Mosasaur
Mosasaurs (from Latin Mosa meaning the 'Meuse', and Greek σαύρος sauros meaning 'lizard') comprise a group of extinct, large marine reptiles from the Late Cretaceous. Their first fossil remains were discovered in a limestone quarry at Maastricht on the Meuse in 1764. They belong to the order Squamata, which includes lizards and snakes.
| Mosasaurs Temporal range: Late Cretaceous, | |
|---|---|
 | |
| Mounted skeleton of a plioplatecarpine (Plesioplatecarpus planifrons), Rocky Mountain Dinosaur Resource Center | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Order: | Squamata |
| Clade: | Pythonomorpha |
| Superfamily: | †Mosasauroidea Gervais, 1853 |
| Subgroups | |
| |
Mosasaurs probably evolved from an extinct group of aquatic lizards[1] known as aigialosaurs in the Earliest Late Cretaceous with 42 described genera. During the last 20 million years of the Cretaceous period (Turonian–Maastrichtian ages), with the extinction of the ichthyosaurs and pliosaurs, mosasaurs became the dominant marine predators. They themselves became extinct as a result of the K-Pg event at the end of the Cretaceous period, about 66 million years ago.
Description

Mosasaurs breathed air, were powerful swimmers, and were well-adapted to living in the warm, shallow inland seas prevalent during the Late Cretaceous period. Mosasaurs were so well adapted to this environment that they most likely gave birth to live young, rather than returning to the shore to lay eggs as sea turtles do.[2]
The smallest-known mosasaur was Dallasaurus turneri, which was less than 1 m (3.3 ft) long. Larger mosasaurs were more typical, with many species growing longer than 4 m (13 ft). Mosasaurus hoffmannii, the largest known species reached up to 17 m (56 ft),[3] but it has been considered to be probably overestimated by Cleary et al. (2018).[4] Currently, the largest publicly exhibited mosasaur skeleton in the world is on display at the Canadian Fossil Discovery Centre in Morden, Manitoba. The specimen, nicknamed "Bruce", is just over 15 m (49 ft) long,[5] but this might be an overestimate as the skeleton was assembled for display prior to a 2010 reassessment of the species that found its original number of vertebrae to be exaggerated, implying that the actual size of the animal was likely smaller.[6][7]
Mosasaurs had a body shape similar to that of modern-day monitor lizards (varanids), but were more elongated and streamlined for swimming. Their limb bones were reduced in length and their paddles were formed by webbing between their long finger and toe bones. Their tails were broad, and supplied their locomotive power.
Until recently, mosasaurs were assumed to have swum in a method similar to the one used today by conger eels and sea snakes, undulating their entire bodies from side to side. However, new evidence suggests that many advanced mosasaurs had large, crescent-shaped flukes on the ends of their tails, similar to those of sharks and some ichthyosaurs. Rather than use snake-like undulations, their bodies probably remained stiff to reduce drag through the water, while their tails provided strong propulsion.[8] These animals may have lurked and pounced rapidly and powerfully on passing prey, rather than chasing after it.[9]
Early reconstructions showed mosasaurs with dorsal crests running the length of their bodies, which were based on misidentified remains of tracheal cartilage. By the time this error was discovered, depicting mosasaurs with such crests in artwork had already become a trend.[10][11]
Paleobiology


Mosasaurs had double-hinged jaws and flexible skulls (much like those of snakes), which enabled them to gulp down their prey almost whole. A skeleton of Tylosaurus proriger from South Dakota included remains of the diving seabird Hesperornis, a marine bony fish, a possible shark, and another, smaller mosasaur (Clidastes). Mosasaur bones have also been found with shark teeth embedded in them.
One of the food items of mosasaurs were ammonites, molluscs with shells similar to those of Nautilus, which were abundant in the Cretaceous seas. Holes have been found in fossil shells of some ammonites, mainly Pachydiscus and Placenticeras. These were once interpreted as a result of limpets attaching themselves to the ammonites, but the triangular shape of the holes, their size, and their presence on both sides of the shells, corresponding to upper and lower jaws, is evidence of the bite of medium-sized mosasaurs. Whether this behaviour was common across all size classes of mosasaurs is not clear.
