Domestication of the dog
The domestication of the dog was the process which created the domestic dog. This included the dog's genetic divergence from the wolf, its domestication, and the emergence of the first dogs. Genetic studies show that all ancient and modern dogs share a common ancestry and descended from an ancient, now-extinct wolf population – or closely related wolf populations – which was distinct from the modern wolf lineage.[3][4] The dog's similarity to the grey wolf is the result of substantial dog-into-wolf gene flow,[3] with the modern grey wolf being the dog's nearest living relative.[5] An extinct Late Pleistocene wolf may have been the ancestor of the dog.[5][1][6]

The dog is a member of the wolf-like canids.[7][8][9] The genetic divergence between the dog's ancestor and modern wolves occurred between 40,000 and 30,000 years ago, just before or during the Last Glacial Maximum[2][1] (20,000–27,000 years ago). This timespan represents the upper time-limit for the commencement of domestication because it is the time of divergence but not the time of domestication, which occurred later.[2][10]
One of the most important transitions in human history was the domestication of animals, which began with the long-term association between wolves and hunter–gatherers more than 30,000 years ago.[4] The dog was the first species and the only large carnivore to have been domesticated.[11][5] The domestication of the dog occurred due to variation among the common ancestor wolf population in the fight-or-flight response where the common ancestor wolves with less aggression and aversion but greater altruism towards humans received fitness benefits (and vice versa for humans), and thus the domestication of the dog is a prominent example of social selection (rather than artificial selection).[12] The archaeological record and genetic analysis show the remains of the Bonn-Oberkassel dog buried beside humans 14,200 years ago to be the first undisputed dog, with disputed remains occurring 36,000 years ago.
The domestication of the dog predates agriculture,[1] and it was not until 11,000 years ago in the Holocene era that people living in the Near East entered into relationships with wild populations of aurochs, boar, sheep, and goats.[2] Where the domestication of the dog took place remains debated; however, literature reviews of the evidence find that the dog was domesticated in Eurasia,[13] with the most plausible proposals being Central Asia, East Asia, and Western Europe.[2][10] By the close of the most recent Ice Age 11,700 years ago, five ancestral lineages had diversified from each other and were represented through ancient dog samples found in the Levant (7,000 years before present YBP), Karelia (10,900 YBP), Lake Baikal (7,000 YBP), ancient America (4,000 YBP), and in the New Guinea singing dog (present day).[3]
In 2021, a literature review of the current evidence infers that the dog was domesticated in Siberia 23,000 years ago by Ancient North Eurasians, then later dispersed eastwards into the Americas and westwards across Eurasia. Ancient dog remains dating to this time and place have yet to be discovered to support this hypothesis.[13]
The oldest known dog skeletons are found in the Altai Mountains of Siberia and a cave in Belgium, dated ~ 33 000 years ago. According to studies, this may indicate that the domestication of dogs occurred simultaneously in different geographic locations.[14]
Canid and human evolution
6 million years ago, towards the close of the Miocene era, the earth's climate gradually cooled. This would lead to the glaciations of the Pliocene and the Pleistocene, which are commonly referred to as the Ice Age. In many areas, forests and savannahs were replaced with steppes or grasslands, and only those species of creature that adapted to these changes would survive.[15]
Divergence from wolves
Genetic studies indicate that the grey wolf is the closest living relative of the dog.[5][16] Attempting to reconstruct the dog's lineage through the phylogenetic analysis of DNA sequences from modern dogs and wolves has given conflicting results for several reasons. Firstly, studies indicate that an extinct Late Pleistocene wolf is the nearest common ancestor to the dog, with modern wolves not being the dog's direct ancestor. Secondly, the genetic divergence (split) between the dog's ancestor and modern wolves occurred over a short period of time, so that the time of the divergence is difficult to date (referred to as incomplete lineage sorting). This is complicated further by the cross-breeding that has occurred between dogs and wolves since domestication (referred to as post-domestication gene flow). Finally, there have been only tens of thousands of generations of dogs since domestication, so that the number of mutations between the dog and the wolf are few and this makes the timing of domestication difficult to date.[5]
Pleistocene wolves

The Late Pleistocene era was a time of glaciation, climate change, and the advance of humans into isolated areas.[17] During the Late Pleistocene glaciation, a vast mammoth steppe stretched from Spain eastwards across Eurasia and over Beringia into Alaska and the Yukon. The close of this era was characterized by a series of severe and rapid climate oscillations with regional temperature changes of up to 16 °C (29 °F), which has been correlated with megafaunal extinctions. There is no evidence of megafaunal extinctions at the height of the Last Glacial Maximum (26,500 YBP), indicating that increasing cold and glaciation were not factors. Multiple events appear to have caused the rapid replacement of one species by another one within the same genus, or one population by another within the same species, across a broad area. As some species became extinct, so too did the predators that depended on them (coextinction).[18]
The grey wolf is one of the few large carnivores to survive the Late Pleistocene megafaunal extinctions, but similar to many other megafaunal species it experienced a global population decline towards the end of this era, which was associated with extinctions of ecomorphs and phylogeographic shifts in populations.[19] Grey wolf mitochondrial genomes (excluding the Himalayan wolf and the Indian plains wolf) indicate that the most recent common ancestor for all C. lupus specimens studied – modern and extinct – dates to 80,000 YBP, and this is more recent than the time suggested by the fossil record.[20][21] The fossil record suggests that the earliest grey wolf specimens were found in what was once eastern Beringia at Old Crow, Yukon, in Canada and at Cripple Creek Sump, Fairbanks, in Alaska. The age is not agreed but could date 1 million YBP.[1] All modern wolves (excluding the Himalayan wolf and the Indian plains wolf) show a most recent common ancestor dating to 32,000 YBP, which coincides with the commencement of their global demographic decline.[19]
The origin of dogs is couched in the biogeography of wolf populations that lived during the Late Pleistocene.[1] The fossil record shows evidence of changes in the morphology and body size of wolves during the Late Pleistocene, which may be due to differences in their prey size. Wolf skeletal development can be changed due to a preference for larger prey which results in larger wolves.[17] Considerable morphological diversity existed among grey wolves by the Late Pleistocene. These are regarded as having been more cranio-dentally robust than modern grey wolves, often with a shortened rostrum, the pronounced development of the temporalis muscle, and robust premolars. It is proposed that these features were specialized adaptations for the processing of carcass and bone associated with the hunting and scavenging of Pleistocene megafauna. Compared with modern wolves, some Pleistocene wolves showed an increase in tooth breakage that is similar to that seen in the extinct dire wolf. This suggests that these either often processed carcasses, or that they competed with other carnivores and needed to quickly consume their prey. The frequency and location of tooth fractures found in these wolves compared with the modern spotted hyena indicates that these wolves were habitual bone crackers.[1] These ancient wolves carried mitochondrial lineages which cannot be found among modern wolves, which implies their extinction.[19]
Grey wolves suffered a species-wide population bottleneck (reduction) approximately 25,000 YBP during the Last Glacial Maximum. This was followed by a single population of modern wolves expanding out of a Beringia refuge to repopulate the wolf's former range, replacing the remaining Late Pleistocene wolf populations across Eurasia and North America as they did so.[22][23][17] This source population probably did not give rise to dogs, but it admixed with dogs which allowed them to gain coat colour genes that are also related to immunity. There is little genetic information available on the ancient wolves that existed prior to the bottleneck. However, studies show that one or more of these ancient populations is more directly ancestral to dogs than are modern wolves, and conceivably these were more prone to domestication by the first humans to expand into Eurasia.[17]
An apex predator sits on the top trophic level of the food chain, while a mesopredator sits further down the food chain and is dependent on smaller animals. Towards the end of the Pleistocene era, most of today's apex predators were mesopredators and this included the wolf. During the ecological upheaval associated with the close of the Late Pleistocene, one type of wolf population rose to become today's apex predator and another joined with humans to become an apex consumer.[24] The domestication of this lineage ensured its evolutionary success through its expansion into a new ecological niche.[19]
For a long time scientists assumed that dogs evolved from the modern grey wolf. But a study published in 2014 concluded that this was incorrect, and that dogs are descended from an extinct type of wolf.[25][26]
It was such a long standing view that the gray wolf that we know today was around for hundreds of thousands of years and that dogs derived from them. We're very surprised that they're not.
