CD80
The Cluster of differentiation 80 (also CD80 and B7-1) is a B7, type I membrane protein[5] in the immunoglobulin superfamily, with an extracellular immunoglobulin constant-like domain and a variable-like domain required for receptor binding. It is closely related to CD86, another B7 protein (B7-2), and often works in tandem. Both CD80 and CD86 interact with costimulatory receptors CD28 and CTLA-4 (CD152).[6][7]
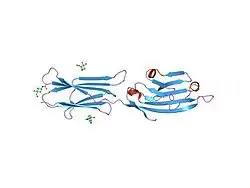




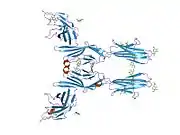
Structure
CD80 is a member of the B7 family, which consists of molecules present at APCs and their receptors present on the T-cells.[7] CD80 is present specifically on DC, activated B-cells, and macrophages, but also T-cells[7][8] CD80 is also a transmembrane glycoprotein and a member of the Ig superfamily.[7] It is composed of 288 amino acids, and its mass is 33 kDa.[8] It consists of two Ig-like extracellular domains (208 AA), a transmembrane helical segment (21 AA), and a short cytoplasmic tail (25 AA).[7][8][9] The Ig-like extracellular domains are formed by single V-type and C2-type domains.[7][6][10] It is expressed as both monomers or dimers, but predominantly dimmers.[7][10][11] These two forms exist in dynamic equilibrium.[12]
CD80 shares 25% of sequences with CD86; however, CD80 has a ten-fold higher affinity for CD28 and CTLA-4 than CD86. Moreover, CD80 interacts with its ligand with faster binding kinetics and slower dissociation constants than CD86. Both human CD80 and CD86 are located at chromosome 3; the exact region is 3q13.3-q21.[7]
Human and murine CD80 share approximately 44% of sequences. Also both human and murine CD80 are able to cross-react with both human and murine CD28. This indicates that the binding site of CD80 is conserved.[7][12]
Function
CD80 can be found on the surface of various immune cells, including B-cells, monocytes, or T-cells, but most typically at antigen-presenting cells (APCs) such as dendritic cells.[6][7][13] CD80 has a crucial role in modulating T-cell immune function as a checkpoint protein at the immunological synapse.[14]
CD80 is the ligand for the proteins CD28 (for autoregulation and intercellular association) and CTLA-4 (for attenuation of regulation and cellular disassociation) found on the surface of T-cells.[6][13] Interaction of CD80 with CD28 triggers costimulatory signals and results in enhanced and sustained T-cell activation. In contrast, contrary interaction of CD80 with CTLA-4 inhibits parts of T-cell effector function. These two ligands are structurally homologous, and they compete with each other for binding sites.[14] However, the bond with CTLA-4 has up to 2500 fold higher avidity than with CD28.[7] This illustrates that inhibitory interaction with CTLA-4 is predominant.[14]
CD80 binds to CD28 and CTLA-4 with lower affinity and fast binding kinetics (Kd = 4 μM for CD28 and 0.42 μM for CTLA-4), allowing for quick interactions between the communicating cells.[15] These interactions result in an important costimulatory signal in the immunological synapse between antigen-presenting cells, B-cells, dendritic cells and T-cells that result in T and B-cell activation, proliferation and differentiation.[11]
When stimulated by CD80, T helper cells preferentially differentiate into Th1 cells.[11] CD80 is an essential component in dendritic cell licensing and cytotoxic T-cell activation. When the major histocompatibility complex class II (MHC class II)-peptide complex on a dendritic cell interacts with the receptor on a T helper cell, CD80 is up-regulated, licensing the dendritic cell and allowing for interaction between the dendritic cell and CD 8+ T-cells via CD28. This helps to signal the T-cell differentiation into a cytotoxic T-cell.[13][16] The expression of CD80, as well as CD86, is increased by the presence of microbes and cytokines, which is the consequence of the presence of microbes. This mechanism ensures that costimulatory molecules for T-cells are present at the right time.[7]
