Ultrastenos
Ultrastenos is an extinct genus of Australian mekosuchine crocodilian first described in 2016. The type species Ultrastenos willisi was discovered at Riversleigh in northwestern Queensland, Australia, and lived during the Late Oligocene era. Following its discovery, it was speculated that Ultrastenos was a slender-snouted animal similar to modern gharials or freshwater crocodiles and that it may have inhabited forest pools and fed on small vertebrates like frogs and lizards. This is based on the peculiar shape of its mandible, which is wide towards the base of the head but constricts rapidly, leading into a narrow and gracile rostrum. Assuming that the holotype skull belonged to an adult individual, Ultrastenos may have been a rather small animal, approximately the size of a modern freshwater crocodile. However, Ultrastenos is only known from very fragmentary remains and thus among the most enigmatic mekosuchines. In a 2023 study multiple authors argue that the fragmentary nature of the animal means that further studies are required to truly test the hypothesis proposed in its original description, especially following the discovery of what is thought to have been a close relative.
| Ultrastenos Temporal range: Late Oligocene, | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Order: | Crocodilia |
| Clade: | †Mekosuchinae |
| Genus: | †Ultrastenos Stein et al., 2016 |
| Type species | |
| Ultrastenos willisi Stein et al., 2018 | |
Discovery and naming
Ultrastenos was described in 2016 based on the holotype specimen QM F42665, a posterior cranium and mandible found at the Low Lion Site of the Riversleigh World Heritage Area. While Riversleigh is known to have been home to a variety of other mekosuchine genera, the discovery of Ultrastenos represented a previously unknown morphology of the native Oligocene to Miocene crocodilian fauna. More specifically, Ultrastenos was the first known Riversleigh crocodilian to display what may have been highly elongated jaws similar to those of modern gharials. In addition to the holotype material, multiple fossil remains were assigned as the paratypes. These remains cover various elements of the postcranial skeleton, including multiple vertebrae of the neck and tail, some osteoderms, a coracoid and limb bones. In addition to the Low Lion material, Stein et al. also suggests that material previously collected and described by Willis from the White Hunter Site could have also belonged to this genus.[2][3] This is however contradicted by Adam Yates, who assigns the White Hunter cranium to Baru wickeni.[4] Many of the interpretations made in the type description were later questioned in a 2023 study on mekosuchines, also involving Stein, which makes mention that a re-evaluation of the material is in preparation and that a related form had been discovered.[5]
The name derives from the Latin "ultra" for extreme and the Greek "stenos" for narrow, chosen to reflect the morphology of the animal's mandible. The species epithet honors Dr. Paul Willis, a prolific paleontologist especially involved in mekosuchine research, having participated in the description of over a dozen mekosuchine species.[2]
Description

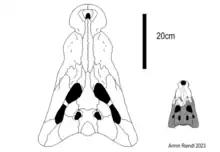
Ultrastenos is unique among mekosuchines thanks to its shallow lower jaw and constricted snout, which was very wide at the back and narrowed rapidly towards the base of the rostrum. As only the back of the skull is known, it is not certain what the snout of Ultrastenos really looked like. One possibility is that the snout was relatively short, stopping not far beyond the point of preservation, which may indicate an altirostral snout form akin to Mekosuchus or modern dwarf caimans and dwarf crocodiles. However, in altirostral forms the mandible generally displays very deep mandibles, which clashes with the anatomy of Ultrastenos. Instead, it is thought to be much more likely that the two halves of the mandible fused to form the mandibular symphysis not far beyond what is known, likely continuing onward to form a longirostrine snout. Such a snout form is seen in a variety of modern crocodilians including gharials, the false gharial, slender snouted crocodiles and freshwater crocodiles. Shared traits between these animals and Ultrastenos include the uniform dentition and shallow mandible. The retroarticular process, while not as elongated as in gavialoids, is similar to that of freshwater crocodiles too.[2] However, in a 2023 study authored by multiple prolific mekosuchine researchers including Stein it is argued that the fragmentary nature of the material means that these interpretations are still questionable.[5]
Skull
The top of the skull table is only known from the White Hunter material, referred as "cranial form 1" by Willis, which is of uncertain affinities. Although Stein et al. refer it to Ultrastenos and base their diagnosis partially on it, Adam Yates would later argue that the fossil actually belonged to Baru wickeni.[4] Whatever the case, the White Hunter cranium displays prominent teardrop-shaped supratemporal fenestra that are unlike those of other mekosuchines. The fenestrae are wider than the postorbital bone and longer than the squamosal, the two bones forming the outer margins of the opening. Overall the skull table is covered in densely packed regular pits that compose most of the ornamentation and at the back end of the element the supraoccipital is exposed and thus visibly in top view. The White Hunter material seems to suggest that the eyesockets were directed laterally, meaning that the eyes possibly faced more towards the sides and not upwards like typical for semi-aquatic crocodilians. This would be similar to mekosuchines like Trilophosuchus and Mekosuchus. The quadrate and quadratojugal are curved inward (medioventrally), especially towards the front where they contact the jugal, which is similarly constricted. Furthermore, the jugal as a whole is gracile and short. The jugal forms the base of the postorbital bar and bears a foramen in this region of the skull.[2][3]
