is complementary to the substrate transition state.[8]
KSI has also been a model system for studying protein folding. Kim et al. studied the effect of folding and tertiary structure on the function of KSI.[9]
References
- PDB: 3VSY; Kobe A, Caaveiro JM, Tashiro S, Kajihara D, Kikkawa M, Mitani T, Tsumoto K (March 2013). "Incorporation of rapid thermodynamic data in fragment-based drug discovery". Journal of Medicinal Chemistry. 56 (5): 2155–9. doi:10.1021/jm301603n. PMID 23419007.
- Pollack RM (October 2004). "Enzymatic mechanisms for catalysis of enolization: ketosteroid isomerase". Bioorganic Chemistry. 32 (5): 341–53. doi:10.1016/j.bioorg.2004.06.005. PMID 15381400.
- Cho HS, Choi G, Choi KY, Oh BH (June 1998). "Crystal structure and enzyme mechanism of Delta 5-3-ketosteroid isomerase from Pseudomonas testosteroni". Biochemistry. 37 (23): 8325–30. doi:10.1021/bi9801614. PMID 9622484.
- Bertolino A, Benson AM, Talalay P (June 1979). "Activation of delta5-3-ketosteroid isomerase of bovine adrenal microsomes by serum albumins". Biochemical and Biophysical Research Communications. 88 (3): 1158–66. doi:10.1016/0006-291X(79)91530-4. PMID 465075.
- Benson AM, Talalay P (April 1976). "Role of reduced glutathione in the delta(5)-3-kitosteroid isomerase reaction of liver". Biochemical and Biophysical Research Communications. 69 (4): 1073–9. doi:10.1016/0006-291X(76)90482-4. PMID 6023.
- Talalay P, Benson AM (1972). "Δ5-3-Ketosteroid Isomerase". In Boyer PD (ed.). The Enzymes. Vol. 6 (3rd ed.). Academic Press. pp. 591–618. ISBN 978-0-12-122706-7.
- Radzicka A, Wolfenden R (January 1995). "A proficient enzyme". Science. 267 (5194): 90–3. Bibcode:1995Sci...267...90R. doi:10.1126/science.7809611. PMID 7809611.
- Kraut DA, Sigala PA, Pybus B, Liu CW, Ringe D, Petsko GA, Herschlag D (April 2006). "Testing electrostatic complementarity in enzyme catalysis: hydrogen bonding in the ketosteroid isomerase oxyanion hole". PLOS Biology. 4 (4): e99. doi:10.1371/journal.pbio.0040099. PMC 1413570. PMID 16602823.

- Kim DH, Nam GH, Jang DS, Yun S, Choi G, Lee HC, Choi KY (April 2001). "Roles of dimerization in folding and stability of ketosteroid isomerase from Pseudomonas putida biotype B". Protein Science. 10 (4): 741–52. doi:10.1110/ps.18501. PMC 2373975. PMID 11274465.
- Ha NC, Kim MS, Lee W, Choi KY, Oh BH (December 2000). "Detection of large pKa perturbations of an inhibitor and a catalytic group at an enzyme active site, a mechanistic basis for catalytic power of many enzymes". The Journal of Biological Chemistry. 275 (52): 41100–6. doi:10.1074/jbc.M007561200. PMID 11007792.
- Wu ZR, Ebrahimian S, Zawrotny ME, Thornburg LD, Perez-Alvarado GC, Brothers P, Pollack RM, Summers MF (April 1997). "Solution structure of 3-oxo-delta5-steroid isomerase". Science. 276 (5311): 415–8. doi:10.1126/science.276.5311.415. PMID 9103200.
- Xue LA, Kuliopulos A, Mildvan AS, Talalay P (May 1991). "Catalytic mechanism of an active-site mutant (D38N) of delta 5-3-ketosteroid isomerase. Direct spectroscopic evidence for dienol intermediates". Biochemistry. 30 (20): 4991–7. doi:10.1021/bi00234a022. PMID 2036366.
- Petrounia IP, Pollack RM (January 1998). "Substituent effects on the binding of phenols to the D38N mutant of 3-oxo-delta5-steroid isomerase. A probe for the nature of hydrogen bonding to the intermediate". Biochemistry. 37 (2): 700–5. doi:10.1021/bi972262s. PMID 9425094.
