Mycobacteriophage
A mycobacteriophage is a member of a group of bacteriophages known to have mycobacteria as host bacterial species. While originally isolated from the bacterial species Mycobacterium smegmatis and Mycobacterium tuberculosis,[2] the causative agent of tuberculosis, more than 4,200 mycobacteriophage have since been isolated from various environmental and clinical sources. 2,042 have been completely sequenced.[3] Mycobacteriophages have served as examples of viral lysogeny and of the divergent morphology and genetic arrangement characteristic of many phage types.[4]
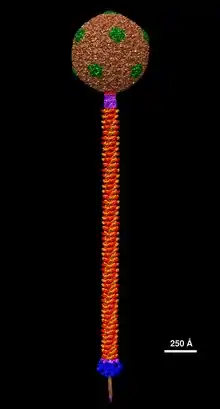
All mycobacteriophages found thus far have had double-stranded DNA genomes and have been classified by their structure and appearance into siphoviridae or myoviridae.[5]
Discovery
A bacteriophage found to infect Mycobacterium smegmatis in 1947 was the first documented example of a mycobacteriophage. It was found in cultures of the bacteria originally growing in moist compost.[6] The first bacteriophage that infects M. tuberculosis was discovered in 1954.[7]
Diversity
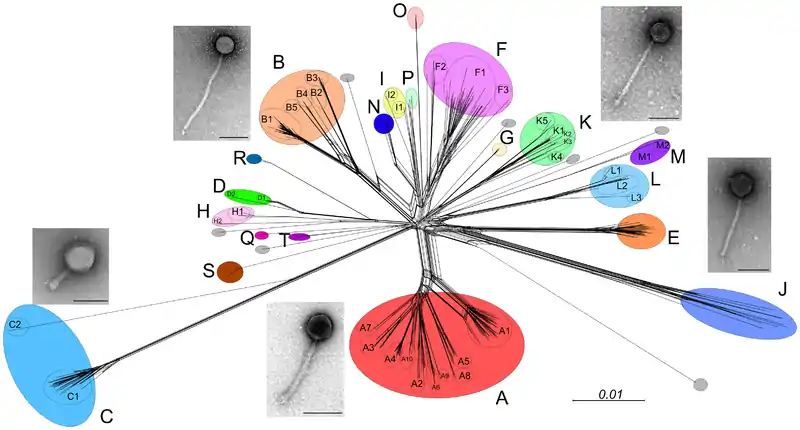
Thousands of mycobacteriophage have been isolated using a single host strain, Mycobacterium smegmatis mc2155, over 1400 of which have been completely sequenced.[3] These are mostly from environmental samples, but mycobacteriophages have also been isolated from stool samples of tuberculosis patients,[8] although these have yet to be sequenced.[9] About 30 distinct types (called clusters, or singletons if they have no relatives) that share little nucleotide sequence similarity have been identified. Many of the clusters span sufficient diversity that the genomes warrant division into subclusters (Figure 1).[9]
There is also considerable range in overall guanine plus cytosine content (GC%), from 50.3% to 70%, with an average of 64% (M. smegmatis is 67.3%). Thus, phage GC% does not necessarily match that of its host, and the consequent mismatch of codon usage profiles does not appear to be detrimental. Because new mycobacteriophages lacking extensive DNA similarity with the extant collection are still being discovered, and as there are at least seven singletons for which no relatives have been isolated, we clearly have yet to saturate the diversity of this particular population.[9]
The collection of >50,000 genes can be sorted into >3,900 groups (so-called phamilies, i.e. phage protein families) according to their shared amino acid sequences. Most of these phamilies (~75%) do not have homologues outside of the mycobacteriophages and are of unknown function. Genetic studies with mycobacteriophage Giles show that 45% of the genes are nonessential for lytic growth.[10]
Host range
Host range analysis shows that not all mycobacteriophages from M. smegmatis infect other strains and only phages in Cluster K and in certain subclusters of Cluster A efficiently infect M. tuberculosis (Figure 1).[11] However, mutants can be readily isolated from some phages that expand their host range to infect these other strains.[11] However, the molecular basis of host range depends on the behavior and presence of specific genes. This raises the probability of a correlation between gene phamilies and the preferred host.[11]
The realms of mycobacteriophage infection are not understood in its entirety because it involves various mechanisms including receptor availability, restriction-modification, abortive infection, and more. These mechanisms can be mediated through several processes like Clustered Regulatory Interspaced Short Palindromic Repeats (CRISPRs) and the translational apparatus being modified. Phages overcome these constraints by evolving, spontaneous mutation, and diversifying.[11]
Genome architecture
The first sequenced mycobacteriophage genome was that of mycobacteriophage L5 in 1993.[12] In the following years hundreds of additional genomes have been sequenced.[3] Mycobacteriophages have highly mosaic genomes. Their genome sequences show evidence of extensive horizontal genetic transfer, both between phages and between phages and their mycobacterial hosts. Comparisons of these sequences have helped to explain how frequently genetic exchanges of this type may occur in nature, as well as how phages may contribute to bacterial pathogenicity.[13]
