Adenylate kinase
Adenylate kinase (EC 2.7.4.3) (also known as ADK or myokinase) is a phosphotransferase enzyme that catalyzes the interconversion of the various adenosine phosphates (ATP, ADP, and AMP). By constantly monitoring phosphate nucleotide levels inside the cell, ADK plays an important role in cellular energy homeostasis.
| ADK_lid | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Bacillus stearothermophilus adenylate kinase | |||||||||
| Identifiers | |||||||||
| Symbol | ADK_lid | ||||||||
| Pfam | PF05191 | ||||||||
| InterPro | IPR007862 | ||||||||
| PROSITE | PDOC00104 | ||||||||
| SCOP2 | 1ake / SCOPe / SUPFAM | ||||||||
| |||||||||
| Adenylate kinase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC no. | 2.7.4.3 | ||||||||
| CAS no. | 2598011 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
| Adenylate kinase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
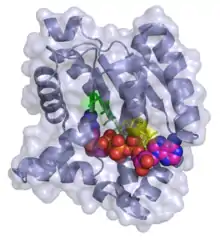 3D ribbon/surface model of adenylate kinase in complex with bis(adenosine)tetraphosphate (ADP-ADP) | |||||||||
| Identifiers | |||||||||
| Symbol | ADK | ||||||||
| Pfam | PF00406 | ||||||||
| InterPro | IPR000850 | ||||||||
| PROSITE | PDOC00104 | ||||||||
| SCOP2 | 1ake / SCOPe / SUPFAM | ||||||||
| |||||||||
Substrate and products
The reaction catalyzed is:
The equilibrium constant varies with condition, but it is close to 1.[1] Thus, ΔGo for this reaction is close to zero. In muscle from a variety of species of vertebrates and invertebrates, the concentration of ATP is typically 7-10 times that of ADP, and usually greater than 100 times that of AMP.[2] The rate of oxidative phosphorylation is controlled by the availability of ADP. Thus, the mitochondrion attempts to keep ATP levels high due to the combined action of adenylate kinase and the controls on oxidative phosphorylation.
Isozymes
To date there have been nine human ADK protein isoforms identified. While some of these are ubiquitous throughout the body, some are localized into specific tissues. For example, ADK7 and ADK8 are both only found in the cytosol of cells; and ADK7 is found in skeletal muscle whereas ADK8 is not.[3] Not only do the locations of the various isoforms within the cell vary, but the binding of substrate to the enzyme and kinetics of the phosphoryl transfer are different as well. ADK1, the most abundant cytosolic ADK isozyme, has a Km about a thousand times higher than the Km of ADK7 and 8, indicating a much weaker binding of ADK1 to AMP.[4] Sub-cellular localization of the ADK enzymes is done by including a targeting sequence in the protein.[3] Each isoform also has different preference for NTP's. Some will only use ATP, whereas others will accept GTP, UTP, and CTP as the phosphoryl carrier.
Some of these isoforms prefer other NTP's entirely. There is a mitochondrial GTP:AMP phosphotransferase, also specific for the phosphorylation of AMP, that can only use GTP or ITP as the phosphoryl donor.[5] ADK has also been identified in different bacterial species and in yeast.[6] Two further enzymes are known to be related to the ADK family, i.e. yeast uridine monophosphokinase and slime mold UMP-CMP kinase. Some residues are conserved across these isoforms, indicating how essential they are for catalysis. One of the most conserved areas includes an Arg residue, whose modification inactivates the enzyme, together with an Asp that resides in the catalytic cleft of the enzyme and participates in a salt bridge.