The occiput, the back of the head, is more confidently assigned to Ultrastenos and is wide yet shallow, forming a concave surface that runs perpendicular to the quadrates. The squamosals are not well expressed at the back of the head and the posterolateral processes are only weakly developed, not extending as far back as in other crocodilians. Something similar can be observed with the paroccipital process, a part of the exoccipital, which is likewise poorly developed and rounded, but still clearly set apart from the quadrate and squamosal. The basioccipital, which forms the base of the occiput, is a tall element that's significantly wider at the top and narrows towards the bottom with a prominent keel running down its vertical surface. The foramen magnum, through which the spinal chord passes, appears as an inverted triangle just above the occipital condyle that articulates with the first neck vertebra.[2]
The pterygoids are large elements that form part of the palate. In Ultrastenos the corners of the pterygoids extend beyond the back of the choanae, which itself is surrounded by ridges. Towards the front, it can be observed that the palatal fenestrae were wide, which in turn means that the palatines were narrow.[2]
Mandible

The most striking feature of Ultrastenos is its mandible. It is incredibly gracile and, when looked at from atop, constricts rapidly creating a pronounced curvature in the bone. The back of the lower jaw bulges outwards, but becomes slightly concave towards the front following the constriction. Looking at the lower jaw in profile view also highlights how shallow it is. The dentary is about as tall as the angular and surangular bone, which extend only weakly above and below the dentary following the region where the three bones come into contact. The dentary does grows slightly shallower towards the front, but given the lack of material it is uncertain how that would have continued. The only thing really breaking up this very low profile is the articular process, the connective element between the cranium and lower jaw, which is rather tall. Both angular and surangular become convex at the back of the lower jaw, forming the articular region and an elongated retroarticular process which is notably longer than in any other mekosuchine. The angular and surangular display prominent ridges along their lower and upper ends respectively, the lower ridge being the more developed and forming a buttress, effectively enclosing the ornamented outer surface of the lower jaw. Like that of the skull table, the ornamentation of the mandible is composed of closely spaced pits, although in this region they are much more irregular. The external mandibular fenestra, located at the meeting point of these three major lower jaw bones, appears to have been highly reduced, but not absent.[2]
Given that only the last four tooth socketss and two teeth are preserved, little is known about the dentition of Ultrastenos. It is clear that the tooth row was constricted and, like the lower jaw as a whole, rather narrow. Based on the anatomy of the sockets, these last four teeth all appear to have been similar in size and the known teeth suggest that they were low, slightly compressed from side-to-side and with cutting edges (carinae).[2]
Postcrania
A variety of isolated bones of the postcranial skeleton of Ultrastenos are known, including fragments of the first cervical vertebra, the atlas. The intercentrum, which forms the lower part of the atlas, is tall and both it and the neural arch are elongated in length. Similar proportions can be observed in the tail vertebrae (caudal vertebrae). In addition to this, the faces of the caudals, the part of the centrum that faces the preceding or following vertebra, are expanded downwards, same as the chevrons. The osteoderms forming part of the paratype stem from the dorsal shield, the armour that is situated along the back of the animal, and are moderately robust in morphology with deep pits but lacking a medial ridge. This anatomy is indistinguishable from what is seen in modern crocodiles, and similarly the toe bones are also identical to those of modern species. The preserved portions of the coracoid are likewsie similar to members of the genus Crocodylus, but the proximal head is elongated and forming an acute triangle. The glenoid facet, which is the coracoids contribution to the shoulder joint, is only weakly displaced towards the back of the element. The only difference between the tibia of Ultrastenos and modern crocodiles is that in the mekosuchine it is flattened front to back and the shaft becomes more square as it moves away from the hip.[2]
Size
The skull of Ultrastenos is approximately 16–18 cm (6.3–7.1 in) wide between the quadrates, but it is currently unknown if the material represents a juvenile or an adult individual. Assuming the latter, this would indicate that Ultrastenos a relatively small-bodied animal similar in size to the extant freshwater crocodile, yet still larger than the known mekosuchine dwarf forms such as Trilophosuchus and Mekosuchus.[2]