Virtually all forms were active predators of fish and ammonites; a few, such as Globidens, had blunt, spherical teeth, specialized for crushing mollusk shells. The smaller genera, such as Platecarpus and Dallasaurus, which were about 1–6 m (3.3–19.7 ft) long, probably fed on fish and other small prey. The smaller mosasaurs may have spent some time in fresh water, hunting for food. The largest mosasaur Mosasaurus hoffmannii was the apex predator of the Late Cretaceous oceans, reaching more than 11 metres (36 ft) in length and weighing up to 10 metric tons (11 short tons) in body mass.[12]
Soft tissue

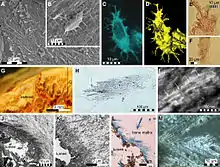
Despite the many mosasaur remains collected worldwide, knowledge of the nature of their skin coverings remains in its early stages. Few mosasaurid specimens collected from around the world retain fossilized scale imprints. This lack may be due to the delicate nature of the scales, which nearly eliminates the possibility of preservation, in addition to the preservation sediment types and the marine conditions under which the preservation occurred. Until the discovery of several mosasaur specimens with remarkably well-preserved scale imprints from late Maastrichtian deposits of the Muwaqqar Chalk Marl Formation of Harrana[13] in Jordan, knowledge of the nature of mosasaur integument was mainly based on very few accounts describing early mosasaur fossils dating back to the upper Santonian–lower Campanian, such as the famous Tylosaurus specimen (KUVP-1075) from Gove County, Kansas.[14]
Material from Jordan has shown that the bodies of mosasaurs, as well as the membranes between their fingers and toes, were covered with small, overlapping, diamond-shaped scales resembling those of snakes. Much like those of modern reptiles, mosasaur scales varied across the body in type and size. In Harrana specimens, two types of scales were observed on a single specimen: keeled scales covering the upper regions of the body and smooth scales covering the lower.[13] As ambush predators, lurking and quickly capturing prey using stealth tactics,[15] they may have benefited from the nonreflective, keeled scales.[13] Additionally, mosasaurs had large pectoral girdles, and such genera as Plotosaurus may have used their front flippers in a breaststroke motion to gain added bursts of speed during an attack on prey.[16]

More recently, a fossil of Platecarpus tympaniticus has been found that preserved not only skin impressions, but also internal organs. Several reddish areas in the fossil may represent the heart, lungs, and kidneys. The trachea is also preserved, along with part of what may be the retina in the eye. The placement of the kidneys is farther forward in the abdomen than it is in monitor lizards, and is more similar to those of cetaceans. As in cetaceans, the bronchi leading to the lungs run parallel to each other instead of splitting apart from one another as in monitors and other terrestrial reptiles. In mosasaurs, these features may be internal adaptations to fully marine lifestyles.[8]
In 2011, collagen protein was recovered from a Prognathodon humerus dated to the Cretaceous.[17]
In 2005, a case study by A.S. Schulp, E.W.A Mulder, and K. Schwenk outlined the fact that mosasaurs had paired fenestrae in their palates. In monitor lizards and snakes, paired fenestrae are associated with a forked tongue, which is flicked in and out to detect chemical traces and provide a directional sense of smell. They therefore proposed that mosasaurs probably also had a sensitive forked tongue.[18]
Metabolism
A study published in 2016 by T. Lyn Harrell, Alberto Pérez-Huerta and Celina Suarez showed that mosasaurs were endothermic. The study contradicted findings published in 2010 indicating mosasaurs were ectothermic. The 2010 study did not use warm-blooded animals for comparison but analogous groups of common marine animals. Based on comparisons with modern warm-blooded animals and fossils of known cold-blooded animals from the same time period, the 2016 study found mosasaurs likely had body temperatures similar to those of contemporary seabirds and were able to internally regulate their temperatures to remain warmer than the surrounding water.[19]
Coloration
The coloration of mosasaurs was unknown until 2014, when the findings of Johan Lindgren of Lund University and colleagues revealed the pigment melanin in the fossilized scales of a mosasaur. Mosasaurs were likely countershaded, with dark backs and light underbellies, much like a great white shark or leatherback sea turtle, the latter of which had fossilized ancestors for which color was also determined. The findings were described in Nature.[20]
Teeth
Mosasaurs possessed a thecodont dentiton, meaning that the roots were cemented deeply into the jaw bone. Mosasaurs did not use permanent teeth but instead constantly shed them. Replacement teeth developed within a pit inside the roots of the original tooth called the resorption pit. This is done through a distinctively unique eight-stage process. The first stage was characterized by the mineralization of a small tooth crown developed elsewhere that descended into the resorption pit by the second stage. In the third stage, the developing crown firmly cemented itself within the resorption pit and grew in size; by the fourth stage, it would be of the same size as the crown in the original tooth. Stages five and six were characterized by the development of the replacement tooth's root: in stage five the root developed vertically, and in stage six the root expanded in all directions to the point that the replacement tooth became exposed and actively pushed on the original tooth. In the seventh stage, the original tooth was shed and the now-independent replacement tooth began to anchor itself into the vacancy. In the eighth and final stage, the replacement tooth has grown to firmly anchor itself.[21]