— Robert K. Wayne[25]
Time of genetic divergence
The date estimated for the divergence of a domestic lineage from a wild one does not necessarily indicate the start of the domestication process but it does provide an upper boundary. The divergence of the lineage that led to the domestic horse from the lineage that led to the modern Przewalski's horse is estimated to have occurred around 45,000 YBP but the archaeological record indicates 5,500 YBP. The variance can be due to modern wild populations not being the direct ancestor of the domestic ones, or to a divergence caused by changes in the climate, topography, or other environmental influences.[10] Recent studies indicate that a genetic divergence occurred between the dog's and modern wolves 20,000–40,000 YBP; however, this is the upper time-limit for domestication because it represents the time of divergence and not the time of domestication.[10][2]
In 2013, the mitochondrial DNA (mDNA) sequencing of ancient wolves together with whole genome sequencing of modern dogs and wolves indicated a divergence time of 19,000–32,000 YBP. In 2014, another study indicated 11,000–16,000 YBP based on the modern wolf's mutation rate. The first draft genome sequence of a Pleistocene wolf was published in 2015. This wolf from the Taymyr Peninsula belonged to a population that had diverged from the ancestors of both modern wolves and dogs. Radiocarbon dating indicates its age to be 35,000 YBP, and this age could then be used to calibrate the wolf's mutation rate, indicating that the genetic divergence between the dog's ancestor and modern wolves occurred before the Last Glacial Maximum, between 27,000 and 40,000 YBP. When the Pleistocene wolf's mutation rate was applied to the timing of the earlier 2014 study which had originally used the modern wolf's mutation rate, that study gave the same result of 27,000–40,000 YBP.[1] In 2017, a study compared the nuclear genome (from the cell nucleus) of three ancient dog specimens and found evidence of a single dog-wolf divergence occurring between 36,900 and 41,500 YBP.[27]
Prior to genetic divergence, the population of wolves ancestral to the dog outnumbered all other wolf populations, and after divergence the dog population underwent a population reduction to be much lower.[28][29]
In 2020, a genomic study of Eurasian wolves found that they and the dog share a common ancestor which is dated to 36,000 YBP. This finding supports the theory that all modern wolves descend from a single population which expanded after the Last Glacial Maximum and replaced other wolf populations that were adapted to different climatic conditions, and the finding of dog-like fossils dated over 30,000 YBP.[30]
Based on modern DNA
Genetic studies have found that the modern dogs from Southeast Asia and South China show greater genetic diversity than those dogs from other regions, suggesting that this was the place of their origin.[31][32][33][34][35][36] A similar study found greater genetic diversity in African village dogs than in breed dogs.[37] An East Asian origin has been questioned because dog fossils have been found in Europe dating around 15,000 YBP but only 12,000 YBP in far eastern Russia.[38] The reply is that archaeological studies in East Asia lag behind those in Europe, and that the environmental conditions in southern East Asia do not favor the preservation of fossils. Although primitive forms of the dog may have existed in Europe in the past, the genetic evidence indicates that these were later replaced by dogs that have migrated from southern East Asia.[36] In 2017, a literature review found that this East Asian study sampled only east Asian indigenous dogs and compared their patterns of genetic diversity to those of breed dogs from other geographic regions. As it is known that the genetic bottlenecks associated with formation of breeds strongly reduce genetic diversity, this was not an appropriate comparison.[5]
One DNA study concluded that dogs originated in Central Asia because dogs from there exhibit the lowest levels of linkage disequilibrium.[39] In 2017, a literature review found that because it is known that the genetic bottlenecks associated with formation of breeds raise linkage disequilibrium, the comparison of purebred with village dogs was not appropriate.[5]
Another DNA study indicated that dogs originated in the Middle East due to the sharing of DNA between dogs and Middle Eastern grey wolves.[40] In 2011, a study found this indication to be incorrect because there had been hybridization between dogs and Middle Eastern grey wolves.[41][33] In 2012, a study indicated that dogs derived from wolves originating in the Middle East and Europe and this was consistent with the archaeological record.[42] In 2014, a genomic study found that no modern wolf from any region was any more genetically closer to the dog than any other, implying that the dog's ancestor was extinct.[41]
Based on ancient DNA
In 2018, a literature review found that most genetic studies conducted over the last two decades were based on modern dog breeds and extant wolf populations, with their findings dependent on a number of assumptions. These studies assumed that the extant wolf was the ancestor of the dog, did not consider genetic admixture between wolves and dogs, nor the impact of incomplete lineage sorting. These pre-genomic studies have suggested an origin of dogs in Southeast Asia, East Asia, Central Asia, the Middle East, or Europe. More recently, the field of Paleogenomics applies the latest molecular technologies to fossil remains that still contain useful ancient DNA.[1]
Arctic Siberia

In 2015, a study recovered mDNA from ancient canid specimens that were discovered on Zhokhov Island and the Yana river, arctic Siberia. These specimens included the mandible of a 360,000–400,000 YBP Canis c.f. variabilis (where c.f. is a Latin term meaning uncertain). Phylogenetic analyses of these canids revealed nine mDNA haplotypes not detected before. The Canis c.f. variabilis specimen clustered with other wolf samples from across Russia and Asia. The mDNA haplotypes of one 8,750 YBP specimen and some 28,000 YBP specimens matched with those of geographically widely-spread modern dogs. One 47,000 YBP canid from Duvanny Yar (which was once a part of western Beringia) was distinct from wolves but was only a few mutations away from those haplotypes found in modern dogs. The authors concluded that the structure of the modern dog gene pool was contributed to from ancient Siberian wolves and possibly from Canis c.f. variabilis.[43][44]
Southern Siberia

- See further: Altai dog
In 2013, a study looked at the well-preserved skull and left mandible of a dog-like canid that was excavated from Razboinichya Cave in the Altai Mountains of southern Siberia.[45] It was dated to 33,300 YBP, which predates the oldest evidence from Western Europe and the Near East[43] The mDNA analysis found it to be more closely related to dogs than wolves.[45] Later in 2013, another study found that the canid could not be classified as a dog nor wolf because it fell between both.[20] In 2017, evolutionary biologists reviewed all of the evidence available on dog divergence and supported the specimens from the Altai mountains as being those of dogs from a lineage that is now extinct, and that was derived from a population of small wolves that is also now extinct.[5]
Europe

Phylogenetic analysis showed that modern dog mDNA haplotypes resolve into four monophyletic clades designated by researchers as clades A-D.[13][28][20]
In 2013, a study sequenced the complete and partial mitochondrial genomes of 18 fossil canids from the Old and New Worlds whose dates range from 1,000 to 36,000 YBP, and compared these with the complete mitochondrial genome sequences from modern wolves and dogs. Clade A included 64% of the modern dogs sampled, and these are a sister group to a clade containing three fossil pre-Columbian New World dogs dated between 1,000 and 8,500 YBP. This finding supports the hypothesis that pre-Columbian New World dogs share ancestry with modern dogs and that they likely arrived with the first humans to the New World. Together, clade A and the pre-Columbian fossil dogs were the sister group to a 14,500 YBP wolf found in the Kessleroch cave near Thayngen in the canton of Schaffhausen, Switzerland, with a most recent common ancestor estimated to 32,100 YBP.[20]
Clade B included 22% of the dog sequences which related to modern wolves from Sweden and the Ukraine, with a common recent ancestor estimated to 9,200 YBP. However, this relationship might represent mitochondrial genome introgression from wolves because dogs were domesticated by this time. Clade C included 12% of the dogs sampled and these were sister to two ancient dogs from the Bonn-Oberkassel cave (14,700 YBP) and the Kartstein cave (12,500 YBP) near Mechernich in Germany, with a common recent ancestor estimated to 16,000–24,000 YBP. Clade D contained sequences from 2 Scandinavian breeds – the Jamthund and Norwegian Elkhound – and is the sister group to another 14,500 YBP wolf sequence also from the Kesserloch cave, with a common recent ancestor estimated to 18,300 YBP. Its branch is phylogenetically rooted in the same sequence as the "Altai dog" (not a direct ancestor). The data from this study indicated a European origin for dogs that was estimated at 18,800–32,100 YBP based on the genetic relationship of 78% of the sampled dogs with ancient canid specimens found in Europe.[46][20] The data supports the hypothesis that dog domestication preceded the emergence of agriculture[47] and was initiated close to the Last Glacial Maximum when hunter-gatherers preyed on megafauna.[20][48]
The study found that three ancient Belgium canids (the 36,000 YBP "Goyet dog" cataloged as Canis species, along with two specimens dated 30,000 YBP and 26,000 YBP cataloged as Canis lupus) formed an ancient clade that was the most divergent group. The study found that the skulls of the "Goyet dog" and the "Altai dog" had some dog-like characteristics and proposed that this may have represented an aborted domestication episode. If so, there may have been originally more than one ancient domestication event for dogs[20] as there was for domestic pigs.[49]
One review considered why the domestication of the wolf occurred so late and at such high latitudes, when humans were living alongside wolves in the Middle East for the past 75,000 years. The proposal is that domestication was a cultural innovation caused through a long and stressful event, which was climate change. Domestication may have happened during one of the five cold Heinrich events that occurred after the arrival of humans in West Europe 37,000, 29,000, 23,000, 16,500, and 12,000 YBP. The theory is that the extreme cold during one of these events caused humans to either shift their location, adapt through a breakdown in their culture and change of their beliefs, or adopt innovative approaches. The adoption of the large wolf/dog was an adaptation to this hostile environment.[50]
A criticism of the European proposal is that dogs in East Asia show more genetic diversity. However, dramatic differences in genetic diversity can be influenced both by an ancient and recent history of inbreeding.[36] A counter-comment is that the modern European breeds only emerged in the 19th century, and that throughout history global dog populations experienced numerous episodes of diversification and homogenization, with each round further reducing the power of genetic data derived from modern breeds to help infer their early history.[38]
In 2019, study of wolf samples from northern Italy using very short lengths of mDNA found that two specimens found in the Cava Filo archaeological site near San Lazzaro di Savena, Bologna fell within the domestic dog clade A haplogroup, with one being radio-carbon dated 24,700 YBP and the other stratigraphy dated to 20,000 YBP.[51][52] The 24,700 YBP specimen matched the haplotype of ancient Bulgarian dogs, 2 historical sled dogs from the North American arctic, and 97 modern dogs. The 20,000 YBP specimen matched the haplotype of ancient Iberian and ancient Bulgarian dogs, Roman dogs from Iberia, and 2 historical sled dogs from the North American arctic. Four dog specimens found in the Bronze Age town of Via Ordiere, Solarolo, Italy dated to 3,600–3,280 years ago shared haplotypes with Late Pleistocene wolves and modern dogs.[52]
In 2020, dog remains were found in two caves, Paglicci Cave and Grotta Romanelli in Apulia, southern Italy. These were dated 14,000 YBP and are the oldest dog remains found in the Mediterranean Basin. One specimen was retrieved from a layer where the sediment was dated 20,000 YBP, indicating the possibility of an earlier timing. The specimens were genetically related to the 14,000 YBP Bonn-Oberkassel dog from Germany and other early dogs from western and central Europe which all fall within the domestic dog mDNA haplogroup C, indicating that these were all derived from a common ancestor. Using genetic timing, this clade's most recent common ancestor dates to 28,500 YBP.[53]
