Tetanospasmin
Tetanus toxin (TeNT) is an extremely potent neurotoxin produced by the vegetative cell of Clostridium tetani[1] in anaerobic conditions, causing tetanus. It has no known function for clostridia in the soil environment where they are normally encountered. It is also called spasmogenic toxin, tentoxilysin, tetanospasmin or, tetanus neurotoxin. The LD50 of this toxin has been measured to be approximately 2.5–3 ng/kg,[2][3] making it second only to the related botulinum toxin (LD50 2 ng/kg)[4] as the deadliest toxin in the world. However, these tests are conducted solely on mice, which may react to the toxin differently from humans and other animals.
| Tetanospasmin | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Identifiers | |||||||
| Organism | |||||||
| Symbol | TeNT | ||||||
| UniProt | P04958 | ||||||
| |||||||
| Tentoxilysin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC no. | 3.4.24.68 | ||||||||
| CAS no. | 107231-12-9 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
C. tetani also produces the exotoxin tetanolysin, a hemolysin, that causes destruction of tissues.[5]
Distribution
Tetanus toxin spreads through tissue spaces into the lymphatic and vascular systems. It enters the nervous system at the neuromuscular junctions and migrates through nerve trunks and into the central nervous system (CNS) by retrograde axonal transport by using dyneins.[6][7]
Structure
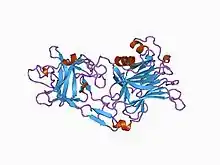
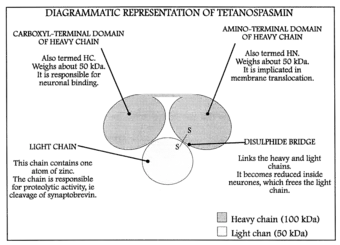
The tetanus toxin protein has a molecular weight of 150 kDa. It is translated from the tetX gene as one protein which is subsequently cleaved into two parts: a 100 kDa heavy or B-chain and a 50 kDa light or A-chain. The chains are connected by a disulfide bond.
- The B-chain binds to disialogangliosides (GD2 and GD1b) on the neuronal membrane and contains a translocation domain which aids the movement of the protein across that membrane and into the neuron.
- The A-chain, an M27-family zinc endopeptidase, attacks the vesicle-associated membrane protein (VAMP).
The TetX gene encoding this protein is located on the PE88 plasmid.[8][9]
Several structures of the binding domain and the peptidase domain have been solved by X-ray crystallography and deposited in the PDB. A summary of these structures is available using the UniPDB Archived 2013-06-13 at the Wayback Machine application at PDBe, for example 1z7h or 3hmy.
Mechanism of action
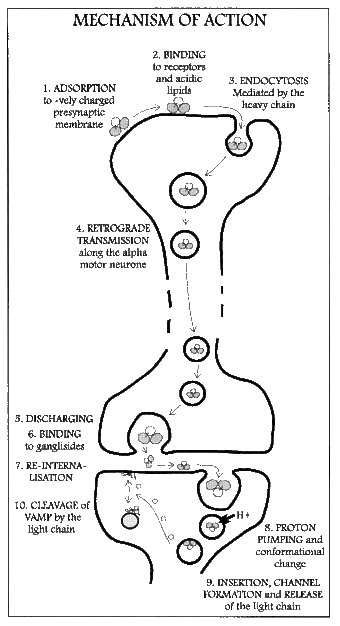
The mechanism of TeNT action can be broken down and discussed in these different steps:
- Transport
- Specific binding in the periphery neurons
- Retrograde axonal transport to the CNS inhibitory interneurons
- Transcytosis from the axon into the inhibitory interneurons
- Action
The first three steps outline the travel of tetanus toxin from the peripheral nervous system to where it is taken up to the CNS and has its final effect. The last three steps document the changes necessary for the final mechanism of the neurotoxin.