Ontogeny and growth
Mosasaur growth is not well understood, as specimens of juveniles are rare, and many were mistaken for hesperornithine birds when discovered 100 years ago. However, the discovery of several specimens of juvenile and neonate-sized mosasaurs unearthed more than a century ago indicate that mosasaurs gave birth to live young, and that they spent their early years of life out in the open ocean, not in sheltered nurseries or areas such as shallow water as previously believed. Whether mosasaurs provided parental care, like other marine reptiles such as plesiosaurs, is currently unknown. The discovery of young mosasaurs was published in the journal Palaeontology.[22]
In late 2014 Guinness World Records awarded the museum with a record for Largest Publicly Displayed Mosasaur – Bruce. The record was added to the 2016 print edition of Guinness World Records.[23]
Possible eggs
A 2020 study published in Nature described a large fossilized hatched egg from Antarctica from the very end of the Cretaceous, about 68 million years ago. The egg is considered one of the largest amniote eggs ever known, rivalling that of the elephant bird, and due to its soft, thin, folded texture, it likely belonged to a marine animal. While the organism that produced it remains unknown, the egg's pore structure is very similar to that of extant lepidosaurs such as lizards and snakes, and presence of mosasaur fossils nearby indicates that it may have been a mosasaur egg. It is unknown whether the egg was laid on land or in the water. The egg was assigned to the newly described oospecies Antarcticoolithus bradyi.[24][25][26]
Environment
Paleotologists compared the taxonomic diversity and patterns of morphological disparity in mosasaurs with sea level, sea surface temperature, and stable carbon isotope curves for the Upper Cretaceous to explore factors that may have influenced their evolution. No single factor unambiguously accounts for all radiations, diversification, and extinctions; however, the broader patterns of taxonomic diversification and morphological disparity point to niche differentiation in a "fishing up" scenario under the influence of "bottom-up" selective pressures. The most likely driving force in mosasaur evolution was high productivity in the Late Cretaceous, driven by tectonically controlled sea levels and climatically controlled ocean stratification and nutrient delivery. When productivity collapsed at the end of the Cretaceous, coincident with bolide impact, mosasaurs became extinct.[27]

Sea levels were high during the Cretaceous period, causing marine transgressions in many parts of the world, and a great inland seaway in what is now North America. Mosasaur fossils have been found in the Netherlands, Belgium, Denmark, Portugal, Sweden, South Africa, Spain, France, Germany, Poland, the Czech Republic, Italy[28] Bulgaria, the United Kingdom,[29][30] Russia, Ukraine, Kazakhstan, Azerbaijan,[31] Japan,[32] Egypt, Israel, Jordan, Syria,[33] Turkey,[34] Niger,[35][36] Angola, Morocco, Australia, New Zealand, and on Vega Island off the coast of Antarctica. Tooth taxon Globidens timorensis is known from the island of Timor; however, the phylogenetic placement of this species is uncertain and it might not even be a mosasaur.[37]
Mosasaurs have been found in Canada in Manitoba and Saskatchewan[38] and in much of the contiguous United States. Complete or partial specimens have been found in Alabama, Mississippi, New Jersey, Tennessee, and Georgia, as well as in states covered by the Cretaceous seaway: Texas, southwest Arkansas, New Mexico, Kansas,[39] Colorado, Nebraska, South Dakota, Montana, Wyoming, and the Pierre Shale/Fox Hills formations of North Dakota.[40] Lastly, mosasaur bones and teeth are also known from California, Mexico, Colombia,[41] Brazil,[33] Peru, and Chile.[42]
Many of the so-called 'dinosaur' remains found on New Zealand are actually mosasaurs and plesiosaurs, both being Mesozoic predatory marine reptiles.
Discovery

The first publicized discovery of a partial fossil mosasaur skull in 1764 by quarry workers in a subterranean gallery of a limestone quarry in Mount Saint Peter, near the Dutch city of Maastricht, preceded any major dinosaur fossil discoveries, but remained little known. However, a second find of a partial skull drew the Age of Enlightenment's attention to the existence of fossilized animals that were different from any known living creatures. When the specimen was discovered between 1770 and 1774, Johann Leonard Hoffmann, a surgeon and fossil collector, corresponded about it with the most influential scientists of his day, making the fossil famous. The original owner, though, was Godding, a canon of Maastricht cathedral.
When the French revolutionary forces occupied Maastricht in 1794, the carefully hidden fossil was uncovered, after a reward, it is said, of 600 bottles of wine, and transported to Paris. After it had been earlier interpreted as a fish, a crocodile, and a sperm whale, the first to understand its lizard affinities was the Dutch scientist Adriaan Gilles Camper in 1799. In 1808, Georges Cuvier confirmed this conclusion, although le Grand Animal fossile de Maëstricht was not actually named Mosasaurus ('Meuse reptile') until 1822 and not given its full species name, Mosasaurus hoffmannii, until 1829. Several sets of mosasaur remains, which had been discovered earlier at Maastricht but were not identified as mosasaurs until the 19th century, have been on display in the Teylers Museum, Haarlem, procured from 1790.
The Maastricht limestone beds were rendered so famous by the mosasaur discovery, they have given their name to the final six-million-year epoch of the Cretaceous, the Maastrichtian.