- Wu Y, Boxer SG (September 2016). "A Critical Test of the Electrostatic Contribution to Catalysis with Noncanonical Amino Acids in Ketosteroid Isomerase". Journal of the American Chemical Society. 138 (36): 11890–5. doi:10.1021/jacs.6b06843. PMC 5063566. PMID 27545569.
- Childs W, Boxer SG (March 2010). "Proton affinity of the oxyanion hole in the active site of ketosteroid isomerase". Biochemistry. 49 (12): 2725–31. doi:10.1021/bi100074s. PMC 2852583. PMID 20143849.
- Kamerlin SC, Sharma PK, Chu ZT, Warshel A (March 2010). "Ketosteroid isomerase provides further support for the idea that enzymes work by electrostatic preorganization". Proceedings of the National Academy of Sciences of the United States of America. 107 (9): 4075–80. Bibcode:2010PNAS..107.4075K. doi:10.1073/pnas.0914579107. PMC 2840163. PMID 20150513.
- Fried SD, Bagchi S, Boxer SG (December 2014). "Extreme electric fields power catalysis in the active site of ketosteroid isomerase". Science. 346 (6216): 1510–4. Bibcode:2014Sci...346.1510F. doi:10.1126/science.1259802. PMC 4668018. PMID 25525245.
- Kim SW, Cha SS, Cho HS, Kim JS, Ha NC, Cho MJ, Joo S, Kim KK, Choi KY, Oh BH (November 1997). "High-resolution crystal structures of delta5-3-ketosteroid isomerase with and without a reaction intermediate analogue". Biochemistry. 36 (46): 14030–6. doi:10.1021/bi971546+. PMID 9369474.
- Thornburg LD, Hénot F, Bash DP, Hawkinson DC, Bartel SD, Pollack RM (July 1998). "Electrophilic assistance by Asp-99 of 3-oxo-Delta 5-steroid isomerase". Biochemistry. 37 (29): 10499–506. doi:10.1021/bi980099a. PMID 9671521.
- Zhao Q, Abeygunawardana C, Gittis AG, Mildvan AS (December 1997). "Hydrogen bonding at the active site of delta 5-3-ketosteroid isomerase". Biochemistry. 36 (48): 14616–26. doi:10.1021/bi971549m. PMID 9398180.
- Natarajan A, Schwans JP, Herschlag D (May 2014). "Using unnatural amino acids to probe the energetics of oxyanion hole hydrogen bonds in the ketosteroid isomerase active site". Journal of the American Chemical Society. 136 (21): 7643–54. doi:10.1021/ja413174b. PMC 4046884. PMID 24787954.
- Holman CM, Benisek WF (October 1995). "Insights into the catalytic mechanism and active-site environment of Comamonas testosteroni delta 5-3-ketosteroid isomerase as revealed by site-directed mutagenesis of the catalytic base aspartate-38". Biochemistry. 34 (43): 14245–53. doi:10.1021/bi00043a032. PMID 7578024.
- Lamba V, Yabukarski F, Pinney M, Herschlag D (August 2016). "Evaluation of the Catalytic Contribution from a Positioned General Base in Ketosteroid Isomerase". Journal of the American Chemical Society. 138 (31): 9902–9. doi:10.1021/jacs.6b04796. PMID 27410422.
- Sigala PA, Fafarman AT, Bogard PE, Boxer SG, Herschlag D (October 2007). "Do ligand binding and solvent exclusion alter the electrostatic character within the oxyanion hole of an enzymatic active site?". Journal of the American Chemical Society. 129 (40): 12104–5. doi:10.1021/ja075605a. PMC 3171184. PMID 17854190.
- Zhao Q, Li YK, Mildvan AS, Talalay P (May 1995). "Ultraviolet spectroscopic evidence for decreased motion of the active site tyrosine residue of delta 5-3-ketosteroid isomerase by steroid binding". Biochemistry. 34 (19): 6562–72. doi:10.1021/bi00019a038. PMID 7756287.
- Zhao Q, Abeygunawardana C, Mildvan AS (February 1996). "13C NMR relaxation studies of backbone and side chain motion of the catalytic tyrosine residue in free and steroid-bound delta 5-3-ketosteroid isomerase". Biochemistry. 35 (5): 1525–32. doi:10.1021/bi9525381. PMID 8634283.
- Schwans JP, Kraut DA, Herschlag D (August 2009). "Determining the catalytic role of remote substrate binding interactions in ketosteroid isomerase". Proceedings of the National Academy of Sciences of the United States of America. 106 (34): 14271–5. Bibcode:2009PNAS..10614271S. doi:10.1073/pnas.0901032106. PMC 2732871. PMID 19706511.