Transport to the CNS inhibitory interneurons begins with the B-chain mediating the neurospecific binding of TeNT to the nerve terminal membrane. It binds to GT1b polysialogangliosides, similarly to the C. botulinum neurotoxin. It also binds to another poorly characterized GPI-anchored protein receptor more specific to TeNT.[10][11] Both the ganglioside and the GPI-anchored protein are located in lipid microdomains and both are requisite for specific TeNT binding.[11] Once it is bound, the neurotoxin is then endocytosed into the nerve and begins to travel through the axon to the spinal neurons. The next step, transcytosis from the axon into the CNS inhibitory interneuron, is one of the least understood parts of TeNT action. At least two pathways are involved, one that relies on the recycling of synaptic vesicle 2 (SV2) system and one that does not.[12]
Once the vesicle is in the inhibitory interneuron, its translocation is mediated by pH and temperature, specifically a low or acidic pH in the vesicle and standard physiological temperatures.[13][14] Once the toxin has been translocated into the cytosol, chemical reduction of the disulfide bond to separate thiols occurs, mainly by the enzyme NADPH-thioredoxin reductase-thioredoxin. The light chain is then free to cleave the Gln76-Phe77 bond of synaptobrevin.[15] Cleavage of synaptobrevin affects the stability of the SNARE core by restricting it from entering the low-energy conformation, which is the target for NSF binding.[16] Synaptobrevin is an integral V-SNARE necessary for vesicle fusion to membranes. The final target of TeNT is the cleavage of synaptobrevin and, even in low doses, has the effect of interfering with exocytosis of neurotransmitters from inhibitory interneurons. The blockage of the neurotransmitters γ-aminobutyric acid (GABA) and glycine is the direct cause of the physiological effects that TeNT induces. GABA inhibits motor neurons, so by blocking GABA, tetanus toxin causes violent spastic paralysis.[17] The action of the A-chain also stops the affected neurons from releasing excitatory transmitters,[18] by degrading the protein synaptobrevin 2.[19] The combined consequence is dangerous overactivity in the muscles from the smallest sensory stimuli, as the damping of motor reflexes is inhibited, leading to generalized contractions of the agonist and antagonist musculature, termed a "tetanic spasm".
Clinical significance
The clinical manifestations of tetanus are caused when tetanus toxin blocks inhibitory impulses, by interfering with the release of neurotransmitters, including glycine and gamma-aminobutyric acid. These inhibitory neurotransmitters inhibit the alpha motor neurons. With diminished inhibition, the resting firing rate of the alpha motor neuron increases, producing rigidity, unopposed muscle contraction and spasm. Characteristic features are risus sardonicus (a rigid smile), trismus (commonly known as "lock-jaw"), and opisthotonus (rigid, arched back). Seizures may occur, and the autonomic nervous system may also be affected. Tetanospasmin appears to prevent the release of neurotransmitters by selectively cleaving a component of synaptic vesicles called synaptobrevin II.[20] Loss of inhibition also affects preganglionic sympathetic neurons in the lateral gray matter of the spinal cord and produces sympathetic hyperactivity and high circulating catecholamine levels. Hypertension and tachycardia alternating with hypotension and bradycardia may develop.[21][22]
Tetanic spasms can occur in a distinctive form called opisthotonos and be sufficiently severe to fracture long bones. The shorter nerves are the first to be inhibited, which leads to the characteristic early symptoms in the face and jaw, risus sardonicus and lockjaw.
Immunity and vaccination
Due to its extreme potency, even a lethal dose of tetanospasmin may be insufficient to provoke an immune response. Naturally acquired tetanus infections thus do not usually provide immunity to subsequent infections. Immunization (which is impermanent and must be repeated periodically) instead uses the less deadly toxoid derived from the toxin, as in the tetanus vaccine and some combination vaccines (such as DTP).