Classification
Relation with snakes or monitor lizards
.jpg.webp)
--Ringelnatter.jpg.webp)
The specific placement of mosasaurs within the Squamata has been controversial since its inception. Cuvier was the first scientist to deeply analyze their possible taxonomic placement through Mosasaurus. While his original 1808 hypothesis that the genus was a lizard with affinities to monitor lizards remained the most popular, Cuvier was uncertain, even at the time, about the accuracy of this placement. He simultaneously proposed a number of alternative hypotheses, with one such alternative suggesting that Mosasaurus instead had closer affinities with iguanas due to their shared presence of pterygoid teeth. With the absence of sufficient fossil evidence, researchers during the early and mid-19th century had little to work with. Instead, they primarily relied on stratigraphic associations and Cuvier's 1808 research on the holotype skull. Thus, in-depth research on the placement of Mosasaurus was not undertaken until the discovery of more complete mosasaur fossils during the late 19th century, which reignited research on the placement of mosasaurs among squamates.[43]
In a span of about 30 to 40 years during the late 19th to early 20th centuries, paleontologists fiercely debated the issue, which created two major schools of thought: one that supported a monitor lizard relationship and one that supported a closer relationship with snakes.[43] The proposition of a snake relationship was spearheaded by Cope, who first published such a hypothesis in 1869 by proposing that mosasaurs, which he classified under a group called the Pythonomorpha, was the sister group of snakes. Some scientists went as far as to interpret mosasaurs as direct ancestors of snakes.[44] Opponents supporting a relationship with monitor lizards argued that mosasaurs should be placed within the infraorder Anguimorpha, with lower classifications varying from placing mosasaurs within Varanoidea or its sister taxa, or as true monitor lizards within Varanidae.[43] These debates spawned higher taxonomic groups that were erected in attempts to classify the placement of mosasaurs (although not all are compatible). One of these was the Mosasauria, a loosely-defined group erected by Marsh in 1880 but is still used by some researchers today that includes all descendants of the last common ancestor of Mosasaurus and some of its ancestral relatives, including the marine squamates dolichosaurs Dolichosaurus and Coniasaurus and Adriosaurus.[45]
In 1923, Charles Lewis Camp published Classification of the Lizards, in which he proposed through the review and rebuttal of previous arguments using his own anatomical observations that all taxa more closely related to Mosasaurus than Dolichosaurus should be classified into a superfamily called the Mosasauroidea, which would be a sister superfamily to the Varanoidea.[43][46] Camp's take on the subject virtually ended the snake-monitor lizard debate for approximately 70 years, with nearly all subsequent studies supporting a relationship with monitor lizards. However, many studies continued to support going further than Camp in the monitor lizard relationship, placing mosasaurs within the Varanoidea.[43]
The debate was reignited with the publication of a 1997 cladistical study by Michael S. Y. Lee, which recovered the Mosasauroidea as a sister taxon to the snake suborder Serpentes and resurrected the argument for a snake relationship.[43][47] Prior, there had never been a modern phylogenetic study specifically testing the relationships between mosasaurs or snakes. Lee also resurrected the defunct Pythonomorpha and redefined it to unify the Mosasauroidea and Serpentes under one clade.[47] Multiple subsequent studies conducted by scientists such as Lee, Caldwell, and Alessandro Palci refined this hypothesis. In some studies, the Mosasauria clade was used to represent mosasaurs.[43][44] However, there still remained little consensus. For example, a large-scale 2008 phylogenetic study by paleontologist Jack Conrad of the American Museum of Natural History recovered the Mosasauria clade in a polytomy, or a clade with unresolved relationships, with monitor lizards and beaded lizards;[45] and a 2012 study by Gauthier et al. recovered Mosasauria as a clade basal to both monitor lizards and snakes.[48]
With the advent of molecular genetics during the 2010s, some scientists argued for the combining of molecular and morphological data to examine relationships between mosasaurs and living squamates.[49][50] An early study was Lee (2009), which based on nuclear and mitochondrial DNA in living squamates and morphological data recovered mosasaurs as a stem group to snakes,[51] which some later authors interpreted as placement of snakes within the Mosasauria clade itself.[49] However, a 2010 study by Weins et al. attempted to replicate Lee (2009) using a larger dataset but instead yielded results that recovered the Mosasauria as a sister clade to the monitor lizards.[49] The frequent discrepancies are due to the high prevalence of convergent evolution in squamates, which creates much room for interpreting molecular and morphological data; many of these studies had results that often contradicted each other, for example in completely different phylogenetic results by simply adding more datasets or varying which taxa are represented, which resulted in various conflicts creating even more uncertainty. Due to this, some scientists argued that a molecular perspective should be abandoned entirely.[50][52] Nevertheless, other scientists have attempted to resolve these problems. One approach was utilized by a 2015 study by Reeder et al.: it closely integrated morphological, molecular, and paleontological data in a large dataset to overcome previous conflicts, which revealed new morphological support for molecular results that recovered Mosasauria as a sister clade to Serpentes.[50] Another approach was developed by R. Alexander Pyron in a 2016 study, which also recovered Mosasauria as a sister clade to the Serpentes.[53]
Lower classifications




The traditional view of mosasaur evolution held that all paddle-limbed (hydropedal) mosasaurs originated from a single common ancestor with functional legs (plesiopedal). However, this was shaken with the discovery of Dallasaurus, a plesiopedal mosasauroid that was found by multiple phylogenetic analyses to be an ancestor of the Mosasaurinae but unrelated to other mosasaurs. Bell and Polycn (2005) grouped these outside mosasaurs into two clades: the Russellosaurina, whose basal members include plesiopedal genera (Tethysaurinae) of their own and derived members consisting of the Plioplatecarpinae and Tylosaurinae; and the Halisauromorpha, containing the Halisaurinae. The placement of Dallasaurus suggested that the Russellosaurina and Halisauromorpha may have evolved a hydropedal form independently, the former through the tethysaurines, meaning that their placement within the Mosasauridae creates an unnatural polyphyly and thus potentially invalid.[54][43] Caldwell informally proposed in a 2012 publication that the definition of a mosasaur must thus be redefined into one that does not consider russellosaurines and halisauromorphs as true mosasaurs, but as an independent group of marine lizards.[43]
However, phylogenetic studies of mosasaurs can be fickle, especially when wild card taxa like Dallasaurus remain poorly understood. For example, some studies such as a 2009 analysis by Dutchak and Caldwell instead found that Dallasaurus was ancestral to both russellosaurines and mosasaurines,[55] although results were inconsistent in later studies.[56] A 2017 study by Simoes et al. noted that utilization of different methods of phylogenetic analyses can yield different findings and ultimately found a consistent indication that tethysaurines were a case of hydropedal mosasaurs reversing back to a plesiopedal condition rather than an independent ancestral feature.[56]
Phylogeny
Cladogram of the Mosasauroidea from Madzia and Cau (2017):[57]
| Mosasauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distribution
Though no individual genus or subfamily is found worldwide, the Mosasauridae as a whole achieved global distribution during the Late Cretaceous with many locations typically having complex mosasaur faunas with multiple different genera and species in different ecological niches.
