Transcription activator-like effector
TAL (transcription activator-like) effectors (often referred to as TALEs, but not to be confused with the three amino acid loop extension homeobox class of proteins) are proteins secreted by some β- and γ-proteobacteria.[1] Most of these are Xanthomonads. Plant pathogenic Xanthomonas bacteria are especially known for TALEs, produced via their type III secretion system. These proteins can bind promoter sequences in the host plant and activate the expression of plant genes that aid bacterial infection. The TALE domain responsible for binding to DNA is known to have 1.5 to 33.5 short sequences that are repeated multiple times (tandem repeats).[2] Each of these repeats was found to be specific for a certain base pair of the DNA.[2] These repeats also have repeat variable residues (RVD) that can detect specific DNA base pairs.[2] They recognize plant DNA sequences through a central repeat domain consisting of a variable number of ~34 amino acid repeats. There appears to be a one-to-one correspondence between the identity of two critical amino acids in each repeat and each DNA base in the target sequence. These proteins are interesting to researchers both for their role in disease of important crop species and the relative ease of retargeting them to bind new DNA sequences. Similar proteins can be found in the pathogenic bacterium Ralstonia solanacearum[3][4][1] and Burkholderia rhizoxinica,[5][1] as well as yet unidentified marine microorganisms.[6] The term TALE-likes is used to refer to the putative protein family encompassing the TALEs and these related proteins.
 | |||||||
| Identifiers | |||||||
|---|---|---|---|---|---|---|---|
| Organism | |||||||
| Symbol | pthXo1 | ||||||
| RefSeq (Prot) | WP_041182630.1 | ||||||
| UniProt | B2SU53 | ||||||
| Other data | |||||||
| Chromosome | Genomic: 1.65 - 1.65 Mb | ||||||
| |||||||
Function in plant pathogenesis
Xanthomonas
Xanthomonas are Gram-negative bacteria that can infect a wide variety of plant species including pepper/capsicum, rice, citrus, cotton, tomato, and soybeans.[7] Some types of Xanthomonas cause localized leaf spot or leaf streak while others spread systemically and cause black rot or leaf blight disease. They inject a number of effector proteins, including TAL effectors, into the plant via their type III secretion system. TAL effectors have several motifs normally associated with eukaryotes including multiple nuclear localization signals and an acidic activation domain. When injected into plants, these proteins can enter the nucleus of the plant cell, bind plant promoter sequences, and activate transcription of plant genes that aid in bacterial infection.[7] Plants have developed a defense mechanism against type III effectors that includes R (resistance) genes triggered by these effectors. Some of these R genes appear to have evolved to contain TAL-effector binding sites similar to site in the intended target gene. This competition between pathogenic bacteria and the host plant has been hypothesized to account for the apparently malleable nature of the TAL effector DNA binding domain.[8]
| X. species | victim crop |
|---|---|
| X. campestris pv. musacerum, X. vasicola pv. musacerum | banana and ensete |
| X. axonopodis pv. phaseoli, X. axonopodis pv. glycines | common bean and soybean |
| X. axonopodis pv. vignicola | cowpea |
| X. campestris pv. campestris, X. campestris pv. armoracieae | all Brassicas |
| X. axonopodis pv. manihotis | cassava |
| X. translucens pv. translucens | barley, rye, wheat, triticale |
| X. axonopodis pv. citri, X. citri, X. citri pv. aurantifolii | citrus |
| X. campestris pv. malvacearum | cotton |
| X. euvesicatoria, X. gardneri | pepper/capsicum |
| X. oryzae pv. oryzae | rice |
| X. arboricola pv. pruni | plum, peach, nectarine |
| X. perforans, X. campestris pv. vesicatoria | tomato |
| X. campestris pv. mangiferaindicae | mango |
| X. arboricola pv. juglandis | walnut |
Non-Xanthomonas