- Kim SW, Cha SS, Cho HS, Kim JS, Ha NC, Cho MJ, Joo S, Kim KK, Choi KY, Oh BH (November 1997). "High-resolution crystal structures of delta5-3-ketosteroid isomerase with and without a reaction intermediate analogue". Biochemistry. 36 (46): 14030–6. doi:10.1021/bi971546+. PMID 9369474.
- Kawahara FS, Wang SF, Talalay P (May 1962). "The preparation and properties of crystalline delta5-3-ketosteroid isomerase". The Journal of Biological Chemistry. 237 (5): 1500–6. doi:10.1016/S0021-9258(19)83730-4. PMID 14454546.
- Talalay P, Dobson MM, Tapley DF (October 1952). "Oxidative degradation of testosterone by adaptive enzymes". Nature. 170 (4328): 620–1. Bibcode:1952Natur.170..620T. doi:10.1038/170620a0. PMID 13002385. S2CID 4181660.
- Lachance Y, Luu-The V, Labrie C, Simard J, Dumont M, de Launoit Y, Guérin S, Leblanc G, Labrie F (February 1992). "Characterization of human 3 beta-hydroxysteroid dehydrogenase/delta 5-delta 4-isomerase gene and its expression in mammalian cells". The Journal of Biological Chemistry. 267 (5): 3551. doi:10.1016/S0021-9258(19)50764-5. PMID 1737804.
- Horinouchi M, Hayashi T, Kudo T (March 2012). "Steroid degradation in Comamonas testosteroni". The Journal of Steroid Biochemistry and Molecular Biology. 129 (1–2): 4–14. doi:10.1016/j.jsbmb.2010.10.008. hdl:10069/24613. PMID 21056662. S2CID 140206626.
- Horinouchi M, Kurita T, Hayashi T, Kudo T (October 2010). "Steroid degradation genes in Comamonas testosteroni TA441: Isolation of genes encoding a Δ4(5)-isomerase and 3α- and 3β-dehydrogenases and evidence for a 100 kb steroid degradation gene hot spot". The Journal of Steroid Biochemistry and Molecular Biology. 122 (4): 253–63. doi:10.1016/j.jsbmb.2010.06.002. PMID 20554032. S2CID 206497547.
- Gerlt JA, Gassman PG (November 1993). "Understanding the rates of certain enzyme-catalyzed reactions: proton abstraction from carbon acids, acyl-transfer reactions, and displacement reactions of phosphodiesters". Biochemistry. 32 (45): 11943–52. doi:10.1021/bi00096a001. PMID 8218268.
- Gerlt JA, Gassman PG (December 1993). "An explanation for rapid enzyme-catalyzed proton abstraction from carbon acids: importance of late transition states in concerted mechanisms". Biochemistry. 115 (24): 11552–11568. doi:10.1021/ja00077a062.
- Warshel A, Papazyan A, Kollman PA (July 1995). "On low-barrier hydrogen bonds and enzyme catalysis". Science. 269 (5220): 102–6. Bibcode:1995Sci...269..102W. doi:10.1126/science.7661987. PMID 7661987.
- Guthrie JP (March 1996). "Short strong hydrogen bonds: can they explain enzymic catalysis?". Chemistry & Biology. 3 (3): 163–70. doi:10.1016/s1074-5521(96)90258-6. PMID 8807842.
Further reading
- Ewald W, Werbin H, Chaikoff IL (November 1965). "Evidence for the presence of 17-hydroxypregnenedione isomerase in beef adrenal cortex". Biochimica et Biophysica Acta (BBA) - General Subjects. 111 (1): 306–12. doi:10.1016/0304-4165(65)90497-6. PMID 5867327.
- Kawahara FS, Talalay P (January 1960). "Crystalline Delta 5-3-ketosteroid isomerase". The Journal of Biological Chemistry. 235: PC1–2. doi:10.1016/S0021-9258(18)69620-6. PMID 14404954.
- Talalay P, Wang VS (October 1955). "Enzymic isomerization of delta5-3-ketosteroids". Biochimica et Biophysica Acta. 18 (2): 300–1. doi:10.1016/0006-3002(55)90079-2. PMID 13276386.
- Steroid+Isomerases at the U.S. National Library of Medicine Medical Subject Headings (MeSH)