Two African countries are particularly rich in mosasaurs: Morocco[58] and Angola.[59][60]
In contemporary culture
Mosasaurs, along with other extinct aquatic Mesozoic reptiles, are often erroneously depicted as 'swimming dinosaurs' in popular culture. The belief that mosasaurs are dinosaurs is a common misconception.[61][62]
References
- Dash, Sean (2008). Prehistoric Monsters Revealed. United States: Workaholic Productions / History Channel. Archived from the original on 2016-01-27. Retrieved December 18, 2015.
- Field, Daniel J.; LeBlanc, Aaron; Gau1, Adrienne; Behlke, Adam D. (10 April 2015). "Pelagic neonatal fossils support viviparity and precocial life history of Cretaceous mosasaurs". Palaeontology. 58 (3): 401–407. doi:10.1111/pala.12165. S2CID 4660322.
- Grigoriev, D.W. (2014). "Giant Mosasaurus hoffmanni (Squamata, Mosasauridae) from the Late Cretaceous (Maastrichtian) of Penza, Russia" (PDF). Proceedings of the Zoological Institute RAS. 318 (2): 148–167. doi:10.31610/trudyzin/2014.318.2.148. Retrieved 26 June 2016.
- Terri J. Cleary; Roger B. J. Benson; Susan E. Evans; Paul M. Barrett (2018). "Lepidosaurian diversity in the Mesozoic–Palaeogene: the potential roles of sampling biases and environmental drivers". Royal Society Open Science. 5: 171830. doi:10.1098/rsos.171830.
- "Largest mosasaur on display". Guinness World Records. 2014. Retrieved 27 June 2016.
- CBC News (August 27, 2008). "Manitoba dig uncovers 80-million-year-old sea creature". CBC. Manitoba. Archived from the original on June 5, 2018.
- Bullard, T.S.; Caldwell, M.W. (2010). "Redescription and rediagnosis of the tylosaurine mosasaur Hainosaurus pembinensis Nicholls, 1988, as Tylosaurus pembinensis (Nicholls, 1988)". Journal of Vertebrate Paleontology. 30 (2): 416–426. doi:10.1080/02724631003621870. S2CID 86297189.
- Lindgren, J.; Caldwell, M.W.; Konishi, T.; Chiappe, L.M. (2010). Farke, Andrew Allen (ed.). "Convergent Evolution in Aquatic Tetrapods: Insights from an Exceptional Fossil Mosasaur". PLOS ONE. 5 (8): e11998. Bibcode:2010PLoSO...511998L. doi:10.1371/journal.pone.0011998. PMC 2918493. PMID 20711249.
- Lindgren, J.; Kaddumi, H. F.; Polcyn, M. J. (2013). "Soft tissue preservation in a fossil marine lizard with a bilobed tail fin". Nature Communications. 4: 2423. Bibcode:2013NatCo...4.2423L. doi:10.1038/ncomms3423. PMID 24022259.
- Osborn, Henry Fairfield (1899). "A Complete Mosasaur Skeleton, Osseous and Cartilaginous". Memoirs of the American Museum of Natural History. 1 (4): 167–188. Bibcode:1899Sci....10..919O. doi:10.1126/science.10.260.919. hdl:2027/mdp.39015042532336. PMID 17837338. Retrieved 25 November 2014.
- Everhart, Mike (13 January 2013). "Origin of the Dorsal Fringe on Mosasaurs". Oceans of Kansas. Retrieved 25 November 2014.
- Fedrico Fanti; Andrea Cau; Alessandra Negri (2014). "A giant mosasaur (Reptilia, Squamata) with an unusually twisted dentition from the Argille Scagliose Complex (late Campanian) of Northern Italy" (PDF). Cretaceous Research. 49 (2014): 91–104. doi:10.1016/j.cretres.2014.01.003.
- Kaddumi, H.F. (2009). "On the latest scale coverings of mosasaurs (Squamata: Mosasauridae) from the Harrana Fauna in addition to the description of s new species of Mosasaurus". Fossils of the Harrana Fauna and the Adjacent Areas. Amman: Eternal River Museum of Natural History. pp. 80–94.
- Snow, F. H. (1878). "On the dermal covering of a mosasauroid reptile". Transactions of the Kansas Academy of Science. 6: 54–58. doi:10.2307/3623557. JSTOR 3623557.
- Massare, J. A. (1987). "Tooth morphology and prey preference of Mesozoic marine reptiles". Journal of Vertebrate Paleontology. 7 (2): 121–137. doi:10.1080/02724634.1987.10011647.