A selection of 60 mycobacteriophages were isolated and had their genomes sequenced in 2009. These genome sequences were grouped into clusters by several methods in an effort to determine similarities between the phages and to explore their genetic diversity. More than half of the phage species were originally found in or near Pittsburgh, Pennsylvania, though others were found in other United States locations, India, and Japan. No distinct differences were found in the genomes of mycobacteriophage species from different global origins.[14]
Mycobacteriophage genomes have been found to contain a subset of genes undergoing more rapid genetic flux than other elements of the genomes. These "rapid flux" genes are exchanged between mycobacteriophage more often and are 50 percent shorter in sequence than the average mycobacteriophage gene.[14]
Applications
Historically, mycobacteriophage have been used to "type" (i.e. "diagnose") mycobacteria, as each phage infects only one or a few bacterial strains.[15] In the 1980s phages were discovered as tools to genetically manipulate their hosts.[16] For instance, phage TM4 was used to construct shuttle phasmids that replicate as large cosmids in Escherichia coli and as phages in mycobacteria.[17] Shuttle phasmids can be manipulated in E. coli and used to efficiently introduce foreign DNA into mycobacteria.
Phages with mycobacterial hosts may be especially useful for understanding and fighting mycobacterial infections in humans. A system has been developed to use mycobacteriophage carrying a reporter gene to screen strains of M. tuberculosis for antibiotic resistance.[18] In the future, mycobacteriophage could be used to treat infections by phage therapy.[19][20]
In 2019 it was reported that three mycobacteriophages were administered intravenously twice daily to a 15 year-old girl with cystic fibrosis and disseminated M. abscessus subsp massiliense infection that occurred following lung transplant.[21] The patient had clear benefit from treatment, and the phage treatment combined with antibiotics was extended for several years. In 2022 it was reported that two mycobacteriophages were administered intravenously twice daily to a young man with treatment-refractory Mycobacterium abscessus subsp abscessus pulmonary infection and severe cystic fibrosis lung disease.[22] Airway cultures for M. abscessus became negative after approximately 100 days of combined phage and antibiotic treatment, and a variety of biomarkers confirmed the therapeutic response. The individual received a bilateral lung transplant after 379 days of treatment, and cultures from the explanted lung tissue confirmed eradication of the bacteria.
References
- Padilla-Sanchez V (24 July 2021), Mycobacteriophage ZoeJ Structural Model at Atomic Resolution, doi:10.5281/zenodo.5132914, retrieved 24 July 2021
- Mankiewicz E (September 1961). "Mycobacteriophages isolated from persons with tuberculous and non-tuberculous conditions". Nature. 191 (4796): 1416–1417. Bibcode:1961Natur.191.1416M. doi:10.1038/1911416b0. PMID 14469307. S2CID 4263411.
- "Mycobacteriophage Database". Phagesdb.org. Retrieved 4 September 2017.
- Hatfull GF (13 October 2010). "Mycobacteriophages: genes and genomes". Annual Review of Microbiology. 64 (1): 331–356. doi:10.1146/annurev.micro.112408.134233. PMID 20528690.
- Pope WH, Jacobs-Sera D, Russell DA, Peebles CL, Al-Atrache Z, Alcoser TA, et al. (January 2011). "Expanding the diversity of mycobacteriophages: insights into genome architecture and evolution". PLOS ONE. 6 (1): e16329. Bibcode:2011PLoSO...616329P. doi:10.1371/journal.pone.0016329. PMC 3029335. PMID 21298013.
- Gardner GM, Weiser RS (October 1947). "A bacteriophage for Mycobacterium smegmatis". Proceedings of the Society for Experimental Biology and Medicine. 66 (1): 205–206. doi:10.3181/00379727-66-16037. PMID 20270730. S2CID 20772051.
- Froman S, Will DW, Bogen E (October 1954). "Bacteriophage active against virulent Mycobacterium tuberculosis. I. Isolation and activity". American Journal of Public Health and the Nation's Health. 44 (10): 1326–1333. doi:10.2105/AJPH.44.10.1326. PMC 1620761. PMID 13197609.