Evolution of the Toxin
Very little has been written or researched on how and why tentanospasmin evolved in animals. The toxin is highly specific to muscular neurons in higher animals and is carried by a plasmid which implies that the host bacterium acquired the plasmid through horizontal gene transfer from another source organism as is the case of most plasmid based toxins such as shigatoxin. Although tentanospasmin is now classified as an extremely potent neurotoxin, it's evolutionary origins may indicate it may have served an entirely different function in early animal life. It is routinely carried by anaerobic organisms and early life on earth was typically capable of both aerobic and anaerobic respiration. By way of example, human DNA still contains all the genes necessary for anaerobic respiration and one example is the lens cells of the human eye, which still operate in anaerobic mode. Most lower animals, and even some mammals are capable of anaerobic respiration during their hibernation phases. The spasms typically caused by this toxin mimic cardiac rhythms in skeletal muscles in most reptiles exposed to the toxin and would provide an evolutionary advantage to a hibernating reptile by keeping blood moving slowly around the organism while in hibernation mode if the organism switched to anaerobic respiration. This theory is supported by the fact the tetanus bacteria uses a "pump and dump" strategy by secreting the toxin inside a plasmid membrane then exocysing the plasmid vacuole into target tissue. The toxin in not immediately released typically. This would imply that tetani evolved as a symbiote in early primitive reptiles to provide a means of pumping blood around during hibernation cycles since a molecule this complex and highly specialized would have to confer a survival advantage to exist at all. There is no evolutionary pressure outside this model which can even remotely explain how and why a substance this potent and highly specialized would exist. Higher mammals host this bacteria as a common component of intestinal gut flora and mouth bacteria without any ill effects and it only becomes a problem in anaerobic conditions. It has absolutely no function whatsoever in the bacterium's host environment which is soil and manure and is harmless to any competing organisms. It only affects higher animals which have neuro muscular junctions which makes it's very existence a mystery to modern science.
References
- "tetanospasmin" at Dorland's Medical Dictionary
- "Pinkbook | Tetanus | Epidemiology of Vaccine Preventable Diseases | CDC". www.cdc.gov. Retrieved 2017-01-18.
- "Toxin Table » Environmental Health & Safety » University of Florida". www.ehs.ufl.edu. Archived from the original on 2017-01-18. Retrieved 2017-01-18.
- "Botulism". World Health Organization. Retrieved 2017-01-18.
- Willey, Joanne (2009). Prescott's Principles of Microbiology. New York City, NY: McGraw-Hill. pp. 481. ISBN 978-0-07-337523-6.
- Farrar JJ; Yen LM; Cook T; Fairweather N; Binh N; Parry J; Parry CM (September 2000). "Tetanus". Journal of Neurology, Neurosurgery, and Psychiatry. 69 (3): 292–301. doi:10.1136/jnnp.69.3.292. PMC 1737078. PMID 10945801.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - AU Lalli G, Gschmeissner S, Schiavo G (November 15, 2003). "Myosin Va and microtubule-based motors are required for fast axonal retrograde transport of tetanus toxin in motor neurons". Journal of Cell Science. 116 (22): 4639–50. doi:10.1242/jcs.00727. PMID 14576357.
- Eisel U, Jarausch W, Goretzki K, Henschen A, Engels J, Weller U, Hudel M, Habermann E, Niemann H (1986). "Tetanus toxin: primary structure, expression in E. coli, and homology with botulinum toxins". EMBO J. 5 (10): 2495–502. doi:10.1002/j.1460-2075.1986.tb04527.x. PMC 1167145. PMID 3536478.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Popp D, Narita A, Lee LJ, Ghoshdastider U, Xue B, Srinivasan R, Balasubramanian MK, Tanaka T, Robinson RC (2012). "Novel actin-like filament structure from Clostridium tetani". The Journal of Biological Chemistry. 287 (25): 21121–9. doi:10.1074/jbc.M112.341016. PMC 3375535. PMID 22514279.
- Munro, P; Kojima, H; Dupont, JL; Bossu, JL; Poulain, B; Boquet, P (30 November 2001). "High sensitivity of mouse neuronal cells to tetanus toxin requires a GPI-anchored protein". Biochemical and Biophysical Research Communications. 289 (2): 623–9. doi:10.1006/bbrc.2001.6031. PMID 11716521.
- Winter, A; Ulrich, WP; Wetterich, F; Weller, U; Galla, HJ (17 June 1996). "Gangliosides in phospholipid bilayer membranes: interaction with tetanus toxin". Chemistry and Physics of Lipids. 81 (1): 21–34. doi:10.1016/0009-3084(96)02529-7. PMID 9450318.