- "Did mosasaurs do the breast stroke?".
- Lindgren, Johan; Uvdal, Per; Engdahl, Anders; Lee, Andrew H.; Alwmark, Carl; Bergquist, Karl-Erik; Nilsson, Einar; Ekström, Peter; Rasmussen, Magnus; Douglas, Desirée A.; Polcyn, Michael J.; Jacobs, Louis L. (29 April 2011). "Microspectroscopic Evidence of Cretaceous Bone Proteins". PLoS ONE. 6 (4): e19445. Bibcode:2011PLoSO...619445L. doi:10.1371/journal.pone.0019445. ISSN 1932-6203. PMC 3084868. PMID 21559386.
- Schulp, A. S.; Mulder, E. W. A.; Schwenk, K. (2005-09-01). "Did mosasaurs have forked tongues?". Netherlands Journal of Geosciences. 84 (3): 359–371. doi:10.1017/S0016774600021144.
- Harrell, T. Lynn; Pérez-Huerta, Alberto; Suarez, Celina A.; Benson, Roger (May 2016). "Endothermic mosasaurs? Possible thermoregulation of Late Cretaceous mosasaurs (Reptilia, Squamata) indicated by stable oxygen isotopes in fossil bioapatite in comparison with coeval marine fish and pelagic seabirds". Palaeontology. 59 (3): 351–363. doi:10.1111/pala.12240. S2CID 130190966.
- "Scientists cite evidence that mosasaurs were warm-blooded". ScienceDaily (Press release). May 6, 2016.
- Lindgren, J.; Sjövall, P.; Carney, R. M.; Uvdal, P.; Gren, J. A.; Dyke, G.; Schultz, B. P.; Shawkey, M. D.; Barnes, K. R.; Polcyn, M. J. (2014). "Skin pigmentation provides evidence of convergent melanism in extinct marine reptiles". Nature. 506 (7489): 484–8. Bibcode:2014Natur.506..484L. doi:10.1038/nature12899. PMID 24402224. S2CID 4468035.
- Michael W. Caldwell (2007). "Ontogeny, anatomy and attachment of the dentition in mosasaurs (Mosasauridae: Squamata)". Zoological Journal of the Linnean Society. 149 (4): 687–700. doi:10.1111/j.1096-3642.2007.00280.x.
- "What life was like for newborn giant sea lizards during the age of the dinosaur". ScienceDaily. Retrieved 2017-08-01.
- "Largest mosasaur on display". Guinness World Records. Retrieved 2020-06-03.
- "Mystery egg likely 'belonged to giant sea reptile'". BBC News. 2020-06-17. Retrieved 2020-06-18.
- Joel, Lucas (2020-06-17). "Life Hatched From Soft Eggs, Some a Foot Long, in Dinosaur Era". The New York Times. ISSN 0362-4331. Retrieved 2020-06-18.
- Legendre, Lucas J.; Rubilar-Rogers, David; Musser, Grace M.; Davis, Sarah N.; Otero, Rodrigo A.; Vargas, Alexander O.; Clarke, Julia A. (2020-06-17). "A giant soft-shelled egg from the Late Cretaceous of Antarctica". Nature. 583 (7816): 411–414. Bibcode:2020Natur.583..411L. doi:10.1038/s41586-020-2377-7. ISSN 1476-4687. PMID 32555453.
- Polcyn, M. J.; Jacobs, L. L.; Araujo, R.; Schulp, A. S.; Mateus, O. (2014). "Physical drivers of mosasaur evolution". Palaeogeography, Palaeoclimatology, Palaeoecology. 400: 17–27. Bibcode:2014PPP...400...17P. doi:10.1016/j.palaeo.2013.05.018.
- "Druhohorní plazi v Čechách II". DinosaurusBlog. 2015-07-13. Retrieved 2017-08-01.
- "St. James' Pit, Norwich (SSSI)" (PDF). Natural England. 2014. Retrieved 25 November 2014.
- Jagt, John W. M.; Motchurova-Dekova, Neda; Ivanov, Plamen; Cappetta, Henri; Schulp, Anne S. (2006). "Latest Cretaceous mosasaurs and lamniform sharks from Labirinta cave, Vratsa District (northwest Bulgaria): A preliminary note". Geološki Anali Balkanskoga Poluostrva. 67 (67): 51–63. doi:10.2298/gabp0667051j.
- Storrs, Glenn W.; Arkhangelskii, Maxim S.; Efimov, Vladimir M. (2000). "Mesozoic marine reptiles of Russia and other former Soviet republics". In Benton, M. J.; Shishkin, M. A.; Unwin, D. M. (eds.). The age of dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press. pp. 187–210. ISBN 978-0521554763.
- Konishi, Takuya; Tanimoto, Masahiro; Utsunomiya, Satoshi; Sato, Masahiro; Watanabe, Katsunori (2012). "A Large Mosasaurine (Squamata: Mosasauridae) from the Latest Cretaceous of Osaka Prefecture (Sw Japan)". Paleontological Research. 16 (2): 79–87. doi:10.2517/1342-8144-16.2.079. S2CID 130109440.