CD80, often in tandem with CD86, plays a large and diverse role in regulating both the adaptive and the innate immune system. As mentioned above, this protein is crucial for immune cell activation in response to pathogens. The interaction of CD80 with CD28, together with TCR and MHC interaction, results in activation of nuclear factor‐κB (NF-ⲕB), mitogen‐activated protein kinase (MAPK), and the calcium‐calcineurin pathway. These changes initiate the production of numerous factors, cytokines, and chemokines by T-cells. Noteworthy is the production of interleukin 2 (IL-2) as well as ɑ-chain of CD25 (which is a receptor of IL-2), CD40 ligand, tumor necrosis factor‐α (TNFα), TNF‐β, and interferon‐γ (IFN‐γ). T-cells also increase the production of macrophage inflammatory proteins 1α and 1β (MIP‐α1 and MIP‐1β) and prevent apoptosis by induction of anti-apoptotic protein expression (e.g., Bcl‐X and Bcl‐2).[14][17][18][19][20] CD80 interaction with CD28 also further stimulates dendritic cells, enhancing cytokine production, specifically IL-6, a pro-inflammatory molecule.[21][22] Neutrophils can also activate macrophages with CD80 via CD28.[22] Last but not least, the interaction of CD80 and CD28 enhances cell‐cycle progression by upregulating the expression levels of D‐cyclin.[14]
In contrast to the stimulatory interaction with CD28, CD80 also regulates the immune system through an inhibitory interaction with CTLA-4. Dendritic cells are suppressed by a CTLA-4-CD80 interaction,[22] and this interaction also promotes the suppressive effects of regulatory T cells, which can prevent an immune response to self-antigen.[18]
In addition to interactions with CD28 and CTLA-4, CD80 is also thought to interact with a separate ligand on Natural Killer cells, triggering the Natural Killer cell-mediated cell death of the CD80 carrier.[23] CD80 may also play a role in the negative regulation of effector and memory T-cells. If the interaction between an antigen-presenting cell and a T-cell is stable enough, the T-cell can remove the CD80 from the antigen-presenting cell. Under the right conditions, this transfer of the CD80 may induce T-cell apoptosis.[24] Finally, CD80 signaling on activated B-cells may regulate antibody secretion during infection.[25]
Another ligand of CD80 is programmed death-ligand 1 (PD‐L1), expressed on the surface of T-cells, B-cells, DCs, and macrophages. This interaction is inhibiting and causes a reduction in T-cell activation as well as reduction of cytokine production. Its dissociation constant with CD80 is between the CD28 and CTLA-40 (Kd = 1.4 μM).[14][26]
Clinical significance
The complicated role CD80 plays in immune system regulation presents an opportunity for CD80 interactions to go rogue in various diseases. The up-regulation of CD80 has been linked to various autoimmune diseases, including multiple sclerosis,[27] systemic lupus erythematosus[28] and sepsis[29] (which may partly be due to over-active T-cells), and CD80 has also been shown to help spread of HIV infection in the body.[30] CD80 is also linked to various cancers, though some experience CD80 induced tolerance via possible regulatory T-cell interaction.[31] Others experience inhibited growth and metastasis-related to CD80 up-regulation,[32] further exemplifies the complicated role CD80 plays.
The triggering of Natural Killer cell-mediated death via CD80 interactions has been explored as possible cancer immunotherapy by inducing CD80 expression on tumor cells.[23]
See also
- Cluster of differentiation
- CD86
- CD28
References
- GRCh38: Ensembl release 89: ENSG00000121594 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000075122 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- McKusick, V. A., & Converse, P. J. (2016, August 05). CD80 Antigen; CD80. Retrieved May 29, 2019
- Peach RJ, Bajorath J, Naemura J, Leytze G, Greene J, Aruffo A, Linsley PS (September 1995). "Both extracellular immunoglobin-like domains of CD80 contain residues critical for binding T cell surface receptors CTLA-4 and CD28". The Journal of Biological Chemistry. 270 (36): 21181–7. doi:10.1074/jbc.270.36.21181. PMID 7545666.
- Mir MA (2015). "Introduction to Costimulation and Costimulatory Molecules". Developing Costimulatory Molecules for Immunotherapy of Diseases. Elsevier: 1–43. doi:10.1016/b978-0-12-802585-7.00001-7. ISBN 978-0-12-802585-7.