Subfamilies
Mechanism
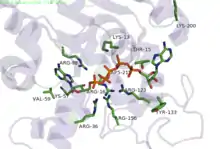
Phosphoryl transfer only occurs on closing of the 'open lid'. This causes an exclusion of water molecules that brings the substrates in proximity to each other,[7] lowering the energy barrier for the nucleophilic attack by the α-phosphoryl of AMP on the γ-phosphoryl group of ATP resulting in formation of ADP by transfer of the γ-phosphoryl group to AMP. In the crystal structure of the ADK enzyme from E. coli with inhibitor Ap5A, the Arg88 residue binds the Ap5A at the α-phosphate group. It has been shown that the mutation R88G results in 99% loss of catalytic activity of this enzyme, suggesting that this residue is intimately involved in the phosphoryl transfer.[8] Another highly conserved residue is Arg119, which lies in the adenosine binding region of the ADK, and acts to sandwich the adenine in the active site. It has been suggested that the promiscuity of these enzymes in accepting other NTP's is due to this relatively inconsequential interactions of the base in the ATP binding pocket.[9] A network of positive, conserved residues (Lys13, Arg123, Arg156, and Arg167 in ADK from E. coli) stabilize the buildup of negative charge on phosphoryl group during the transfer. Two distal aspartate residues bind to the arginine network, causing the enzyme to fold and reduces its flexibility. A magnesium cofactor is also required, essential for increasing the electrophilicity of the phosphate on AMP, though this magnesium ion is only held in the active pocket by electrostatic interactions and dissociates easily.[9]
Structure

Flexibility and plasticity allow proteins to bind to ligands, form oligomers, aggregate, and perform mechanical work.[10] Large conformational changes in proteins play an important role in cellular signaling. Adenylate Kinase is a signal transducing protein; thus, the balance between conformations regulates protein activity. ADK has a locally unfolded state that becomes depopulated upon binding.[11]
A 2007 study by Whitford et al. shows the conformations of ADK when binding with ATP or AMP.[10] The study shows that there are three relevant conformations or structures of ADK—CORE, Open, and Closed. In ADK, there are two small domains called the LID and NMP.[12] ATP binds in the pocket formed by the LID and CORE domains. AMP binds in the pocket formed by the NMP and CORE domains. The Whitford study also reported findings that show that localized regions of a protein unfold during conformational transitions. This mechanism reduces the strain and enhances catalytic efficiency. Local unfolding is the result of competing strain energies in the protein.[10]
The local (thermodynamic) stability of the substrate-binding domains ATPlid and AMPlid has been shown to be significantly lower when compared with the CORE domain in ADKE. coli.[13] Furthermore, it has been shown that the two subdomains (ATPlid and AMPlid) can fold and unfold in a "non-cooperative manner."[13] Binding of the substrates causes preference for 'closed' conformations amongst those that are sampled by ADK. These 'closed' conformations are hypothesized to help with removal of water from the active site to avoid wasteful hydrolysis of ATP in addition to helping optimize alignment of substrates for phosphoryl-transfer.[14] Furthermore, it has been shown that the apoenzyme will still sample the 'closed' conformations of the ATPlid and AMPlid domains in the absence of substrates.[7] When comparing the rate of opening of the enzyme (which allows for product release) and the rate of closing that accompanies substrate binding, closing was found to be the slower process.
Function
Metabolic monitoring
The ability for a cell to dynamically measure energetic levels provides it with a method to monitor metabolic processes.[15] By continually monitoring and altering the levels of ATP and the other adenyl phosphates (ADP and AMP levels) adenylate kinase is an important regulator of energy expenditure at the cellular level.[16] As energy levels change under different metabolic stresses adenylate kinase is then able to generate AMP; which itself acts as a signaling molecule in further signaling cascades. This generated AMP can, for example, stimulate various AMP-dependent receptors such as those involved in glycolytic pathways, K-ATP channels, and 5' AMP-activated protein kinase (AMPK).[15] Common factors that influence adenine nucleotide levels, and therefore ADK activity are exercise, stress, changes in hormone levels, and diet.[15] It facilitates decoding of cellular information by catalyzing nucleotide exchange in the intimate “sensing zone” of metabolic sensors.[15]
ADK shuttle
Adenylate kinase is present in mitochondrial and myofibrillar compartments in the cell, and it makes two high-energy phosphoryls (β and γ) of ATP available to be transferred between adenine nucleotide molecules.[15][16] In essence, adenylate kinase shuttles ATP to sites of high energy consumption and removes the AMP generated over the course of those reactions. These sequential phosphotransfer relays ultimately result in propagation of the phosphoryl groups along collections of ADK molecules.[15] This process can be thought of as a bucket brigade of ADK molecules that results in changes in local intracellular metabolic flux without apparent global changes in metabolite concentrations.[15] This process is extremely important for overall homeostasis of the cell.[15]
Disease relevance
Nucleoside diphosphate kinase deficiency
Nucleoside diphosphate (NDP) kinase catalyzes in vivo ATP-dependent synthesis of ribo- and deoxyribonucleoside triphosphates. In mutated Escherichia coli that had a disrupted nucleoside diphosphate kinase, adenylate kinase performed dual enzymatic functions. ADK complements nucleoside diphosphate kinase deficiency.[17]
Nucleoside diphosphate (NDP) kinase catalyzes in vivo ATP-dependent synthesis of ribo- and deoxyribonucleoside triphosphates. In mutated Escherichia coli that had a disrupted nucleoside diphosphate kinase, adenylate kinase performed dual enzymatic functions. ADK complements nucleoside diphosphate kinase deficiency.[17]
AK1 and post-ischemic coronary reflow
Knock out of AK1 disrupts the synchrony between inorganic phosphate and turnover at ATP-consuming sites and ATP synthesis sites. This reduces the energetic signal communication in the post-ischemic heart and precipitates inadequate coronary reflow following ischemia-reperfusion.[18]
ADK2 deficiency
Adenylate Kinase 2 (AK2) deficiency in humans causes hematopoietic defects associated with sensorineural deafness.[19] Reticular dysgenesis is an autosomal recessive form of human combined immunodeficiency. It is also characterized by an impaired lymphoid maturation and early differentiation arrest in the myeloid lineage. AK2 deficiency results in absent or a large decrease in the expression of proteins. AK2 is specifically expressed in the stria vascularis of the inner ear which indicates why individuals with an AK2 deficiency will have sensorineural deafness.[19]
Structural adaptations
AK1 genetic ablation decreases tolerance to metabolic stress. AK1 deficiency induces fiber-type specific variation in groups of transcripts in glycolysis and mitochondrial metabolism.[20] This supports muscle energy metabolism.