- Bardet, Nathalie; Pereda Suberbiola, Xabier; Iarochène, Mohamed; Amalik, Mohamed; Bouya, Baadi (Sep 2005). "Durophagous Mosasauridae (Squamata) from the Upper Cretaceous phosphates of Morocco, with description of a new species of Globidens". Netherlands Journal of Geosciences. 84 (3): 167–175. doi:10.1017/S0016774600020953.
- Bardet, Nathalie; Tunoğlu, Cemal (19 September 2002) [24 Aug 2010]. "The first mosasaur (Squamata) from the Late Cretaceous of Turkey". Journal of Vertebrate Paleontology. 22 (3): 712–715. doi:10.1671/0272-4634(2002)022[0712:TFMSFT]2.0.CO;2. ISSN 0272-4634.
- Lingham-Soliar, Theagarten (1991). "Mosasaurs from the upper Cretaceous of Niger". Palaeontology. 34 (3): 653–670 – via BioStor.
- Lingham-Soliar, Theagarten (1998). "A new mosasaur Pluridens walkeri from the Upper Cretaceous, Maastrichtian of the Iullemmeden Basin, southwest Niger". Journal of Vertebrate Paleontology. 18 (4): 709–717. doi:10.1080/02724634.1998.10011100.
- Martin, James E. (2007). "A new species of the durophagous mosasaur, Globidens (Squamata: Mosasauridae) from the Late Cretaceous Pierre Shale Group of central South Dakota, USA". In Martin, James E.; Parris, David C. (eds.). The Geology and Paleontology of the Late Cretaceous Marine Deposits of the Dakotas. Vol. 427. The Geological Society of America. pp. 177–198. doi:10.1130/2007.2427(13). ISBN 978-0-8137-2427-0.
- "General Information". Canadian Fossil Discovery Centre. 2014. Retrieved 25 November 2014.
- Michael J. Everhart (2005). "Chapter 9: Enter the Mosasaurs". Oceans of Kansas: a natural history of the western interior sea. Bloomington: Indiana University Press. ISBN 978-0-253-34547-9.
- Getman, Myron (1994). Occurrences of Mosasaur and other reptilian fossil remains from the Fox Hills Formation (Maastrichtian: late Cretaceous) of North Dakota (Geology Honors thesis). St. Lawrence University Dept. of Geology.
- Páramo-Fonseca, María Eurídice (1 March 2012). "Mosasauroids from Colombia". Bulletin de la Société Géologique de France. 183 (2): 103–109. doi:10.2113/gssgfbull.183.2.103. ISSN 0037-9409 – via GeoScienceWorld.
- Otero, Rodrigo A.; Parham, James F.; Soto-Acuña, Sergio; Jimenez-Huidobro, Paulina; Rubilar-Rogers, David (2012). "Marine reptiles from Late Cretaceous (early Maastrichtian) deposits in Algarrobo, central Chile". Cretaceous Research. 35: 124–132. doi:10.1016/j.cretres.2011.12.003.
- Caldwell, M.W. (2012). "A challenge to categories: "What, if anything, is a mosasaur?"". Bulletin de la Société Géologique de France. 183 (1): 17–34. doi:10.2113/gssgfbull.183.1.7.
- Alessandro Palci. On the Origin and Evolution of the Ophidia (PDF) (PhD). University of Alberta. doi:10.7939/R3NG4H314.
- Jack L. Conrad (2008). "Phylogeny And Systematics Of Squamata (Reptilia) Based On Morphology". Bulletin of the American Museum of Natural History. 310: 1–182. doi:10.1206/310.1. S2CID 85271610.
- Charles Lewis Camp (1923). "Classification of the Lizards". Bulletin of the American Museum of Natural History. 48 (11): 289–481. hdl:2246/898.
- Michael S. Y. Lee (1997). "The Phylogeny of Varanoid Lizards and the Affinities of Snakes". Philosophical Transactions: Biological Sciences. 352 (1349): 53–91. Bibcode:1997RSPTB.352...53L. doi:10.1098/rstb.1997.0005. JSTOR 56451. PMC 1691912.
- Jacques A. Gauthier; Maureen Kearney; Jessica Anderson Maisano; Olivier Rieppel; Adam D.B. Behlke (2012). "Assembling the Squamate Tree of Life: Perspectives from the Phenotype and the Fossil Record". Bulletin of the Peabody Museum of Natural History. 53 (1): 3–308. doi:10.3374/014.053.0101. S2CID 86355757.
- John J. Wiens; Caitlin A. Kuczynski; Ted Townsend; Tod W. Reeder; Daniel G. Mulcahy; Jack W. Sites Jr. (2012). "Combining Phylogenomics and Fossils in Higher-Level Squamate Reptile Phylogeny: Molecular Data Change the Placement of Fossil Taxa" (PDF). Systematic Biology. 59 (6): 674–88. doi:10.1093/sysbio/syq048. PMID 20930035.
- Tod W. Reeder; Ted M. Townsend; Daniel G. Mulcahy; Brice P. Noonan; Perry L. Wood, Jr.; Jack W. Sites, Jr.; John J. Wiens (2015). "Integrated Analyses Resolve Conflicts over Squamate Reptile Phylogeny and Reveal Unexpected Placements for Fossil Taxa". PLOS ONE. 10 (3): e0118199. Bibcode:2015PLoSO..1018199R. doi:10.1371/journal.pone.0118199. PMC 4372529. PMID 25803280.