R. solanacearum, B. rhizoxinica, and banana blood disease (a bacterium not yet definitively identified, in the R. solanacearum species group).[1]
DNA recognition
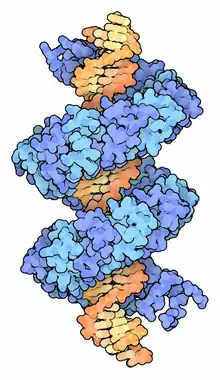
The most distinctive characteristic of TAL effectors is a central repeat domain containing between 1.5 and 33.5 repeats that are usually 34 residues in length (the C-terminal repeat is generally shorter and referred to as a “half repeat”).[7] A typical repeat sequence is LTPEQVVAIASHDGGKQALETVQRLLPVLCQAHG, but the residues at the 12th and 13th positions are hypervariable (these two amino acids are also known as the repeat variable diresidue or RVD). There is a simple relationship between the identity of these two residues in sequential repeats and sequential DNA bases in the TAL effector's target site.[8] The crystal structure of a TAL effector bound to DNA indicates that each repeat comprises two alpha helices and a short RVD-containing loop where the second residue of the RVD makes sequence-specific DNA contacts while the first residue of the RVD stabilizes the RVD-containing loop.[10][11] Target sites of TAL effectors also tend to include a thymine flanking the 5’ base targeted by the first repeat; this appears to be due to a contact between this T and a conserved tryptophan in the region N-terminal of the central repeat domain.[10] However, this "zero" position does not always contain a thymine, as some scaffolds are more permissive.[12]
The TAL-DNA code was broken by two separate groups in 2010.[8] The first group, headed by Adam Bogdanove, broke this code computationally by searching for patterns in protein sequence alignments and DNA sequences of target promoters derived from a database of genes upregulated by TALEs.[13] The second group (Boch) deduced the code through molecular analysis of the TAL effector AvrBs3 and its target DNA sequence in the promoter of a pepper gene activated by AvrBs3.[14] The experimentally validated code between RVD sequence and target DNA base can be expressed as follows:
| Residue | Base | Notes | References |
|---|---|---|---|
| NI | A | [14] | |
| HD | C | Not 5-methyl-C | [14] |
| NG | T, 5mC | [14][15] | |
| NN | R | Purine: G or A | [14] |
| NS | N | Any | [14] |
| NK | G | Reduced TALEN activity if used exclusively | [16][17] |
| NH | G | [9] |
Target genes
TAL effectors can induce susceptibility genes that are members of the NODULIN3 (N3) gene family. These genes are essential for the development of the disease. In rice two genes, Os-8N3 and Os-11N3, are induced by TAL effectors. Os-8N3 is induced by PthXo1 and Os-11N3 is induced by PthXo3 and AvrXa7. Two hypotheses exist about possible functions for N3 proteins:
- They are involved in copper transport, resulting in detoxification of the environment for bacteria. The reduction in copper level facilitates bacterial growth.
- They are involved in glucose transport, facilitating glucose flow. This mechanism provides nutrients to bacteria and stimulates pathogen growth and virulence
Engineering TAL effectors
This simple correspondence between amino acids in TAL effectors and DNA bases in their target sites makes them useful for protein engineering applications. Numerous groups have designed artificial TAL effectors capable of recognizing new DNA sequences in a variety of experimental systems.[14][16][17][18][19][20] Such engineered TAL effectors have been used to create artificial transcription factors that can be used to target and activate or repress endogenous genes in tomato,[16] Arabidopsis thaliana,[16] and human cells.[17][19][9][21]
Genetic constructs to encode TAL effector-based proteins can be made using either conventional gene synthesis or modular assembly.[19][21][22][23][24][25][26][27] A plasmid kit for assembling custom TALEN and other TAL effector constructs is available through the public, not-for-profit repository Addgene. Webpages providing access to public software, protocols, and other resources for TAL effector-DNA targeting applications include the TAL Effector-Nucleotide Targeter and taleffectors.com.