Plastidial ADK deficiency in Arabidopsis thaliana
Enhanced growth and elevated photosynthetic amino acid is associated with plastidial adenylate kinase deficiency in Arabidopsis thaliana.[21]
References
- The NIST Thermodynamics of Enzyme-Catalyzed Reactions database, http://xpdb.nist.gov/enzyme_thermodynamics/enzyme1.pl, Goldberg RN, Tewari YB, Bhat TN (November 2004). "Thermodynamics of enzyme-catalyzed reactions--a database for quantitative biochemistry". Bioinformatics. 20 (16): 2874–7. doi:10.1093/bioinformatics/bth314. PMID 15145806., gives equilibrium constants, search for adenylate kinase under enzymes
- Beis I, Newsholme EA (October 1975). "The contents of adenine nucleotides, phosphagens and some glycolytic intermediates in resting muscles from vertebrates and invertebrates". The Biochemical Journal. 152 (1): 23–32. doi:10.1042/bj1520023. PMC 1172435. PMID 1212224.
- Panayiotou C, Solaroli N, Karlsson A (April 2014). "The many isoforms of human adenylate kinases". The International Journal of Biochemistry & Cell Biology. 49: 75–83. doi:10.1016/j.biocel.2014.01.014. PMID 24495878.
- Panayiotou C, Solaroli N, Xu Y, Johansson M, Karlsson A (February 2011). "The characterization of human adenylate kinases 7 and 8 demonstrates differences in kinetic parameters and structural organization among the family of adenylate kinase isoenzymes" (PDF). The Biochemical Journal. 433 (3): 527–34. doi:10.1042/BJ20101443. PMID 21080915.
- Tomasselli AG, Noda LH (January 1979). "Mitochondrial GTP-AMP phosphotransferase. 2. Kinetic and equilibrium dialysis studies". European Journal of Biochemistry. 93 (2): 263–7. doi:10.1111/j.1432-1033.1979.tb12819.x. PMID 218813.
- Cooper AJ, Friedberg EC (May 1992). "A putative second adenylate kinase-encoding gene from the yeast Saccharomyces cerevisiae". Gene. 114 (1): 145–8. doi:10.1016/0378-1119(92)90721-Z. PMID 1587477.
- Henzler-Wildman KA, Thai V, Lei M, Ott M, Wolf-Watz M, Fenn T, Pozharski E, Wilson MA, Petsko GA, Karplus M, Hübner CG, Kern D (December 2007). "Intrinsic motions along an enzymatic reaction trajectory". Nature. 450 (7171): 838–44. Bibcode:2007Natur.450..838H. doi:10.1038/nature06410. PMID 18026086. S2CID 4406037.
- Reinstein J, Gilles AM, Rose T, Wittinghofer A, Saint Girons I, Bârzu O, Surewicz WK, Mantsch HH (May 1989). "Structural and catalytic role of arginine 88 in Escherichia coli adenylate kinase as evidenced by chemical modification and site-directed mutagenesis". The Journal of Biological Chemistry. 264 (14): 8107–12. doi:10.1016/S0021-9258(18)83156-8. PMID 2542263.
- Müller CW, Schulz GE (March 1992). "Structure of the complex between adenylate kinase from Escherichia coli and the inhibitor Ap5A refined at 1.9 A resolution. A model for a catalytic transition state". Journal of Molecular Biology. 224 (1): 159–77. doi:10.2210/pdb1ake/pdb. PMID 1548697.