- Yeh, FL; Dong, M; Yao, J; Tepp, WH; Lin, G; Johnson, EA; Chapman, ER (24 November 2010). "SV2 mediates entry of tetanus neurotoxin into central neurons" (PDF). PLOS Pathogens. 6 (11): e1001207. doi:10.1371/journal.ppat.1001207. PMC 2991259. PMID 21124874.
- Pirazzini, M; Rossetto, O; Bertasio, C; Bordin, F; Shone, CC; Binz, T; Montecucco, C (4 January 2013). "Time course and temperature dependence of the membrane translocation of tetanus and botulinum neurotoxins C and D in neurons". Biochemical and Biophysical Research Communications. 430 (1): 38–42. doi:10.1016/j.bbrc.2012.11.048. PMID 23200837.
- Burns, JR; Baldwin, MR (8 August 2014). "Tetanus neurotoxin utilizes two sequential membrane interactions for channel formation". The Journal of Biological Chemistry. 289 (32): 22450–8. doi:10.1074/jbc.m114.559302. PMC 4139251. PMID 24973217.
- Pirazzini, M; Bordin, F; Rossetto, O; Shone, CC; Binz, T; Montecucco, C (16 January 2013). "The thioredoxin reductase-thioredoxin system is involved in the entry of tetanus and botulinum neurotoxins in the cytosol of nerve terminals". FEBS Letters. 587 (2): 150–5. doi:10.1016/j.febslet.2012.11.007. PMID 23178719.
- Pellegrini, LL; O'Connor, V; Lottspeich, F; Betz, H (2 October 1995). "Clostridial neurotoxins compromise the stability of a low-energy SNARE complex mediating NSF activation of synaptic vesicle fusion". The EMBO Journal. 14 (19): 4705–13. doi:10.1002/j.1460-2075.1995.tb00152.x. PMC 394567. PMID 7588600.
- Kumar, Vinay; Abbas, Abul K.; Fausto, Nelson; Aster, Jon (2009-05-28). Robbins and Cotran Pathologic Basis of Disease, Professional Edition: Expert Consult - Online (Robbins Pathology) (Kindle Locations 19359-19360). Elsevier Health. Kindle Edition.
- Kanda K, Takano K (February 1983). "Effect of tetanus toxin on the excitatory and the inhibitory post-synaptic potentials in the cat motoneurone". J. Physiol. 335: 319–333. doi:10.1113/jphysiol.1983.sp014536. PMC 1197355. PMID 6308220.
- Schiavo G, Benfenati F, Poulain B, Rossetto O, Polverino de Laureto P, DasGupta BR, Montecucco C (October 29, 1992). "Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin". Nature. 359 (6398): 832–5. Bibcode:1992Natur.359..832S. doi:10.1038/359832a0. PMID 1331807. S2CID 4241066.
- Todar, Ken (2005). "Pathogenic Clostridia, including Botulism and Tetanus". Todar's Online Textbook of Bacteriology. Retrieved 24 June 2018.
- Loscalzo, Joseph; Fauci, Anthony S.; Braunwald, Eugene; Dennis L. Kasper; Hauser, Stephen L; Longo, Dan L. (2008). Harrison's principles of internal medicine. McGraw-Hill Medical. ISBN 978-0-07-146633-2.
- "Tetanus in Emergency Medicine". Emedicine. Retrieved 2011-09-01.
External links
- tetanospasmin at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
 Media related to Tetanus neurotoxin at Wikimedia Commons
Media related to Tetanus neurotoxin at Wikimedia Commons- Tentoxilysin at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Pellizzari R, Rossetto O, Schiavo G, Montecucco C (1999). "Tetanus and botulinum neurotoxins: mechanism of action and therapeutic uses". Philos. Trans. R. Soc. Lond. B Biol. Sci. 354 (1381): 259–68. doi:10.1098/rstb.1999.0377. PMC 1692495. PMID 10212474.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Neurotoxin in Toxicon
- The journey of tetanus and botulinum neurotoxins in neurons
- How do tetanus and botulinum toxins bind to neuronal membranes?