Phylogeny
Stein et al. recovered Ultrastenos as a mekosuchine crocodilian and sister taxa to Trilophosuchus rackhami, with which it shares a vertical exoccipital. Based on prior phylogenetic analysis this would suggest placement within Mekosuchini, however in Stein et al.'s analysis Mekosuchini as previously defined has lost cohesion. This is because in the recovered results, Trilophosuchus was recovered as more basal than Kambara, a platyrostral species from the Eocene. Although Ultrastenos is only fragmentary, the issues with the phylogeny are not caused by it and instead the result of the uncertain placement of Harpacochampsa, which according to some may be a gavialoid.[2] Another study discussing the phylogenetic position of Ultrastenos was that of Rio and Mannion, published in 2021. Their study argued that many previously established mekosuchines were not mekosuchines at all but instead belonged to different groups. Ultrastenos did maintain its status as a mekosuchine, but was recovered in a polytomy alongside the various species of Mekosuchus, with Trilophosuchus being placed as said groups sister taxon.[1]
|
|
However, due to its fragmentary nature Ultrastenos is commonly not included in phylogenetic analysis. For instance, Ultrastenos was not included by Lee and Yates in their 2019 study, which recovered Mekosuchinae as being markedly different from both the results of Stein et al. and Rio and Mannion.[6] Similar results were again recovered by Ristevski et al. in 2023, which although discussing Ultrastenos did not include it in its phylogeny. Ristevski and colleagues make specific mention of the uncertain nature of Ultrastenos and the fact that it requires significant revisions, mentioning that material of a close relative had been found.[5]
Paleobiology
The narrow mandible and possibly longirostrine nature of Ultrastenos suggests it inhabited a niche previously not known from Riversleigh. Typically narrow, elongated jaws in crocodylomorphs are associated with a diet heavily featuring fish, as examplified by the modern gharial. However fish fossils are not commonly found in Oligocene Riversleigh sites and are less common than frogs, snakes and lizards. Subsequently, Stein et al. suggest that rather than being a piscivore, Ultrastenos may have fed on small reptiles and amphibians. Regardless of prey preference, the shallow mandibles of Ultrastenos may have served the same function as in modern longirostrine crocodilians, which is to streamline movement of the rostrum against the surrounding water, while the uniform teeth would be suited to catch, hold and crush small prey. Furthermore, it can be inferred based on how gracile the jaws were that Ultrastenos was incapable of tackling large prey as done by Baru.[2][3]
During the Late Oligocene the Low Lion Site was predominantly covered by open forests. The presence of a longirostrine crocodilian would suggest the presence of bodies of water, but Stein et al. argue that there is no evidence for extensive riversystems or wetlands from this region. Instead, Ultrastenos may have been more at home in forest lakes and ponds that were drained through the underlying karstic limestone. This may find support in the type of environments nowadays inhabited by freshwater crocodiles, which may persist in bodies of water that are only temporary and not available all year round. Following the interpretation that the cranium described by Willis also belonged to Ultrastenos, as suggested by Stein et al., then the genus would have also inhabited the White Hunter Site which is well known for its diverse crocodile fauna. If the White Hunter cranium truly belonged to Ultrastenos, then it may have lived alongside the terrestrial Mekosuchus whitehunterensis and Quinkana meboldi, the small bodied semi-aquatic "Baru" huberi and the much larger macropredator Baru wickeni.[2][3]
References
- Rio, Jonathan P.; Mannion, Philip D. (6 September 2021). "Phylogenetic analysis of a new morphological dataset elucidates the evolutionary history of Crocodylia and resolves the long-standing gharial problem". PeerJ. 9: e12094. doi:10.7717/peerj.12094. PMC 8428266. PMID 34567843.
- Stein, Michael; Hand, Suzanne J.; Archer, Michael (26 May 2016). "A new crocodile displaying extreme constriction of the mandible, from the late Oligocene of Riversleigh, Australia". Journal of Vertebrate Paleontology. 36 (5): e1179041. doi:10.1080/02724634.2016.1179041. S2CID 88895724.
- Willis, P.M.A. (1997). "New crocodilians from the late Oligocene White Hunter Site, Riversleigh, northwestern Queensland". Memoirs of the Queensland Museum. 41: 423–438. ISSN 0079-8835.
- Yates, A.M. (2017). "The biochronology and palaeobiogeography of Baru (Crocodylia: Mekosuchinae) based on new specimens from the Northern Territory and Queensland, Australia". PeerJ. 5.
- Ristevski, J.; Willis, P.M.A.; Yates, A.M.; White, M.A.; Hart, L.J.; Stein, M.D.; Price, G.J.; Salisbury, S.W. (2023). "Migrations, diversifications and extinctions: the evolutionary history of crocodyliforms in Australasia". Alcheringa: An Australasian Journal of Palaeontology: 1–46. doi:10.1080/03115518.2023.2201319. S2CID 258878554.
- Michael S. Y. Lee; Adam M. Yates (27 June 2018). "Tip-dating and homoplasy: reconciling the shallow molecular divergences of modern gharials with their long fossil". Proceedings of the Royal Society B. 285 (1881). doi:10.1098/rspb.2018.1071. PMC 6030529. PMID 30051855.
Extinct crocodilians | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||