Applications
Engineered TAL effectors can also be fused to the cleavage domain of FokI to create TAL effector nucleases (TALEN) or to meganucleases (nucleases with longer recognition sites) to create "megaTALs."[28] Such fusions share some properties with zinc finger nucleases and may be useful for genetic engineering and gene therapy applications.[29]
TALEN-based approaches are used in the emerging fields of gene editing and genome engineering. TALEN fusions show activity in a yeast-based assay,[18][30] at endogenous yeast genes,[22] in a plant reporter assay,[20] at an endogenous plant gene,[23] at endogenous zebrafish genes,[31][32] at an endogenous rat gene,[33] and at endogenous human genes.[17][23][34] The human HPRT1 gene has been targeted at detectable, but unquantified levels.[23] In addition, TALEN constructs containing the FokI cleavage domain fused to a smaller portion of the TAL effector still containing the DNA binding domain have been used to target the endogenous NTF3 and CCR5 genes in human cells with efficiencies of up to 25%.[17] TAL effector nucleases have also been used to engineer human embryonic stem cells and induced pluripotent stem cells (IPSCs)[34] and to knock out the endogenous ben-1 gene in C. elegans.[35]
TALE-induced non-homologous end joining modification has been used to produce novel disease resistance in rice.[1]
See also
References
- Schornack S, Moscou MJ, Ward ER, Horvath DM (2013-08-04). "Engineering plant disease resistance based on TAL effectors". Annual Review of Phytopathology. Annual Reviews. 51 (1): 383–406. doi:10.1146/annurev-phyto-082712-102255. PMID 23725472.
- Deng, Dong; Yan, Chuangye; Pan, Xiaojing; Mahfouz, Magdy; Wang, Jiawei; Zhu, Jian-Kang; Shi, Yigong; Yan, Nieng (2012-02-10). "Structural basis for sequence-specific recognition of DNA by TAL effectors". Science. 335 (6069): 720–723. Bibcode:2012Sci...335..720D. doi:10.1126/science.1215670. ISSN 1095-9203. PMC 3586824. PMID 22223738.
- Heuer H, Yin YN, Xue QY, Smalla K, Guo JH (July 2007). "Repeat domain diversity of avrBs3-like genes in Ralstonia solanacearum strains and association with host preferences in the field". Applied and Environmental Microbiology. 73 (13): 4379–84. Bibcode:2007ApEnM..73.4379H. doi:10.1128/AEM.00367-07. PMC 1932761. PMID 17468277.
- Li L, Atef A, Piatek A, Ali Z, Piatek M, Aouida M, et al. (July 2013). "Characterization and DNA-binding specificities of Ralstonia TAL-like effectors". Molecular Plant. 6 (4): 1318–30. doi:10.1093/mp/sst006. PMC 3716395. PMID 23300258.
- de Lange O, Wolf C, Dietze J, Elsaesser J, Morbitzer R, Lahaye T (June 2014). "Programmable DNA-binding proteins from Burkholderia provide a fresh perspective on the TALE-like repeat domain". Nucleic Acids Research. 42 (11): 7436–49. doi:10.1093/nar/gku329. PMC 4066763. PMID 24792163.
- de Lange O, Wolf C, Thiel P, Krüger J, Kleusch C, Kohlbacher O, Lahaye T (November 2015). "DNA-binding proteins from marine bacteria expand the known sequence diversity of TALE-like repeats". Nucleic Acids Research. 43 (20): 10065–80. doi:10.1093/nar/gkv1053. PMC 4787788. PMID 26481363.
- Boch J, Bonas U (September 2010). "Xanthomonas AvrBs3 family-type III effectors: discovery and function". Annual Review of Phytopathology. 48: 419–36. doi:10.1146/annurev-phyto-080508-081936. PMID 19400638.
- Voytas DF, Joung JK (December 2009). "Plant science. DNA binding made easy". Science. 326 (5959): 1491–2. Bibcode:2009Sci...326.1491V. doi:10.1126/science.1183604. PMC 7814878. PMID 20007890. S2CID 33257689.
- Cong L, Zhou R, Kuo YC, Cunniff M, Zhang F (July 2012). "Comprehensive interrogation of natural TALE DNA-binding modules and transcriptional repressor domains". Nature Communications. 968. 3 (7): 968. Bibcode:2012NatCo...3..968C. doi:10.1038/ncomms1962. PMC 3556390. PMID 22828628.
- Mak AN, Bradley P, Cernadas RA, Bogdanove AJ, Stoddard BL (February 2012). "The crystal structure of TAL effector PthXo1 bound to its DNA target". Science. 335 (6069): 716–9. Bibcode:2012Sci...335..716M. doi:10.1126/science.1216211. PMC 3427646. PMID 22223736.