Morphological divergence
_Wolf.png.webp)
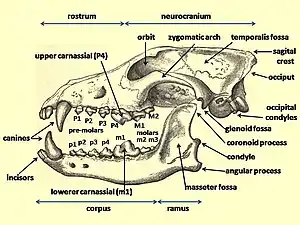
The first dogs were certainly wolflike; however, the phenotypic changes that coincided with the dog–wolf genetic divergence are not known.[5] Identifying the earliest dogs is difficult because the key morphological characters that are used by zooarchaeologists to differentiate domestic dogs from their wild wolf ancestors (size and position of teeth, dental pathologies, and size and proportion of cranial and postcranial elements) were not yet fixed during the initial phases of the domestication process. The range of natural variation among these characters that may have existed in ancient wolf populations, and the time it took for these traits to appear in dogs, are unknown.[38]
The fossil record suggests an evolutionary history that may include both morphologically dog-like wolves and wolf-like dogs. If the earliest dogs followed humans scavenging on carcasses that they left behind, then early selection may have favoured a wolf-like morphology. Perhaps when humans became more sedentary and dogs became closely associated with them was there selection for smaller, phenotypically distinct dogs, even if a reduced body size in dogs may have occurred before agriculture.[5]
When, where, and how many times wolves may have been domesticated remains debated because only a small number of ancient specimens have been found, and both archaeology and genetics continue to provide conflicting evidence. The most widely accepted earliest dog remains are those of the Bonn-Oberkassel dog which date to 15,000 YBP. Earlier remains dating back to 30,000 YBP have been described as Paleolithic dogs but their status as dogs or wolves remains debated.[2]
Dual ancestry of the domestic dogs of West Asia, Africa and southern Europe
More recent research analysing the genomes of 72 ancient wolves, specimens from Europe, Siberia and North America spanning the past 100,000 years has confirmed that both early and modern dogs are more similar genetically to ancient wolves from Asia than from Europe. This suggests that domestication occurred in the East. The research also found evidence that dogs have a dual ancestry, meaning that two separate populations of wolves contributed DNA to dogs. Early dogs from northeastern Europe, Siberia and the Americas appear to have a single, shared origin from the eastern source. But early dogs from the Middle East, Africa and southern Europe appear to have some ancestry from another source related to wolves in the Middle East, in addition to the eastern source. It is possible that wolves underwent domestication more than once, with different populations then mixing together. Or, that domestication happened once only, and that dual ancestry is related to early dogs then mixing with wild wolves. The research also demonstrated how wolf DNA changed during the 30,000 generations that were represented in their 100,000-year timeline. This identified the effects of natural selection as particular genes spread within wolf populations. One gene variant, over a period of around 10,000 years, went from being very rare to being present in every wolf, and it is still present in all wolves and dogs today. The variant affects a gene, IFT88, which is involved in the development of bones in the skull and jaw. It is possible that the spread of this variant could have been driven by a change in the types of prey available during the Ice Age, giving an advantage to wolves with a certain head shape. "This is the first time scientists have directly tracked natural selection in a large animal [the wolf] over a time-scale of 100,000 years, seeing evolution play out in real time rather than trying to reconstruct it from DNA today," said study senior author Pontus Skoglund.[54]
Dog domestication
... Remove domestication from the human species, and there's probably a couple of million of us on the planet, max. Instead, what do we have? Seven billion people, climate change, travel, innovation and everything. Domestication has influenced the entire earth. And dogs were the first. For most of human history, we're not dissimilar to any other wild primate. We're manipulating our environments, but not on a scale bigger than, say, a herd of African elephants. And then, we go into partnership with this group of wolves. They altered our relationship with the natural world. ...
Animal domestication is a coevolutionary process in which a population responds to selective pressure while adapting to a novel niche that included another species with evolving behaviors.[11]
One of the most important transitions in human history was the domestication of animals, which began with the long-term association between wolves and hunter–gatherers more than 15,000 years ago.[4] Dogs were the first domesticated species,[5][13] the only animal known to have entered into a domestic relationship with humans during the Pleistocene,[13] and the only large carnivore to have been domesticated.[5] It was not until 11,000 YBP that people living in the Near East entered into relationships with wild populations of aurochs, boar, sheep, and goats. A domestication process then began to develop. The earlier association of dogs with humans may have allowed dogs to have a profound influence on the course of early human history and the development of civilization.[5]
The questions of when and where dogs were first domesticated have taxed geneticists and archaeologists for decades.[11] Genetic studies suggest a domestication process commencing over 25,000 YBP, in one or several wolf populations in either Europe, the high Arctic, or eastern Asia.[16] There is clear evidence that dogs were derived from grey wolves during the initial phases of domestication. The wolf population(s) that were involved are likely to be extinct. Despite numerous genetic studies of both modern dogs and ancient dog remains, there is no firm consensus regarding either the timing or location(s) of domestication, the number of wolf populations that were involved, or the long-term effects domestication has had on the dog's genome.[16]
Around 10,000 YBP agriculture was developed resulting in a sedentary lifestyle, along with phenotype divergence of the dog from its wolf ancestors, including variance in size.[5] Two population bottlenecks have occurred to the dog lineage, one due to the initial domestication and one due to the formation of dog breeds.[5][16]
Socialization
Humans and wolves both exist in complex social groups. How humans and wolves got together remains unknown. One view holds that domestication is a process that is difficult to define. The term was developed by anthropologists with a human-centric view in which humans took wild animals (ungulates) and bred them to be "domestic", usually in order to provide improved food or materials for human consumption. That term may not be appropriate for a large carnivore such as the dog. This alternate view regards dogs as being either socialized and able to live among humans, or unsocialized. There exist today dogs that live with their human families but are unsocialized and will threaten strangers defensively and aggressively no differently than a wild wolf. There also exists a number of cases where wild wolves have approached people in remote places, attempting to initiate play and to form companionship.[57] One such notable wolf was Romeo, a gentle black wolf that formed relationships with the people and dogs of Juneau, Alaska.[58] This view holds that before there could have been domestication of the wolf, there had to have been its socialization.[57][59]
Even today, the wolves on Ellesmere Island do not fear humans, which is thought to be due to them seeing humans so little, and they will approach humans cautiously, curiously and closely.[60][61][62][63]
Commensal pathway

The dog is a classic example of a domestic animal that likely traveled a commensal pathway into domestication.[2][38] The dog was the first domesticant, and was domesticated and widely established across Eurasia before the end of the Pleistocene, well before cultivation or the domestication of other animals.[38] It may have been inevitable that the first domesticated animal came from the order of carnivores as these are less afraid when approaching other species. Within the carnivores, the first domesticated animal would need to exist without an all-meat diet, possess a running and hunting ability to provide its own food, and be of a controllable size to coexist with humans, indicating the family Canidae, and the right temperament[64] with wolves being among the most gregarious and cooperative animals on the planet.[65][66]
Human campfire theory
Ancient DNA supports the hypothesis that dog domestication preceded the emergence of agriculture[20][47] and was initiated close to the Last Glacial Maximum when hunter-gatherers preyed on megafauna, and when proto-dogs might have taken advantage of carcasses left on site by early hunters, assisted in the capture of prey, or provided defense from large competing predators at kill-sites.[20] Wolves were probably attracted to human campfires by the smell of meat being cooked and discarded refuse in the vicinity, first loosely attaching themselves and then considering these as part of their home territory where their warning growls would alert humans to the approach of outsiders.[67] The wolves most likely drawn to human camps were the less-aggressive, subdominant pack members with lowered flight response, higher stress thresholds, less wary around humans, and therefore better candidates for domestication.[68]
Migratory wolves theory

On the mammoth steppe the wolf's ability to hunt in packs, to share risk fairly among pack members, and to cooperate moved them to the top of the food chain above lions, hyenas and bears. Some wolves followed the great reindeer herds, eliminating the unfit, the weaklings, the sick and the aged, and therefore improved the herd. These wolves had become the first pastoralists hundreds of thousands of years before humans also took to this role.[69] The wolves' advantage over their competitors was that they were able to keep pace with the herds, move fast and enduringly, and make the most efficient use of their kill by their ability to "wolf down" a large part of their quarry before other predators had detected the kill. One study proposed that during the Last Glacial Maximum, some of our ancestors teamed up with those pastoralist wolves and learned their techniques.[65][70]
Many of our ancestors remained gatherers and scavengers, or specialized as fish-hunters, hunter-gatherers, and hunter-gardeners. However, some ancestors adopted the pastoralist wolves' lifestyle as herd followers and herders of reindeer, horses, and other hoofed animals. They harvested the best stock for themselves while the wolves kept the herd strong, and this group of humans was to become the first herders and this group of wolves was to become the first dogs.[69][65]
The remains of large carcasses left by human hunter-gatherers may have led some wolves into entering a migratory relationship with humans. This could have led to their divergence from those wolves that remained in the one territory. A closer relationship between these wolves — or proto-dogs — and humans may have then developed, such as hunting together and mutual defence from other carnivores and other humans.[5] A maternal mDNA, paternal yDNA, and microsatellite assessment of two wolf populations in North America and combined with satellite telemetry data revealed significant genetic and morphological differences between one population that migrated with and preyed upon caribou, and another territorial ecotype population that remained in a boreal coniferous forest. Though these two populations spend a period of the year in the same place, and though there was evidence of gene flow between them, the difference in prey–habitat specialization has been sufficient to maintain genetic and even coloration divergence.[11][71] A study has identified the remains of a population of extinct Pleistocene Beringian wolves with unique mDNA signatures. The skull shape, tooth wear, and isotopic signatures suggested these were specialist megafauna hunters and scavengers that became extinct while less specialized wolf ecotypes survived.[11][72] Analogous to the modern wolf ecotype that has evolved to track and prey upon caribou, a Pleistocene wolf population could have begun following mobile hunter-gatherers, thus slowly acquiring genetic and phenotypic differences that would have allowed them to more successfully adapt to the human habitat.[11]