- Whitford PC, Miyashita O, Levy Y, Onuchic JN (March 2007). "Conformational transitions of adenylate kinase: switching by cracking". Journal of Molecular Biology. 366 (5): 1661–71. doi:10.1016/j.jmb.2006.11.085. PMC 2561047. PMID 17217965.
- Schrank TP, Bolen DW, Hilser VJ (October 2009). "Rational modulation of conformational fluctuations in adenylate kinase reveals a local unfolding mechanism for allostery and functional adaptation in proteins". Proceedings of the National Academy of Sciences of the United States of America. 106 (40): 16984–9. Bibcode:2009PNAS..10616984S. doi:10.1073/pnas.0906510106. PMC 2761315. PMID 19805185.
- Daily MD, Phillips GN, Cui Q (July 2010). "Many local motions cooperate to produce the adenylate kinase conformational transition". Journal of Molecular Biology. 400 (3): 618–31. doi:10.1016/j.jmb.2010.05.015. PMC 2902635. PMID 20471396.
- Rundqvist L, Adén J, Sparrman T, Wallgren M, Olsson U, Wolf-Watz M (March 2009). "Noncooperative folding of subdomains in adenylate kinase". Biochemistry. 48 (9): 1911–27. doi:10.1021/bi8018042. PMID 19219996.
- Olsson U, Wolf-Watz M (November 2010). "Overlap between folding and functional energy landscapes for adenylate kinase conformational change". Nature Communications. 1 (8): 111. Bibcode:2010NatCo...1..111O. doi:10.1038/ncomms1106. PMID 21081909.
- Dzeja P, Terzic A (April 2009). "Adenylate kinase and AMP signaling networks: metabolic monitoring, signal communication and body energy sensing". International Journal of Molecular Sciences. 10 (4): 1729–72. doi:10.3390/ijms10041729. PMC 2680645. PMID 19468337.
- Dzeja PP, Chung S, Faustino RS, Behfar A, Terzic A (April 2011). "Developmental enhancement of adenylate kinase-AMPK metabolic signaling axis supports stem cell cardiac differentiation". PLOS ONE. 6 (4): e19300. Bibcode:2011PLoSO...619300D. doi:10.1371/journal.pone.0019300. PMC 3083437. PMID 21556322.
- Lu Q, Inouye M (June 1996). "Adenylate kinase complements nucleoside diphosphate kinase deficiency in nucleotide metabolism". Proceedings of the National Academy of Sciences of the United States of America. 93 (12): 5720–5. Bibcode:1996PNAS...93.5720L. doi:10.1073/pnas.93.12.5720. PMC 39127. PMID 8650159.
- Dzeja PP, Bast P, Pucar D, Wieringa B, Terzic A (October 2007). "Defective metabolic signaling in adenylate kinase AK1 gene knock-out hearts compromises post-ischemic coronary reflow". The Journal of Biological Chemistry. 282 (43): 31366–72. doi:10.1074/jbc.M705268200. PMC 3232003. PMID 17704060.
- Lagresle-Peyrou C, Six EM, Picard C, Rieux-Laucat F, Michel V, Ditadi A, Demerens-de Chappedelaine C, Morillon E, Valensi F, Simon-Stoos KL, Mullikin JC, Noroski LM, Besse C, Wulffraat NM, Ferster A, Abecasis MM, Calvo F, Petit C, Candotti F, Abel L, Fischer A, Cavazzana-Calvo M (January 2009). "Human adenylate kinase 2 deficiency causes a profound hematopoietic defect associated with sensorineural deafness". Nature Genetics. 41 (1): 106–11. doi:10.1038/ng.278. PMC 2612090. PMID 19043416.
- Janssen E, de Groof A, Wijers M, Fransen J, Dzeja PP, Terzic A, Wieringa B (April 2003). "Adenylate kinase 1 deficiency induces molecular and structural adaptations to support muscle energy metabolism". The Journal of Biological Chemistry. 278 (15): 12937–45. doi:10.1074/jbc.M211465200. PMID 12562761.
- Carrari F, Coll-Garcia D, Schauer N, Lytovchenko A, Palacios-Rojas N, Balbo I, Rosso M, Fernie AR (January 2005). "Deficiency of a plastidial adenylate kinase in Arabidopsis results in elevated photosynthetic amino acid biosynthesis and enhanced growth". Plant Physiology. 137 (1): 70–82. doi:10.1104/pp.104.056143. PMC 548839. PMID 15618410.
External links
- Adenylate+kinase at the US National Library of Medicine Medical Subject Headings (MeSH)