- Deng D, Yan C, Pan X, Mahfouz M, Wang J, Zhu JK, et al. (February 2012). "Structural basis for sequence-specific recognition of DNA by TAL effectors". Science. 335 (6069): 720–3. Bibcode:2012Sci...335..720D. doi:10.1126/science.1215670. PMC 3586824. PMID 22223738.
- Stella S, Molina R, Yefimenko I, Prieto J, Silva G, Bertonati C, et al. (September 2013). "Structure of the AvrBs3-DNA complex provides new insights into the initial thymine-recognition mechanism". Acta Crystallographica. Section D, Biological Crystallography. 69 (Pt 9): 1707–16. doi:10.1107/S0907444913016429. PMC 3760130. PMID 23999294.
- Moscou MJ, Bogdanove AJ (December 2009). "A simple cipher governs DNA recognition by TAL effectors". Science. 326 (5959): 1501. Bibcode:2009Sci...326.1501M. doi:10.1126/science.1178817. PMID 19933106. S2CID 6648530.
- Boch J, Scholze H, Schornack S, Landgraf A, Hahn S, Kay S, et al. (December 2009). "Breaking the code of DNA binding specificity of TAL-type III effectors". Science. 326 (5959): 1509–12. Bibcode:2009Sci...326.1509B. doi:10.1126/science.1178811. PMID 19933107. S2CID 206522347.
- Deng D, Yin P, Yan C, Pan X, Gong X, Qi S, et al. (October 2012). "Recognition of methylated DNA by TAL effectors". Cell Research. 22 (10): 1502–4. doi:10.1038/cr.2012.127. PMC 3463267. PMID 22945353.
- Morbitzer R, Römer P, Boch J, Lahaye T (December 2010). "Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors". Proceedings of the National Academy of Sciences of the United States of America. 107 (50): 21617–22. Bibcode:2010PNAS..10721617M. doi:10.1073/pnas.1013133107. PMC 3003021. PMID 21106758.
- Miller JC, Tan S, Qiao G, Barlow KA, Wang J, Xia DF, et al. (February 2011). "A TALE nuclease architecture for efficient genome editing". Nature Biotechnology. 29 (2): 143–8. doi:10.1038/nbt.1755. PMID 21179091. S2CID 53549397.
- Christian M, Cermak T, Doyle EL, Schmidt C, Zhang F, Hummel A, et al. (October 2010). "Targeting DNA double-strand breaks with TAL effector nucleases". Genetics. 186 (2): 757–61. doi:10.1534/genetics.110.120717. PMC 2942870. PMID 20660643.
- Zhang F, Cong L, Lodato S, Kosuri S, Church GM, Arlotta P (February 2011). "Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription". Nature Biotechnology. 29 (2): 149–53. doi:10.1038/nbt.1775. PMC 3084533. PMID 21248753.
- Mahfouz MM, Li L, Shamimuzzaman M, Wibowo A, Fang X, Zhu JK (February 2011). "De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks". Proceedings of the National Academy of Sciences of the United States of America. 108 (6): 2623–8. Bibcode:2011PNAS..108.2623M. doi:10.1073/pnas.1019533108. PMC 3038751. PMID 21262818.
- Geissler R, Scholze H, Hahn S, Streubel J, Bonas U, Behrens SE, Boch J (2011). Shiu S (ed.). "Transcriptional activators of human genes with programmable DNA-specificity". PLOS ONE. 6 (5): e19509. Bibcode:2011PLoSO...619509G. doi:10.1371/journal.pone.0019509. PMC 3098229. PMID 21625585.
- Li T, Huang S, Zhao X, Wright DA, Carpenter S, Spalding MH, et al. (August 2011). "Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes". Nucleic Acids Research. 39 (14): 6315–25. doi:10.1093/nar/gkr188. PMC 3152341. PMID 21459844.
- Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, et al. (July 2011). "Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting". Nucleic Acids Research. 39 (12): e82. doi:10.1093/nar/gkr218. PMC 3130291. PMID 21493687.
- Morbitzer R, Elsaesser J, Hausner J, Lahaye T (July 2011). "Assembly of custom TALE-type DNA binding domains by modular cloning". Nucleic Acids Research. 39 (13): 5790–9. doi:10.1093/nar/gkr151. PMC 3141260. PMID 21421566.