Food partitioning theory
Dogs were the only animal to be domesticated by mobile hunter-gatherers. Humans and wolves were both persistent pack hunters of large prey, were competing in overlapping territory, and are both capable of killing each other. One study proposes how humans may have domesticated such a dangerous competitor. Humans and wolves are members of the large carnivore guild, and when there is abundant game the top members leave carcasses for the other members to scavenge. When game is scarce there is often conflict. Humans are unusual members of this guild because their ancestors were primates, therefore their ability to process meat is limited by the capacity of the liver to metabolize protein, and they can only derive 20% of their energy requirements from protein. High protein consumption in humans can lead to illness.[73]
During the harsh winters of the Last Glacial Maximum plant foods would have not been available, and meat would not be the favoured food but fat and grease would be, as is prized by some high-latitude dwelling peoples in modern times. Game meat would have been devoid of fat, but the limbs and crania contain fat deposits, and limb bones contain fatty oils. There is evidence of such processing during this period. Wolves are typical carnivores and can survive on a protein-based diet for months. Calculations of the lipid content of arctic and subarctic game available across the cold steppe environment at this time and today shows that in order to gain the necessary quantity of fat and oils, there would have been enough excess animal calories to feed either proto-dogs or wolves with no need for competition. Hunting together and protection from other predators would have been advantageous to both species, leading to domestication.[73]
Genetic changes


The Yellow Dog Study
Domestic dogs exhibit diverse coat colours and patterns. In many mammals, different colour patterns are the result of the regulation of the Agouti gene, which can cause hair follicles to switch from making black or brown pigments to yellow or nearly white pigments. The most common coat pattern found in modern wolves is agouti, in which the upperside of the body has banded hairs and the underside exhibits lighter shading. The colour yellow is dominant to the colour black and is found in dogs across much of the world and the dingo in Australia.[74]
In 2021, a study of whole genome sequences taken from dogs and wolves focused on the genetic relationships between them based on coat colour. The study found that most dog colour haplotypes were similar to most wolf haplotypes, however dominant yellow in dogs was closely related to white in arctic wolves from North America. This result suggests a common origin for dominant yellow in dogs and white in wolves but without recent gene flow, because this light colour clade was found to be basal to the golden jackal and genetically distinct from all other canids. The most recent common ancestor of the golden jackal and the wolf lineage dates back to 2 million YBP. The study proposes that 35,000 YBP there was genetic introgression into the Late Pleistocene grey wolf from a ghost population of an extinct canid which had diverged from the grey wolf lineage over 2 million YBP. This colour diversity could be found 35,000 YBP in wolves and 9,500 YBP in dogs. A closely related haplotype exists among those wolves of Tibet which possess yellow shading in their coats. The study explains the colour relationships between modern dogs and wolves, white wolves from North America, yellow dogs, and yellowish wolves from Tibet. The study concludes that during the Late Pleistocene, natural selection laid the genetic foundation for modern coat colour diversity in dogs and wolves.[74]
Dietary adaptation
Selection appears to have acted on the dog's metabolic functions to cope with changes in dietary fat, followed later with a dietary increase in starch associated with a more commensal lifestyle.[5]
The dog genome compared to the wolf genome shows signs of having undergone positive selection, these include genes relating to brain function and behavior, and to lipid metabolism. This ability to process lipids indicates a dietary target of selection that was important when proto-dogs hunted and fed alongside hunter-gatherers. The evolution of the dietary metabolism genes may have helped process the increased lipid content of early dog diets as they scavenged on the remains of carcasses left by hunter-gatherers.[26] Prey capture rates may have increased in comparison to wolves and with it the amount of lipid consumed by the assisting proto-dogs.[26][48][75] A unique dietary selection pressure may have evolved both from the amount consumed, and the shifting composition of, tissues that were available to proto-dogs once humans had removed the most desirable parts of the carcass for themselves.[26] A study of the mammal biomass during modern human expansion into the northern Mammoth steppe found that it had occurred under conditions of unlimited resources, and that many of the animals were killed with only a small part consumed or were left unused.[76]
- See further: Dietary phenotypic plasticity
Behaviour
The key phase in domestication appears to have been changes in social behaviour and its corresponding oxytocin receptor genes and neural-related genes. Behaviour differences between dogs and wolves may be contributed by structural variation in the genes that are associated with human Williams-Beuren syndrome. This syndrome causes increased hyper-sociability, which may have been important during domestication.[16]
In 2014, a whole genome study of the DNA differences between wolves and dogs found that the dogs tameness was not a reduced fear response but did show greater synaptic plasticity. Synaptic plasticity is widely believed to be the cellular correlate of learning and memory. The study proposes that the improved learning and memory abilities of dogs also helped to lower their level of fear around humans.[77]
Unlike other domestic species which were primarily selected for production-related traits, dogs were initially selected for their behaviors.[78][79] In 2016, a study found that there were only 11 fixed genes that showed variation between wolves and dogs. These gene variations were unlikely to have been the result of natural evolution, and indicate selection on both morphology and behavior during dog domestication. There was evidence of selection during dog domestication of genes that affect the adrenaline and noradrenaline biosynthesis pathway. These genes are involved in the synthesis, transport and degradation of a variety of neurotransmitters, particularly the catecholamines, which include dopamine and noradrenaline. Recurrent selection on this pathway and its role in emotional processing and the fight-or-flight response[79][80] suggests that the behavioral changes we see in dogs compared to wolves may be due to changes in this pathway, leading to tameness and an emotional processing ability.[79] Dogs generally show reduced fear and aggression compared to wolves.[79][81] Some of these genes have been associated with aggression in some dog breeds, indicating their importance in both the initial domestication and then later in breed formation.[79]
Role of epigenetics
Differences in hormonal expression that are associated with domestication syndrome may be linked to epigenetic modifications. A recent study that compared the methylation patterns of dogs with those of wolves found 68 significantly different methylated sites. These included sites which are linked to two neurotransmitter genes associated with cognition.[2] There is a direct association between the dog's social behaviour and OXTR, which is a receptor for the neurotransmitter Oxytocin, and this has been caused through the epigenetic methylation of the OXTR gene.[82] DNA methylation differences have been found between wolves and dogs, and between different dog breeds. This implies that epigentic factors may have been important for both dog domestication and the divergence of dog breeds.[83]
Similar to humans, wolves show strong social and emotional bonds within their groupings, and this relationship might have been the foundation for the evolution of dog-human bonding.[84][85] In 2019, a literature review led to a new theory named Active Social Domestication, in which the social environment of the dog ancestor induced neuro-physiological changes that caused an epigenetic cascade, which led to the rapid development of domestication syndrome.[84][86]
Parallel evolution
Being the first domesticated species has created a strong bond between dogs and humans and entwined their histories. There is an extensive list of genes that showed signatures of parallel evolution in dogs and humans. A suite of 311 genes under positive selection in dogs are related to a large number of overlapping loci which show the same patterns in humans, and these play a role in digestion, neurological processes, and some being involved with cancers. This fact can be used to study the coevolution of gene function. Dogs accompanied humans when they first migrated into new environments. Both dogs and humans have adapted to different environmental conditions, with their genomes showing parallel evolution. These include adaptation to high altitude, low oxygen hypoxia conditions, and genes that play a role in digestion, metabolism, neurological processes, and some related to cancer. It can be inferred from those genes which act on the serotonin system in the brain that these have given rise to less aggressive behavior when living in a crowded environment.[1] Dogs suffer from the same common diseases – such as cancer, diabetes, heart disease, and neurological disorders – as do humans. The underlying disease pathology is similar to humans, as is their responses and outcomes to treatment.[16]
Behavioral evidence
Convergent evolution is when distantly related species independently evolve similar solutions to the same problem. For example, fish, penguins and dolphins have each separately evolved flippers as a solution to the problem of moving through the water. What has been found between dogs and humans is something less frequently demonstrated: psychological convergence. Dogs have independently evolved to be cognitively more similar to humans than we are to our closest genetic relatives.[87] Dogs have evolved specialized skills for reading human social and communicative behavior. These skills seem more flexible – and possibly more human-like – than those of other animals more closely related to humans phylogenetically, such as chimpanzees, bonobos and other great apes. This raises the possibility that convergent evolution has occurred: both Canis familiaris and Homo sapiens might have evolved some similar (although obviously not identical) social-communicative skills – in both cases adapted for certain kinds of social and communicative interactions with human beings.[88]
Studies support coevolution in that dogs can follow the human pointing gesture,[89] discriminate the emotional expressions of human faces,[90] and that most people can tell from a bark whether a dog is alone, being approached by a stranger, playing, or being aggressive,[91] and can tell from a growl how big the dog is.[92]
In 2015, a study found that when dogs and their owners interact, extended eye contact (mutual gaze) increases oxytocin levels in both the dog and its owner. As oxytocin is known for its role in maternal bonding, it is considered likely that this effect has supported the coevolution of human-dog bonding.[93]
The dog could have arisen only from animals predisposed to human society by lack of fear, attentiveness, curiosity, necessity, and recognition of advantage gained through collaboration....the humans and wolves involved in the conversion were sentient, observant beings constantly making decisions about how they lived and what they did, based on the perceived ability to obtain at a given time and place what they needed to survive and thrive. They were social animals willing, even eager, to join forces with another animal to merge their sense of group with the others' sense and create an expanded super-group that was beneficial to both in multiple ways. They were individual animals and people involved, from our perspective, in a biological and cultural process that involved linking not only their lives but the evolutionary fate of their heirs in ways, we must assume, they could never have imagined. Powerful emotions were in play that many observers today refer to as love – boundless, unquestioning love.
— Mark Derr[94]
Human adoption of some wolf behaviors
... Isn't it strange that, our being such an intelligent primate, we didn't domesticate chimpanzees as companions instead? Why did we choose wolves even though they are strong enough to maim or kill us? ...