- "CD80 - T-lymphocyte activation antigen CD80 precursor - Homo sapiens (Human) - CD80 gene & protein". www.uniprot.org. Retrieved 2021-06-09.
- Abbas AK (2021). Cellular and molecular immunology. ISBN 978-0-323-75749-2. OCLC 1173994133.
- Bhatia S, Edidin M, Almo SC, Nathenson SG (April 2006). "B7-1 and B7-2: similar costimulatory ligands with different biochemical, oligomeric and signaling properties". Immunology Letters. 104 (1–2): 70–5. doi:10.1016/j.imlet.2005.11.019. PMID 16413062.
- Bhatia S, Edidin M, Almo SC, Nathenson SG (October 2005). "Different cell surface oligomeric states of B7-1 and B7-2: implications for signaling". Proceedings of the National Academy of Sciences of the United States of America. 102 (43): 15569–74. Bibcode:2005PNAS..10215569B. doi:10.1073/pnas.0507257102. PMC 1266120. PMID 16221763.
- Ikemizu S, Gilbert RJ, Fennelly JA, Collins AV, Harlos K, Jones EY, et al. (January 2000). "Structure and dimerization of a soluble form of B7-1". Immunity. 12 (1): 51–60. doi:10.1016/s1074-7613(00)80158-2. PMID 10661405.
- Owen JA, Punt J, Stranford SA, Jones PP, Kuby J (2013). Kuby Immunology (7th ed.). New York: W.H. Freeman and Company.
- Chen R, Ganesan A, Okoye I, Arutyunova E, Elahi S, Lemieux MJ, Barakat K (March 2020). "Targeting B7-1 in immunotherapy". Medicinal Research Reviews. 40 (2): 654–682. doi:10.1002/med.21632. PMID 31448437. S2CID 201748060.
- van der Merwe PA, Bodian DL, Daenke S, Linsley P, Davis SJ (February 1997). "CD80 (B7-1) binds both CD28 and CTLA-4 with a low affinity and very fast kinetics". The Journal of Experimental Medicine. 185 (3): 393–403. doi:10.1084/jem.185.3.393. PMC 2196039. PMID 9053440.
- Fujii S, Liu K, Smith C, Bonito AJ, Steinman RM (June 2004). "The linkage of innate to adaptive immunity via maturing dendritic cells in vivo requires CD40 ligation in addition to antigen presentation and CD80/86 costimulation". The Journal of Experimental Medicine. 199 (12): 1607–18. doi:10.1084/jem.20040317. PMC 2212806. PMID 15197224.
- Snanoudj R, Frangié C, Deroure B, François H, Créput C, Beaudreuil S, et al. (September 2007). "The blockade of T-cell co-stimulation as a therapeutic stratagem for immunosuppression: Focus on belatacept". Biologics: Targets and Therapy. 1 (3): 203–13. PMC 2721321. PMID 19707331.
- Zheng Y, Manzotti CN, Liu M, Burke F, Mead KI, Sansom DM (March 2004). "CD86 and CD80 differentially modulate the suppressive function of human regulatory T cells". Journal of Immunology. 172 (5): 2778–84. doi:10.4049/jimmunol.172.5.2778. PMID 14978077.
- Boise LH, Minn AJ, Noel PJ, June CH, Accavitti MA, Lindsten T, Thompson CB (July 1995). "CD28 costimulation can promote T cell survival by enhancing the expression of Bcl-XL". Immunity. 3 (1): 87–98. doi:10.1016/1074-7613(95)90161-2. PMID 7621080.
- Kovalev GI, Franklin DS, Coffield VM, Xiong Y, Su L (September 2001). "An important role of CDK inhibitor p18(INK4c) in modulating antigen receptor-mediated T cell proliferation". Journal of Immunology. 167 (6): 3285–92. doi:10.4049/jimmunol.167.6.3285. PMC 4435948. PMID 11544316.
- Orabona C, Grohmann U, Belladonna ML, Fallarino F, Vacca C, Bianchi R, et al. (November 2004). "CD28 induces immunostimulatory signals in dendritic cells via CD80 and CD86". Nature Immunology. 5 (11): 1134–42. doi:10.1038/ni1124. PMID 15467723. S2CID 6080497.