- Weber E, Gruetzner R, Werner S, Engler C, Marillonnet S (2011). Bendahmane M (ed.). "Assembly of designer TAL effectors by Golden Gate cloning". PLOS ONE. 6 (5): e19722. Bibcode:2011PLoSO...619722W. doi:10.1371/journal.pone.0019722. PMC 3098256. PMID 21625552.
- Sanjana NE, Cong L, Zhou Y, Cunniff MM, Feng G, Zhang F (January 2012). "A transcription activator-like effector toolbox for genome engineering". Nature Protocols. 7 (1): 171–92. doi:10.1038/nprot.2011.431. PMC 3684555. PMID 22222791.
- Briggs AW, Rios X, Chari R, Yang L, Zhang F, Mali P, Church GM (August 2012). "Iterative capped assembly: rapid and scalable synthesis of repeat-module DNA such as TAL effectors from individual monomers". Nucleic Acids Research. 40 (15): e117. doi:10.1093/nar/gks624. PMC 3424587. PMID 22740649.
- Boissel S, Jarjour J, Astrakhan A, Adey A, Gouble A, Duchateau P, et al. (February 2014). "megaTALs: a rare-cleaving nuclease architecture for therapeutic genome engineering". Nucleic Acids Research. 42 (4): 2591–601. doi:10.1093/nar/gkt1224. PMC 3936731. PMID 24285304.
- DeFrancesco L (August 2011). "Move over ZFNs". Nature Biotechnology. 29 (8): 681–4. doi:10.1038/nbt.1935. PMID 21822235. S2CID 29925336.
- Li T, Huang S, Jiang WZ, Wright D, Spalding MH, Weeks DP, Yang B (January 2011). "TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain". Nucleic Acids Research. 39 (1): 359–72. doi:10.1093/nar/gkq704. PMC 3017587. PMID 20699274.
- Huang P, Xiao A, Zhou M, Zhu Z, Lin S, Zhang B (August 2011). "Heritable gene targeting in zebrafish using customized TALENs". Nature Biotechnology. 29 (8): 699–700. doi:10.1038/nbt.1939. PMID 21822242. S2CID 28802632.
- Sander JD, Cade L, Khayter C, Reyon D, Peterson RT, Joung JK, Yeh JR (August 2011). "Targeted gene disruption in somatic zebrafish cells using engineered TALENs". Nature Biotechnology. 29 (8): 697–8. doi:10.1038/nbt.1934. PMC 3154023. PMID 21822241.
- Tesson L, Usal C, Ménoret S, Leung E, Niles BJ, Remy S, et al. (August 2011). "Knockout rats generated by embryo microinjection of TALENs". Nature Biotechnology. 29 (8): 695–6. doi:10.1038/nbt.1940. PMID 21822240. S2CID 13525337.
- Hockemeyer D, Wang H, Kiani S, Lai CS, Gao Q, Cassady JP, et al. (July 2011). "Genetic engineering of human pluripotent cells using TALE nucleases". Nature Biotechnology. 29 (8): 731–4. doi:10.1038/nbt.1927. PMC 3152587. PMID 21738127.
- Wood AJ, Lo TW, Zeitler B, Pickle CS, Ralston EJ, Lee AH, et al. (July 2011). "Targeted genome editing across species using ZFNs and TALENs". Science. 333 (6040): 307. Bibcode:2011Sci...333..307W. doi:10.1126/science.1207773. PMC 3489282. PMID 21700836.
Further reading
- Schornack S, Boch J (2010). "Unraveling a 20-year enigma". IS-MPMI Reporter. International Society for Molecular Plant-Microbe Interactions (1): 3–4.
- Goodsell D (December 2014). "TAL Effectors". Protein Data Bank (PDB) Molecule of the Month. Research Collaboratory for Structural Bioinformatics (RCSB). doi:10.2210/rcsb_pdb/mom_2014_12.
External links
- "A comprehensive, publicly available resource for engineered TAL effector technology". TALengineering.org.
- "TALengineering newsgroup".
Newsgroup for discussion of engineered TAL effector technology
- "An open resource for TAL effector constructs". www.taleffectors.com.