— Wolfgang Schleidt[65]
In 2002, a study proposed that immediate human ancestors and wolves may have domesticated each other through a strategic alliance that would change both respectively into humans and dogs. The effects of human psychology, hunting practices, territoriality and social behavior would have been profound.[95]
Early humans moved from scavenging and small-game hunting to big-game hunting by living in larger, socially more-complex groups, learning to hunt in packs, and developing powers of cooperation and negotiation in complex situations. As these are characteristics of wolves, dogs and humans, it can be argued that these behaviors were enhanced once wolves and humans began to cohabit. Communal hunting led to communal defense. Wolves actively patrol and defend their scent-marked territory, and perhaps humans had their sense of territoriality enhanced by living with wolves.[95] One of the keys to recent human survival has been the forming of partnerships. Strong bonds exist between same-sex wolves, dogs and humans, and these bonds are stronger than exist between other same-sex animal pairs. Today, the most widespread form of inter-species bonding occurs between humans and dogs. The concept of friendship has ancient origins, but it may have been enhanced through the inter-species relationship to give a survival advantage.[95][96]
In 2003, a study compared the behavior and ethics of chimpanzees, wolves and humans. Cooperation among humans' closest genetic relative is limited to occasional hunting episodes or the persecution of a competitor for personal advantage, which had to be tempered if humans were to become domesticated.[65][97] One might therefore argue that the closest approximation to human morality that can be found in nature is that of the grey wolf. Wolves are among the most gregarious and cooperative of animals on the planet,[65][66] and their ability to cooperate in well-coordinated drives to hunt prey, carry items too heavy for an individual, provisioning not only their own young but also the other pack members, babysitting etc. are rivaled only by that of human societies. Similar forms of cooperation are observed in two closely related canids, the African wild dog and the Asian dhole, therefore it is reasonable to assume that canid sociality and cooperation are old traits that in terms of evolution predate human sociality and cooperation. Today's wolves may even be less social than their ancestors, as they have lost access to large herds of ungulates and now tend more toward a lifestyle similar to coyotes, jackals, and even foxes.[65] Social sharing within families may be a trait that early humans learned from wolves,[65][98] and, with wolves digging dens long before humans constructed huts, it is not clear who domesticated whom.[69][65][97]
First dogs
Dogs domesticated in Siberia 23,000 years ago
Locating the origin of dogs is made difficult by the lack of data on extinct Pleistocene wolves, the small morphological changes that occurred between wild and domestic populations during the first phases of domestication, and the lack of an accompanying human material culture at this time.[4]
In 2016, a genetic study found that ancient and modern dogs fall into an Eastern Eurasian clade and a Western Eurasian clade.[28] In 2017, another genetic study found evidence of a single dog-wolf divergence occurring between 36,900 and 41,500 YBP, followed by a divergence between Eastern Eurasian and Western Eurasian dogs 17,500–23,900 YBP and this indicates a single dog domestication event occurring between 20,000 and 40,000 YBP.[27]
In 2021, a review of the current evidence infers from the timings provided by DNA studies that the dog was domesticated in Siberia 23,000 years ago by ancient North Siberians. The dog later dispersed from Siberia with the migration of peoples eastwards into the Americas and westwards across Eurasia. The ancient North Siberians were once a people whose ancestors archaeological remains have been found at the Paleolithic Yana RHS (Rhinoceros Horn Site) on the Yana River delta in arctic northern Siberia that is dated 31,600 YBP, and at the Mal'ta site near Lake Baikal in southern Siberia just north of Mongolia that is dated 24,000 YBP. Ancient dog remains dating to this time and place have yet to be discovered to support this hypothesis.[13]
The review theorizes that the harsh climate of the Last Glacial Maximum may have brought humans and wolves closer together while they were isolated inside refuge areas. Both species hunt the same prey, and their increased interactions may have resulted in the shared scavenging of kills, wolves drawn to human campsites, a shift in their relationship, and eventually domestication.[13]
Mitochondrial DNA indicates that almost all modern dogs fall into one of four monophyletic haplogroups which are named haplogroups A, B, C, and D. The majority of dogs fall within haplogroup A. The mDNA "molecular clock" indicates that 22,800 YBP the first genetic divergence (split) occurred in haplogroup A, resulting in the lineages A1b and A2. This timing is the oldest known between any two dog mDNA lineages. As humans migrated across Siberia, through Beringia, and down through the Americas, archaeological remains indicate that their mDNA lineages diverged several times. Based on these timings, and the timings of several dog divergences found from early dog remains across these regions, it was discovered that there was a correlation between human and dog migrations and population divergences. This correlation suggests that where people went, their dogs also went. Tracing back through these human and dog lineages and timings led to the inference that the dog was first domesticated in Siberia nearly 23,000 YBP by North Siberians.[13]
Another study undertook an analysis of the complete mitogenome sequences of 555 modern and ancient dogs. The sequences showed an increase in the population size approximately 23,500 YBP, which broadly coincides with the proposed genetic divergence of the ancestors of dogs from modern wolves. A ten-fold increase in the population size occurred after 15,000 YBP, which is consistent with the demographic dependence of dogs on the human population.[99]
Earlier in 2018, a study proposes that the Yana site showed evidence of wolf pre-domestication. There are remains of medium-sized canids found there that could not be referred to as dogs, however they showed indications of living with people. These included worn and partially-missing teeth, and the skull of an almost-adult showing juvenile features. The morphologic and morphometric anomalies in the specimens indicate commensalism and the earliest stage of domestication.[100]
Admixture
Studies indicate admixture between the dog-wolf ancestor and golden jackals.[5] However, since domestication, there was almost negligible gene flow from wolves into dogs but substantial gene flow from dogs into wolves. There were some wolves that were related to all ancient and modern dogs. A very small amount of gene flow was detected between coyotes and ancient American dogs, and between the African wolf and African dogs but in which direction could not be determined.[3] The short divergence time between dogs and wolves followed by their continuous admixture has led to 20% of the genome of East Asian wolves and 7–25% of the genome of European and Middle Eastern wolves showing contributions from dogs.[1] The β-defensin gene responsible for the black coat of North American wolves was the result of a single introgression from early Native American dogs in the Yukon between 1,600 and 7,200 YBP.[101] Dogs and wolves living in the Himalayas and on the Tibetan plateau carry the EPAS1 allele that is associated with high-altitude oxygen adaptation, which has been contributed by a ghost population of an unknown wolf-like canid. This ghost population is deeply-diverged from modern Holarctic wolves and dogs, and has contributed 39% to the Himalayan wolf's nuclear genome.[102] Limited gene flow has likely occurred in arctic dogs.[4]
Bonn-Oberkassel dog

The generally accepted earliest dog remains were discovered in Bonn-Oberkassel, Germany.[103][104] Contextual, isotopic, genetic, and morphological evidence shows that this dog was clearly not a local wolf.[13] The dog was dated to 14,223 YBP.[103]
In 1914, on the eve of the First World War, two human skeletons were discovered during basalt quarrying at Oberkassel, Bonn in Germany. With them were found a right mandible of a "wolf" and other animal bones.[105] After the end of the First World War, in 1919 a full study was made of these remains. The mandible was recorded as "Canis lupus, the wolf" and some of the other animal bones were assigned to it.[106] The remains were then stored and forgotten for fifty years. In the late 1970s there was renewed interest in the Oberkassel remains and the mandible was re-examined and reclassified as belonging to a domesticated dog.[107][108][109] The mitochondrial DNA sequence of the mandible was matched to Canis familiaris – a dog[20] and falls within mDNA haplogroup C of dogs.[45] The bodies were dated to 14,223 YBP.[103] This implies that in Western Europe there were morphologically and genetically "modern" dogs in existence around 14,500 YBP.[110]
Later studies assigned more of the other animal bones to the dog until most of a skeleton could be assembled.[110] The humans were a man aged 40 years and a woman aged 25 years. All three skeletal remains were found sprayed with red hematite powder and covered with large 20 cm thick basalt blocks.[103] The consensus is that a dog was buried along with two humans.[110] A tooth belonging to a smaller and older dog was also identified but it had not been sprayed with red powder.[103] The cause of the death of the two humans is not known.[110] A pathology study of the dog remains suggests that it had died young after suffering from canine distemper between ages 19 and 23 weeks.[103] The dog could not have survived during this period without intensive human care.[110][103] During this period the dog was of no utilitarian use to humans,[103] and suggests the existence of emotional or symbolic ties between these humans and this dog.[110] In conclusion, near the end of the Late Pleistocene at least some humans regarded dogs not just materialistically, but had developed emotional and caring bonds for their dogs.[103]
Ice Age dogs
In 2020, the sequencing of ancient dog genomes indicates that dogs share a common ancestry and descended from an ancient, now-extinct wolf population - or closely related wolf populations - which was distinct from the modern wolf lineage. By the close of the last Ice Age (11,700 YBP), five ancestral lineages had diversified from each other and were expressed in dog samples taken from the Neolithic era Levant (7,000 YBP), Mesolithic era Karelia (10,900 YBP), Mesolithic era Baikal (7,000 YBP), ancient America (4,000 YBP), and the New Guinea singing dog (present day).[3]
The world's ancient and modern dog population structure can be classified into Arctic/Americas, East Asian, and West Eurasian.[111][104] The Arctic/Americas lineage includes modern arctic breeds, a 9,500 YBP dog from Zhokhov Island, ancient pre-European contact American dogs, mid-Holocene dogs from Lake Baikal, historical dogs from across Siberia, and dogs from the Yamalo-Nenets Autonomous Okrug region in northwestern Siberia. The East Asian lineage includes modern dogs from China, Vietnam, Island South East Asia, and the dingo and the New Guinea singing dog[111] that represent unadmixed East Asian ancestry.[3] The West Eurasian lineage includes ancient Levantine and ancient Near Eastern dogs, ancient and modern European dogs, modern African dogs, and Bronze Age dogs from the Eurasian Steppe.[111]