- Hatfull GF (March 2014). "Mycobacteriophages: windows into tuberculosis". PLOS Pathogens. 10 (3): e1003953. doi:10.1371/journal.ppat.1003953. PMC 3961340. PMID 24651299.
- Dedrick RM, Marinelli LJ, Newton GL, Pogliano K, Pogliano J, Hatfull GF (May 2013). "Functional requirements for bacteriophage growth: gene essentiality and expression in mycobacteriophage Giles". Molecular Microbiology. 88 (3): 577–589. doi:10.1111/mmi.12210. PMC 3641587. PMID 23560716.
- Jacobs-Sera D, Marinelli LJ, Bowman C, Broussard GW, Guerrero Bustamante C, Boyle MM, et al. (Science Education Alliance Phage Hunters Advancing Genomics And Evolutionary Science Sea-Phages Program) (December 2012). "On the nature of mycobacteriophage diversity and host preference". Virology. 434 (2): 187–201. doi:10.1016/j.virol.2012.09.026. PMC 3518647. PMID 23084079.
- Hatfull GF, Sarkis GJ (February 1993). "DNA sequence, structure and gene expression of mycobacteriophage L5: a phage system for mycobacterial genetics". Molecular Microbiology. 7 (3): 395–405. doi:10.1111/j.1365-2958.1993.tb01131.x. PMID 8459766. S2CID 10188307.
- Pedulla ML, Ford ME, Houtz JM, Karthikeyan T, Wadsworth C, Lewis JA, et al. (April 2003). "Origins of highly mosaic mycobacteriophage genomes". Cell. 113 (2): 171–182. doi:10.1016/S0092-8674(03)00233-2. PMID 12705866. S2CID 14055875.
- Hatfull GF, Jacobs-Sera D, Lawrence JG, Pope WH, Russell DA, Ko CC, et al. (March 2010). "Comparative genomic analysis of 60 Mycobacteriophage genomes: genome clustering, gene acquisition, and gene size". Journal of Molecular Biology. 397 (1): 119–143. doi:10.1016/j.jmb.2010.01.011. PMC 2830324. PMID 20064525.
- Jones WD (1975). "Phage typing report of 125 strains of "Mycobacterium tuberculosis"". Annali Sclavo; Rivista di Microbiologia e di Immunologia. 17 (4): 599–604. PMID 820285.
- Jacobs WR Jr. 2000. Mycobacterium tuberculosis: a once genetically intractable organism. In Molecular Genetics of the Mycobacteria, ed. GF Hatfull, WR Jacobs Jr, pp. 1–16. Washington, DC: ASM Press
- Jacobs WR, Tuckman M, Bloom BR (1987). "Introduction of foreign DNA into mycobacteria using a shuttle phasmid". Nature. 327 (6122): 532–535. Bibcode:1987Natur.327..532J. doi:10.1038/327532a0. PMID 3473289. S2CID 9192407.
- Mulvey MC, Sacksteder KA, Einck L, Nacy CA (2012). "Generation of a novel nucleic acid-based reporter system to detect phenotypic susceptibility to antibiotics in Mycobacterium tuberculosis". mBio. 3 (2): e00312–11. doi:10.1128/mBio.00312-11. PMC 3312217. PMID 22415006.
- Danelishvili L, Young LS, Bermudez LE (2006). "In vivo efficacy of phage therapy for Mycobacterium avium infection as delivered by a nonvirulent mycobacterium". Microbial Drug Resistance. 12 (1): 1–6. doi:10.1089/mdr.2006.12.1. PMID 16584300.
- McNerney R, Traoré H (2005). "Mycobacteriophage and their application to disease control". Journal of Applied Microbiology. 99 (2): 223–233. doi:10.1111/j.1365-2672.2005.02596.x. PMID 16033452. S2CID 43134099.
- Dedrick RM, Guerrero-Bustamante CA, Garlena RA, Russell DA, Ford K, Harris K, et al. (May 2019). "Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus". Nature Medicine. 25 (5): 730–733. doi:10.1038/s41591-019-0437-z. PMC 6557439. PMID 31068712.
- Nick JA, Dedrick RM, Gray AL, Vladar EK, Smith BE, Freeman KG, et al. (May 2022). "Host and pathogen response to bacteriophage engineered against Mycobacterium abscessus lung infection". Cell. 185 (11): 1860–1874.e12. doi:10.1016/j.cell.2022.04.024. PMID 35568033. S2CID 248755782.