- Michael S. Y. Lee (2009). "Hidden support from unpromising data sets strongly unites snakes with anguimorph 'lizards'". Journal of Evolutionary Biology. 22 (6): 1308–1316. doi:10.1111/j.1420-9101.2009.01751.x. PMID 19490385. S2CID 43087460.
- N. B. Ananjeva (2019). "Current State of the Problems in the Phylogeny of Squamate Reptiles (Squamata, Reptilia)". Biology Bulletin Reviews. 9 (2): 119–128. doi:10.1134/s2079086419020026. S2CID 162184418.
- A. Alexander Pyron (2016). "Novel Approaches for Phylogenetic Inference from Morphological Data and Total-Evidence Dating in Squamate Reptiles (Lizards, Snakes, and Amphisbaenians)" (PDF). Systematic Biology. 66 (1): 38–56. doi:10.1093/sysbio/syw068. PMID 28173602. S2CID 3697004.
- Bell, G.L. Jr.; Polcyn, M.J. (2005). "Dallasaurus turneri, a new primitive mosasauroid from the Middle Turonian of Texas and comments on the phylogeny of the Mosasauridae (Squamata)". Netherlands Journal of Geoscience. 84 (3): 177–194. doi:10.1017/S0016774600020965.
- Dutchak, A.R.; Caldwell, M.W. (2009). "A redescription of Aigialosaurus (=Opetiosaurus) bucchichi (Kornhuber, 1901) (Squamata: Aigialosauridae) with comments on mosasauroid systematics". Journal of Vertebrate Paleontology. 29 (2): 437–452. doi:10.1671/039.029.0206. S2CID 86140123.
- Simoes, T.R.; Vernygora, O.; Paparella, I.; Jimenez-Huidobro, P.; Caldwell, M.W. (2017). "Mosasauroid phylogeny under multiple phylogenetic methods provides new insights on the evolution of aquatic adaptations in the group". PLOS ONE. 12 (5): e0176773. Bibcode:2017PLoSO..1276773S. doi:10.1371/journal.pone.0176773. PMC 5415187. PMID 28467456.
- Madzia, Daniel; Cau, Andrea (2017-09-15). "Inferring 'weak spots' in phylogenetic trees: application to mosasauroid nomenclature". PeerJ. 5: e3782. doi:10.7717/peerj.3782. ISSN 2167-8359. PMC 5602675. PMID 28929018.
- Bardet, Nathalie; Pereda Suberbiola, Xabier; Iarochene, Mohamed; Bouyahyaoui, Fatima; Bouya, Baadi; Amaghzaz, Mbarek (May 2004). "Mosasaurus beaugei Arambourg, 1952 (Squamata, Mosasauridae) from the Late Cretaceous phosphates of Morocco". Geobios. 37 (3): 315–324. doi:10.1016/j.geobios.2003.02.006.
- Polcyn, Michael J.; Jacobs, Louis L.; Schulp, Anne S.; Mateus, Octávio (March 2010). "The North African Mosasaur Globidens phosphaticus from the Maastrichtian of Angola". Historical Biology. 22 (1–3): 175–185. doi:10.1080/08912961003754978. S2CID 62882332.
- Mateus, Octávio; Callapez, Pedro M.; Polcyn, Michael J.; Schulp, Anne S.; Gonçalves, António Olímpio; Jacobs, Louis L. (2019). "The Fossil Record of Biodiversity in Angola Through Time: A Paleontological Perspective". Biodiversity of Angola: Science & Conservation: A Modern Synthesis. Springer International Publishing. pp. 53–76. doi:10.1007/978-3-030-03083-4_4. ISBN 978-3-030-03082-7. S2CID 133717540.
- "Famous Prehistoric Animals That Weren't Actually Dinosaurs". Feb 17, 2021.
- "The real sea monsters". Science News for Students. 2015-06-19. Retrieved 2022-02-12.
External links
- Palaeos: Vertebrates: Mosasaurs
- BBC Science and Nature: Mosasaurs
- Mike Everhart and David Lewis, "Mesozoic marine monsters of the Mangahouanga": New Zealand fossil fauna
- Mike Everhart, "A day in the life of a Mosasaur": life in the Sea of Kansas, illus. by Carl Buell
- Mike Everhart, "Mosasaurus hoffmani" until 1829.
- Mosasaurus maximus mounted skeleton at University of Texas Memorial Museum
- Canadian Fossil Discovery Centre
- "The Mosasaur of Maastricht" by Hennie Reuvers in Crossroads web magazine
- "Mosasaurs terrorized Cretaceous rivers" Planet Earth online
- Georgia Southern University Museum Mosasaur Exhibit
- Kansas Geological Survey Vol IV (1899), containing the famous summary of American mosasaurs by Samuel Williston.
- William R. Wahl * MOSASAUR BITE MARKS ON AN AMMONITE. PRESERVATION OF AN ABORTED ATTACK?
- Mosasaur diet
| Mosasaurinae |
|  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Halisaurinae |
| ||||||||||
| Russellosaurina |
| ||||||||||
| Related groups and genera |
| ||||||||||
| Related articles |
| ||||||||||