Ancient and modern European dogs have a closer relationship with arctic dogs than do Near Eastern dogs, indicating a major admixture event in Europe. Analyses of a recently sequenced genome from the Mesolithic site of Veretye, Karelia (∼10,000 YBP) in Northeast Europe recapitulate findings that show these dogs possessed ancestry related to both Arctic (~70%) and Western Eurasian (~30%) lineages. This suggests that other modern and ancient (post-Mesolithic) European dogs sequenced to date, Mesolithic dogs in Europe already possessed both Arctic and Western Eurasian ancestry. The fact that the ancient Siberian dog from Zhokhov Island, dated ∼1,000 years after the Veretye, Karelia dogs, possesses no Western Eurasian ancestry, indicates that Western dog ancestry had not yet reached the Siberian Arctic by 9,500 YBP.[3][111][112] The 9,500 YBP Zhokhov dog is more closely related to a 6,000 YPB dog from Lake Baikal than related to the ancient dogs found in North America, which supports that a genetic split had occurred between the early Arctic and North American dogs and that their common ancestor dates much older than the 9,500 YBP Zhokhov dog[111] The earliest Neolithic European dog dated 7,000 YBP was found to be a mixture of the Karelian and the Levantine lineages. The lineage of a Neolithic dog dated 5,000 YBP found in southwestern Sweden was the ancestor of 90-100% of modern European dogs. This implies that in Europe a population of half-Karelian and half-Levantine dogs similar to this one - but not necessarily originating in Sweden - replaced all of the other dog populations. These findings together support a dual ancestry for modern European dogs, which possess 54% Karelian and 46% Levantine ancestries.[3]
Siberian dogs were genetically similar 9,500–7,000 YBP showing Arctic ancestry, however the introduction of dogs from the Eurasian Steppe and Europe led to substantial genetic admixture, with ancient and historical Siberian dogs exhibiting varying levels of Arctic and Near East ancestry. Dogs from the Bronze Age Eurasian Steppe exhibited 40% ancient arctic and 60% ancient Near East ancestry until the Middle Ages. This implies that dogs migrated as part of the Neolithic expansion of farming from the Near East into the steppes. In the Yamalo-Nenets Autonomous Okrug region in northwestern Siberia, dogs from 2,000 YBP were less related to the dogs of the Eurasian Steppe and Europe than dogs 1,000 YBP. The archaeological presence of glass beads and metal items indicate that this region was connected to a large trade network which included the Near East, the Black Sea region, and the Eurasian Steppe which led to the acquiring of dogs from these regions. The acquisition of dogs from the Near East adapted to farming, and the Eurasian Steppe adapted to pastoralism, may have provided behavioural and morphological characteristics when admixed with Arctic dogs, leading to their adaptation from foraging to reindeer pastoralism. Two dog specimens that are nearly 100 years old and obtained from the Nenets people on the Yamal Peninsula found that these are related to two specimens dated 2,000 years old and 850 years old, which suggests continuity of the lineage in this region. The two 100 year old dogs were closely related with the Samoyed breed. Siberian Huskies show a genetic affinity with historical East Siberian dogs and ancient Lake Baikal dogs. Together, this indicates that the ancient arctic lineage lives on in some modern Siberian breeds.[111]
Ancient dog genomes were compared with ancient human genomes across time, space, and cultural context to reveal that these generally matched each other. These generally share similar features but they differ across time. There were some large differences: the same dogs could be found in both the Neolithic Levant and later in Chalcolithic Iran (5,800 YBP) although the human populations of each were different; in Neolithic Ireland (4,800 YBP) and Germany (7,000 YBP) the dogs are more associated with northern European hunter-gatherers while the humans were more associated with people from the Levant; and on the Bronze Age Pontic–Caspian steppe (3,800 YBP) and in Corded Ware culture Germany (4,700 YBP) the human population had shifted away from the Neolithic European populations but the dogs had not. European dogs have a stronger genetic relationship to Siberian and ancient American dogs than to the New Guinea singing dog, which has an East Asian origin, reflecting an early polar relationship between humans in the Americas and Europe. People living in the Lake Baikal region 18,000—24,000 YBP were genetically related to western Eurasians and contributed to the ancestry of Native Americans, however these were then replaced by other populations. Ten thousand years later, around 7,000 YBP, the dogs in the Lake Baikal region still exhibited a relationship with Europe and the Americas. This implies that there was a shared population structure for both dogs and humans across circumpolar northern Eurasia.[3]
Ancient human genomes show a major ancestry transformation which coincided with the expansion of Neolithic farmers from the Near East into Europe. Ancient dog mitochondria suggests these were accompanied by dogs, which led to an associated ancestry transformation for dogs in Europe. The expansions of steppe pastoralists associated with the Corded Ware culture and the Yamnaya culture into Late Neolithic and Bronze Age Europe transformed the ancestry of human populations but their accompanying dogs had no major impact on European dog populations. The steppe pastoralists also expanded eastwards but had little impact on the ancestry of East Asian people. However, many Chinese dogs appear to be a product of admixture between the lineage of a 3,800 YBP western Eurasian Srubnaya culture dog and the ancestor of the dingo and New Guinea singing dog. Populations of modern Siberian dogs also show ancestry from 7,000 YBP Lake Baikal dogs but little or no New Guinea singing dog ancestry, indicating no East Asian ancestry.[3]
The AMY2B gene codes a protein which assists with the first step in the digestion of dietary starch and glycogen. An expansion of this gene would enable early dogs to exploit a starch-rich diet. At the beginning of agriculture, only some dogs possessed this adaptation which became widespread several thousand years later.[3]
Dogs migrated alongside humans but the movement of the two did not always align, indicating that in some cases humans migrated without dogs or that dogs moved between human groups, possibly as a cultural or trade item. Dogs appear to have been dispersed across Eurasia and into the Americas without any major human population movement being involved, which remains a mystery. Past studies have suggested the dog's place of origin but these studies were based upon today's patterns of genomic diversity or possible links to modern wolf populations. The dog's history was obscured to these studies because of recent gene flow and population dynamics – the geographical origin of the dog remains unknown.[3]
First dogs as a hunting technology

During the Upper Paleolithic (50,000–10,000 YBP), the increase in human population density, advances in blade and hunting technology, and climate change may have altered prey densities and made scavenging crucial to the survival of some wolf populations. Adaptations to scavenging such as tameness, small body size, and a decreased age of reproduction would reduce their hunting efficiency further, eventually leading to obligated scavenging.[39][113] Whether these earliest dogs were simply human-commensal scavengers or they played some role as companions or hunters that hastened their spread is unknown.[39]
Researchers have proposed that in the past a hunting partnership existed between humans and dogs that was the basis for dog domestication.[114][115][116] Petroglyph rock art dating to 8,000 YBP at the sites of Shuwaymis and Jubbah, in northwestern Saudi Arabia, depict large numbers of dogs participating in hunting scenes with some being controlled on leashes.[117] The transition from the Late Pleistocene into the early Holocene was marked by climatic change from cold and dry to warmer, wetter conditions and rapid shifts in flora and fauna, with much of the open habitat of large herbivores being replaced by forests.[116] In the early Holocene, it is proposed that along with changes in arrow-head technology that hunting dogs were used by hunters to track and retrieve wounded game in thick forests.[115][116] The dog's ability to chase, track, sniff out and hold prey can significantly increase the success of hunters in forests, where human senses and location skills are not as sharp as in more open habitats. Dogs are still used for hunting in forests today.[116]
First dog breeds developed in arctic northeastern Siberia

The domestic dog was present 9,500 YBP on what is now Zhokhov Island, arctic northeastern Siberia. The archaeological discoveries at the Zhokhov site includes the remains of dog harness straps similar to those used by the modern Inuit, the bone remains of polar bears and reindeer which suggests a wide hunting range and the transport of large body parts back to the site, and tools made from obsidian transported from 1,500 kilometres away. These findings suggest long-distance transport through the use of sled dogs.[118]
A study of dog remains indicates that these were selectively bred to be either as sled dogs or as hunting dogs, which implies that a sled dog standard and a hunting dog standard existed at that time. The optimal maximum size for a sled dog is 20–25 kg based on thermo-regulation, and the ancient sled dogs were between 16 and 25 kg. The same standard has been found in the remains of sled dogs from this region 2,000 YBP and in the modern Siberian husky breed standard. Other dogs were more massive at 30 kg and appear to be dogs that had been crossed with wolves and used for polar bear hunting. At death, the heads of the dogs had been carefully separated from their bodies by humans, probably for ceremonial reasons.[119]
The study proposes that after having diverged from the common ancestor shared with the grey wolf, the evolution of the dog proceeded in three stages. The first was natural selection based on feeding behavior within the ecological niche that had been formed through human activity. The second was artificial selection based on tamability. The third was directed selection based on forming breeds that possessed qualities to help with specific tasks within the human economy. The process commenced 30,000–40,000 YBP with its speed increasing in each stage until domestication became complete.[119]
The Zhokhov dogs are the oldest known dogs to exhibit colour patterns. These possessed black colour patterns on their backs, which helped to distinguish them from white arctic wolves.[74]
Dogs enter North America from northeastern Siberia
Material culture provides evidence for dog harnessing in the Arctic 9,000 YBP. Ancient DNA from the remains of these dogs indicates that they belong to the same genetic lineage as modern Arctic dogs, and that this lineage gave rise to the earliest native American dogs. Since the earliest native American dogs, multiple, genetically different lineages of dogs were introduced by the Thule people and European settlers. The European dogs replaced the dog lineages that were introduced more than 10,000 years ago.[4]
In North America, the earliest dog remains were found in Lawyer's Cave on the Alaskan mainland east of Wrangell Island in the Alexander Archipelago of southeast Alaska, radiocarbon dating indicates 10,150 YBP. A genetic-based estimate indicates that this dog's lineage had split from the Siberian Zhokhov Island dog lineage 16,700 YBP. This timing coincides with the suggested opening of the North Pacific coastal route into North America. Stable isotope analysis can be used to identify some chemical elements, allowing researchers to make inferences about the diet of a species. An isotope analysis of bone collagen indicates a marine diet.[120] The next earliest dogs were found in Illinois and radiocarbon dating indicates 9,900 YBP. These include three isolated burials at the Koster Site near the lower Illinois River in Greene County, and one burial 35 km away at the Stilwell II site in Pike County. These dogs were medium-sized adults around 50 cm (20 in) in height and around 17 kilograms (37 lb) in weight, with very active lifestyles and varied morphologies. Stable isotope analysis indicates a diet consisting largely of freshwater fish. Similar dog burials across Eurasia are thought to be due to the dog's importance in hunting to people who were trying to adapt to the changing environments and prey species during the Pleistocene-Holocene transition. In these places, the dog had gained an elevated social status.[121]