- Nolan A, Kobayashi H, Naveed B, Kelly A, Hoshino Y, Hoshino S, et al. (August 2009). "Differential role for CD80 and CD86 in the regulation of the innate immune response in murine polymicrobial sepsis". PLOS ONE. 4 (8): e6600. Bibcode:2009PLoSO...4.6600N. doi:10.1371/journal.pone.0006600. PMC 2719911. PMID 19672303.
- Chambers BJ, Salcedo M, Ljunggren HG (October 1996). "Triggering of natural killer cells by the costimulatory molecule CD80 (B7-1)". Immunity. 5 (4): 311–7. doi:10.1016/S1074-7613(00)80257-5. PMID 8885864.
- Sabzevari H, Kantor J, Jaigirdar A, Tagaya Y, Naramura M, Hodge J, Bernon J, Schlom J (February 2001). "Acquisition of CD80 (B7-1) by T cells". Journal of Immunology. 166 (4): 2505–13. doi:10.4049/jimmunol.166.4.2505. PMID 11160311.
- Rau FC, Dieter J, Luo Z, Priest SO, Baumgarth N (December 2009). "B7-1/2 (CD80/CD86) direct signaling to B cells enhances IgG secretion". Journal of Immunology. 183 (12): 7661–71. doi:10.4049/jimmunol.0803783. PMC 2795108. PMID 19933871.
- Butte MJ, Keir ME, Phamduy TB, Sharpe AH, Freeman GJ (July 2007). "Programmed death-1 ligand 1 interacts specifically with the B7-1 costimulatory molecule to inhibit T cell responses". Immunity. 27 (1): 111–22. doi:10.1016/j.immuni.2007.05.016. PMC 2707944. PMID 17629517.
- Windhagen A, Newcombe J, Dangond F, Strand C, Woodroofe MN, Cuzner ML, Hafler DA (December 1995). "Expression of costimulatory molecules B7-1 (CD80), B7-2 (CD86), and interleukin 12 cytokine in multiple sclerosis lesions". The Journal of Experimental Medicine. 182 (6): 1985–96. doi:10.1084/jem.182.6.1985. PMC 2192240. PMID 7500044.
- Wong CK, Lit LC, Tam LS, Li EK, Lam CW (August 2005). "Aberrant production of soluble costimulatory molecules CTLA-4, CD28, CD80 and CD86 in patients with systemic lupus erythematosus". Rheumatology. Oxford, England. 44 (8): 989–94. doi:10.1093/rheumatology/keh663. PMID 15870153.
- Nolan A, Weiden M, Kelly A, Hoshino Y, Hoshino S, Mehta N, Gold JA (February 2008). "CD40 and CD80/86 act synergistically to regulate inflammation and mortality in polymicrobial sepsis". American Journal of Respiratory and Critical Care Medicine. 177 (3): 301–8. doi:10.1164/rccm.200703-515OC. PMC 2218847. PMID 17989345.
- Pinchuk LM, Polacino PS, Agy MB, Klaus SJ, Clark EA (July 1994). "The role of CD40 and CD80 accessory cell molecules in dendritic cell-dependent HIV-1 infection". Immunity. 1 (4): 317–25. doi:10.1016/1074-7613(94)90083-3. PMID 7534204.
- Yang R, Cai Z, Zhang Y, Yutzy WH, Roby KF, Roden RB (July 2006). "CD80 in immune suppression by mouse ovarian carcinoma-associated Gr-1+CD11b+ myeloid cells". Cancer Research. 66 (13): 6807–15. doi:10.1158/0008-5472.CAN-05-3755. PMID 16818658.
- Imasuen I, Bozeman E, He S, Patel J, Selvaraj P (May 2013). "Increased B7-1 (CD80) expression reduces overall tumorigenicity and metastatic potential of the murine pancreatic cancer cell model Pan02 (P2085)". The Journal of Immunology. 190 (1 Supplement).
External links
- Human CD80 genome location and CD80 gene details page in the UCSC Genome Browser.
- Overview of all the structural information available in the PDB for UniProt: P33681 (T-lymphocyte activation antigen CD80) at the PDBe-KB.
PDB gallery | |
|---|---|
|