In 2018, a study compared sequences of North American dog fossils with Siberian dog fossils and modern dogs. The nearest relative to the North American fossils was a 9,000 YBP fossil discovered on Zhokhov Island, arctic north-eastern Siberia, which was connected to the mainland at that time. The study inferred from mDNA that all of the North American dogs shared a common ancestor dated 14,600 YBP, and this ancestor had diverged along with the ancestor of the Zhokhov dog from their common ancestor 15,600 YBP. The timing of the Koster dogs shows that dogs entered North America from Siberia 4,500 years after humans did, were isolated for the next 9,000 years, and after contact with Europeans these no longer exist because they were replaced by Eurasian dogs. The (earlier wave of, prior to the Thule/Inuit introduction) pre-contact dogs exhibit a unique genetic signature that is now gone, with nDNA indicating that their nearest genetic relatives today are the arctic breed dogs: Alaskan malamutes, Greenland dogs, and Alaskan huskies and Siberian huskies.[122]
In 2019, a study found that those dogs brought initially into the North American Arctic from northeastern Siberia were later replaced by dogs accompanying the Inuit during their expansion beginning 2,000 years ago. These Inuit dogs were more genetically diverse and more morphologically divergent when compared with the earlier dogs. Today, Arctic sledge dogs are among the last descendants in the Americas of this pre-European dog lineage.[123] In 2020, the sequencing of ancient dog genomes indicates that in two Mexican breeds the Chihuahua retains 4% and the Xoloitzcuintli 3% pre-colonial ancestry.[3]
Late Pleistocene wolf admixture

In 2015, a study mapped the first genome of a 35,000 YBP Pleistocene wolf fossil found in the Taimyr Peninsula, arctic northern Siberia and compared it with those of modern dogs and grey wolves. The Taimyr wolf was identified through mDNA as Canis lupus but from a population which had diverged from the dog–grey wolf lineage immediately before the dog and grey wolf diverged from each other, which implies that the majority of grey wolf populations today stems from an ancestral population that lived less than 35,000 years ago but before the inundation of the Bering Land Bridge with the subsequent isolation of Eurasian and North American wolves.[124]
The Taimyr wolf was equally related to both dogs and modern wolves, but shared more alleles (i.e. gene expressions) with those breeds that are associated with high latitudes and arctic human populations: the Siberian husky and Greenland dog, and to a lesser extent the Shar Pei and Finnish spitz. The Greenland dog shows 3.5% Taimyr wolf ancestry, which indicates admixture between the Taimyr wolf population and the ancestral dog population of these four high-latitude breeds. These results can be explained either by a very early presence of dogs in northern Eurasia or by the genetic legacy of the Taimyr wolf being preserved in northern wolf populations until the arrival of dogs into high latitudes. This introgression could have provided early dogs living in high latitudes with adaptations to the new and challenging environment. It also indicates that the ancestry of present-day dog breeds descends from more than one region.[124]: 3–4 An attempt to explore admixture between the Taimyr wolf and grey wolves produced unreliable results.[124]: 23
As the Taimyr wolf had contributed to the genetic makeup of the Arctic breeds, this indicates that the descendants of the Taimyr wolf survived until dogs were domesticated in Europe and arrived at high latitudes where they mixed with local wolves, and these both contributed to the modern Arctic breeds. Based on the most widely accepted oldest zooarchaeological dog remains, domestic dogs most likely arrived at high latitudes within the last 15,000 years.[1]
The nuclear genome sequence was generated for a dog specimen that was found in the Late Neolithic passage grave at Newgrange, Ireland and radiocarbon dated at 4,800 YBP. A genetic analysis of the Newgrange dog showed that it was male, did not possess genetic variants associated with modern coat length nor color, was not as able to process starch as efficiently as modern dogs but more efficiently than wolves, and showed ancestry from a population of wolves that could not be found in other dogs nor wolves today.[49] The mutation rates calibrated from both the Taimyr wolf and the Newgrange dog genomes suggest that the modern wolf and dog populations diverged from a common ancestor between 20,000 and 60,000 YBP. This indicates that either dogs were domesticated much earlier than their first appearance in the archaeological record, or they arrived in the Arctic early, or both.[10] Another view is that because northern breeds can trace at least some of their ancestry back to the Taimyr wolf, this indicates the possibility of more than one domestication event.[1]
In 2020, the nuclear genome was generated of a 33,000 YBP Pleistocene wolf from an archaeological site on the Yana River, arctic northeastern Siberia. The Yana wolf sequence was more closely related to the 35,000 YBP Taimyr wolf than it was to modern wolves. There was evidence of gene flow between the Yana-Taimyr wolves and the Pre-Columbian, Zhokhov, and modern sled dogs. This suggests that genetic admixture has occurred between the Pleistocene wolves and the ancestor of these dogs. There was no evidence of admixture between sled dogs and the modern grey wolf for the past 9,500 years. Greenland sled dogs have been kept isolated from other breeds since their arrival in Greenland with the Inuit 850 years ago. Their lineage traces more genomic history to the Zhokhov dogs than any other arctic breed. Sled dogs do not show an adaptation to a starch-rich diet when compared with other dogs but do show an adaptation to a high intake of fat and fatty acids, which was not found in the Zhokhov dogs. The same adaptation has been found in Inuit and other arctic peoples. This suggests that the sled dogs adapted to the low starch and high fat diet of the people they coexisted with.[118]
In 2021, a study of another 4 Late Pleistocene northeast Siberian wolf sequences showed that they are genetically similar to the Taimyr and Yana wolves. These 6 extinct wolves sequentially branched off from the lineage that leads to the modern wolf and dog. The 50,000 YBP Tirekhtyakh River, 48,000 YBP Bunge-Toll site, and 32,000 YBP Yana RHS specimens were separate lineages not related to each other. The 16,800 YBP Ulakhan Sular and the 14,100 YBP Tumat specimens both cluster with a modern wolf from Ellesmere Island, indicating that these 2 specimens derive from the same lineage as the North American wolves. All 6 Late Pleistocene wolves share alleles with Arctic dogs: Greenland dogs, Siberian and Alaskan huskies, Alaskan malamutes, the extinct Zhokhov dog and the extinct pre-European contact dogs of North America. It is possible that another population of extinct wolves, that were related to all six specimens, may have contributed to the ancestry of the Arctic dogs. There was evidence that 4 of the extinct Siberian wolves had contributed to the ancestry of modern wolf populations in Shanxi, western China, and possibly Chukotka and Inner Mongolia.[125]
Dogs enter Japan
The oldest fossil of a dog that has been found in Japan dates to 9,500 YBP.[126] With the beginning of the Holocene and its warmer weather, temperate deciduous forests rapidly spread onto the main island of Honshu and caused an adaption away from hunting megafauna (Naumann's elephant and Yabe's giant deer) to hunting the quicker sika deer and wild boar in dense forest. With this came a change in hunting technology, including a shift to smaller, triangular points for arrows. A study of the Jōmon people that lived on the Pacific coast of Honshu during the early Holocene shows that they were conducting individual dog burials and were probably using dogs as tools for hunting sika deer and wild boar, as hunters in Japan still do today.[116]
Hunting dogs make major contributions to forager societies and the ethnographic record shows them being given proper names, treated as family members, and considered separate to other types of dogs.[116][127] This special treatment includes separate burials with markers and grave-goods,[116][128][129] with those that were exceptional hunters or that were killed on the hunt often venerated.[116][130] A dog's value as a hunting partner gives them status as a living weapon and the most skilled elevated to taking on a "personhood", with their social position in life and in death similar to that of the skilled hunters.[116][131]
Intentional dog burials together with ungulate hunting is also found in other early Holocene deciduous forest forager societies in Europe[132] and North America,[133][134] indicating that across the Holarctic temperate zone hunting dogs were a widespread adaptation to forest ungulate hunting.[116]
Dogs from the Near East enter Africa

In 2020, the sequencing of ancient dog genomes indicates that the lineage of modern dogs in sub-Sahara Africa shares a single origin from the Levant, where an ancestral specimen was dated to 7,000 YBP. This finding mirrors the gene flow of humans from the Levant into Africa during the Neolithic, along with cattle. Since then, there has been limited gene flow into African dogs until the past few hundred years. The descendants of a dog from Iran dated 5,800 YBP and dogs from Europe completely replaced the Levant dog lineage 2,300 YBP. This was associated with human migration from Iran and some minor migration from Europe. Today all Near Eastern dogs show 81% ancient Iranian and 19% Neolithic European ancestry.[3]
The oldest dog remains to be found in Africa date 5,900 YBP and were discovered at the Merimde Beni-Salame Neolithic site in the Nile Delta, Egypt. The next oldest remains date 5,500 YBP and were found at Esh Shareinab on the Nile in Sudan. This suggests that the dog arrived from Asia at the same time as domestic sheep and goats.[135] The dog then spread north to south down Africa beside livestock herders, with remains found in archaeological sites dated 925–1,055 YBP at Ntusi in Uganda, dated 950–1,000 YBP at Kalomo in Zambia, and then at sites south of the Limpopo River and into southern Africa.[136] In 2020, the sequencing of ancient dog genomes indicates that the southern African Rhodesian Ridgeback retains 4% pre-colonial ancestry.[3]
Dogs enter South East Asia and Oceania from southern China
In 2020, an mDNA study of ancient dog fossils from the Yellow River and Yangtze River basins of southern China showed that most of the ancient dogs fell within haplogroup A1b, as do the Australian dingoes and the pre-colonial dogs of the Pacific, but in low frequency in China today. The specimen from the Tianluoshan archaeological site, Zhejiang province dates to 7,000 YBP and is basal to the entire lineage. The dogs belonging to this haplogroup were once widely distributed in southern China, then dispersed through Southeast Asia into New Guinea and Oceania, but were replaced in China 2,000 YBP by dogs of other lineages.[137]
See also
References
- Thalmann, Olaf; Perri, Angela R. (2018). "Paleogenomic Inferences of Dog Domestication". In Lindqvist, C.; Rajora, O. (eds.). Paleogenomics. Population Genomics. Springer, Cham. pp. 273–306. doi:10.1007/13836_2018_27. ISBN 978-3-030-04752-8.
- Irving-Pease, Evan K.; Ryan, Hannah; Jamieson, Alexandra; Dimopoulos, Evangelos A.; Larson, Greger; Frantz, Laurent A. F. (2018). "Paleogenomics of Animal Domestication". In Lindqvist, C.; Rajora, O. (eds.). Paleogenomics. Population Genomics. Springer, Cham. pp. 225–272. doi:10.1007/13836_2018_55. ISBN 978-3-030-04752-8. Archived from the original on 8 December 2019. Retrieved 4 December 2019.
- Bergström, Anders; Frantz, Laurent; Schmidt, Ryan; Ersmark, Erik; Lebrasseur, Ophelie; Girdland-Flink, Linus; Lin, Audrey T.; Storå, Jan; Sjögren, Karl-Göran; Anthony, David; Antipina, Ekaterina; Amiri, Sarieh; Bar-Oz, Guy; Bazaliiskii, Vladimir I.; Bulatović, Jelena; Brown, Dorcas; Carmagnini, Alberto; Davy, Tom; Fedorov, Sergey; Fiore, Ivana; Fulton, Deirdre; Germonpré, Mietje; Haile, James; Irving-Pease, Evan K.; Jamieson, Alexandra; Janssens, Luc; Kirillova, Irina; Horwitz, Liora Kolska; Kuzmanovic-Cvetković, Julka; Kuzmin, Yaroslav; Losey, Robert J.; Dizdar, Daria Ložnjak; Mashkour, Marjan; Novak, Mario; Onar, Vedat; Orton, David; Pasaric, Maja; Radivojevic, Miljana; Rajkovic, Dragana; Roberts, Benjamin; Ryan, Hannah; Sablin, Mikhail; Shidlovskiy, Fedor; Stojanovic, Ivana; Tagliacozzo, Antonio; Trantalidou, Katerina; Ullén, Inga; Villaluenga, Aritza; Wapnish, Paula; Dobney, Keith; Götherström, Anders; Linderholm, Anna; Dalén, Love; Pinhasi, Ron; Larson, Greger; Skoglund, Pontus (2020). "Origins and genetic legacy of prehistoric dogs". Science. 370 (6516): 557–564. doi:10.1126/science.aba9572. PMC 7116352. PMID 33122379. S2CID 225956269.
- Frantz, Laurent A. F.; Bradley, Daniel G.; Larson, Greger; Orlando, Ludovic (2020). "Animal domestication in the era of ancient genomics". Nature Reviews Genetics. 21 (8): 449–460. doi:10.1038/s41576-020-0225-0. PMID 32265525. S2CID 214809393. Archived from the original on 3 October 2020. Retrieved 15 September 2020.
- Freedman, Adam H; Wayne, Robert K (2017). "Deciphering the Origin of Dogs: From Fossils to Genomes". Annual Review of Animal Biosciences. 5 (1): 281–307. doi:10.1146/annurev-animal-022114-110937. PMID 27912242.
- Lord, Kathryn A.; Larson, Greger; Coppinger, Raymond P.; Karlsson, Elinor K. (2020). "The History of Farm Foxes Undermines the Animal Domestication Syndrome". Trends in Ecology & Evolution. 35 (2): 125–136. doi:10.1016/j.tree.2019.10.011. PMID 31810775.
- Wayne, Robert K. (June 1993). "Molecular evolution of the dog family". Trends in Genetics. 9 (6): 218–224. doi:10.1016/0168-9525(93)90122-x. PMID 8337763. Archived from the original on 24 February 2021. Retrieved 16 February 2021.
- Lindblad-Toh, Kerstin; Wade, Claire M.; Mikkelsen, Tarjei S.; Karlsson, Elinor K.; Jaffe, David B.; Kamal, Michael; et al. (2005). "Genome sequence, comparative analysis and haplotype structure of the domestic dog". Nature. 438 (7069): 803–819. Bibcode:2005Natur.438..803L. doi:10.1038/nature04338. PMID 16341006.
- Castelló, J.R. (2018). "Ch.2 - Wolf-like Canids". Canids of the World: Wolves, Wild Dogs, Foxes, Jackals, Coyotes, and Their Relatives. Princeton University Press. p. 74. ISBN 978-0-691-18372-5. Archived from the original on 15 April 2021. Retrieved 8 October 2022.
- Machugh, David E.; Larson, Greger; Orlando, Ludovic (2016). "Taming the Past: Ancient DNA and the Study of Animal Domestication". Annual Review of Animal Biosciences. 5: 329–351. doi:10.1146/annurev-animal-022516-022747. PMID 27813680.
- Larson, G.; Bradley, D.G. (2014). "How Much Is That in Dog Years? The Advent of Canine Population Genomics". PLOS Genetics. 10 (1): e1004093. doi:10.1371/journal.pgen.1004093. PMC 3894154. PMID 24453989.
- Nesse, Randolph M. (2007). "Runaway social selection for displays of partner value and altruism". Biological Theory. Springer Science+Business Media. 2 (2): 146. doi:10.1162/biot.2007.2.2.143. S2CID 195097363.
- Perri, Angela R.; Feuerborn, Tatiana R.; Frantz, Laurent A. F.; Larson, Greger; Malhi, Ripan S.; Meltzer, David J.; Witt, Kelsey E. (2021). "Dog domestication and the dual dispersal of people and dogs into the Americas". Proceedings of the National Academy of Sciences. 118 (6): e2010083118. Bibcode:2021PNAS..11810083P. doi:10.1073/pnas.2010083118. PMC 8017920. PMID 33495362. Archived from the original on 15 February 2021. Retrieved 7 February 2021.
Advances in the isolation and sequencing of ancient DNA [... suggest] that dogs were domesticated in Siberia by ∼23,000 y ago, possibly while both people and wolves were isolated during the harsh climate of the Last Glacial Maximum.
- "Ancient Domesticated Dog Skull Found in Siberian Cave". 17 January 2012. Archived from the original on 31 March 2022. Retrieved 24 June 2022.
- Nowak, R.M. (2003). "Chapter 9 - Wolf evolution and taxonomy". In Mech, L. David; Boitani, Luigi (eds.). Wolves: Behaviour, Ecology and Conservation. University of Chicago Press. pp. 239–258. ISBN 978-0-226-51696-7. page 239
- Ostrander, Elaine A.; Wang, Guo-Dong; Larson, Greger; Vonholdt, Bridgett M.; Davis, Brian W.; Jagannathan, Vidyha; Hitte, Christophe; Wayne, Robert K.; Zhang, Ya-Ping (2019). "Dog10K: An international sequencing effort to advance studies of canine domestication, phenotypes, and health". National Science Review. 6 (4): 810–824. doi:10.1093/nsr/nwz049. PMC 6776107. PMID 31598383.
- Schweizer, Rena M.; Wayne, Robert K. (2020). "Illuminating the mysteries of wolf history". Molecular Ecology. 29 (9): 1589–1591. doi:10.1111/MEC.15438. PMID 32286714.
- Cooper, A. (2015). "Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover". Science. 349 (6248): 602–6. Bibcode:2015Sci...349..602C. doi:10.1126/science.aac4315. PMID 26250679. S2CID 31686497.
- Pilot, Małgorzata; Moura, Andre E.; Okhlopkov, Innokentiy M.; Mamaev, Nikolay V.; Alagaili, Abdulaziz N.; Mohammed, Osama B.; Yavruyan, Eduard G.; Manaseryan, Ninna H.; Hayrapetyan, Vahram; Kopaliani, Natia; Tsingarska, Elena; Krofel, Miha; Skoglund, Pontus; Bogdanowicz, Wiesław (2019). "Global Phylogeographic and Admixture Patterns in Grey Wolves and Genetic Legacy of an Ancient Siberian Lineage". Scientific Reports. 9 (1): 17328. Bibcode:2019NatSR...917328P. doi:10.1038/s41598-019-53492-9. PMC 6874602. PMID 31757998.
- Thalmann, O; Shapiro, B; Cui, P; Schuenemann, V. J; Sawyer, S. K; Greenfield, D. L; Germonpre, M. B; Sablin, M. V; Lopez-Giraldez, F; Domingo-Roura, X; Napierala, H; Uerpmann, H.-P; Loponte, D. M; Acosta, A. A; Giemsch, L; Schmitz, R. W; Worthington, B; Buikstra, J. E; Druzhkova, A; Graphodatsky, A. S; Ovodov, N. D; Wahlberg, N; Freedman, A. H; Schweizer, R. M; Koepfli, K.- P; Leonard, J. A; Meyer, M; Krause, J; Paabo, S; et al. (2013). "Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs". Science. 342 (6160): 871–4. Bibcode:2013Sci...342..871T. doi:10.1126/science.1243650. hdl:10261/88173. PMID 24233726. S2CID 1526260.
- Koblmüller, Stephan; Vilà, Carles; Lorente-Galdos, Belen; Dabad, Marc; Ramirez, Oscar; Marques-Bonet, Tomas; Wayne, Robert K.; Leonard, Jennifer A. (2016). "Whole mitochondrial genomes illuminate ancient intercontinental dispersals of grey wolves (Canis lupus)". Journal of Biogeography. 43 (9): 1728–1738. doi:10.1111/jbi.12765. hdl:10261/153364. S2CID 88740690.
- Loog, Liisa; Thalmann, Olaf; Sinding, Mikkel‐Holger S.; Schuenemann, Verena J.; Perri, Angela; Germonpré, Mietje; Bocherens, Herve; Witt, Kelsey E.; Samaniego Castruita, Jose A.; Velasco, Marcela S.; Lundstrøm, Inge K.C.; Wales, Nathan; Sonet, Gontran; Frantz, Laurent; Schroeder, Hannes; Budd, Jane; Jimenez, Elodie‐Laure; Fedorov, Sergey; Gasparyan, Boris; Kandel, Andrew W.; Lázničková‐Galetová, Martina; Napierala, Hannes; Uerpmann, Hans‐Peter; Nikolskiy, Pavel A.; Pavlova, Elena Y.; Pitulko, Vladimir V.; Herzig, Karl‐Heinz; Malhi, Ripan S.; Willerslev, Eske; et al. (2019). "Ancient DNA suggests modern wolves trace their origin to a late Pleistocene expansion from Beringia". Molecular Ecology. 29 (9): 1596–1610. doi:10.1111/mec.15329. PMC 7317801. PMID 31840921.
- Werhahn, Geraldine; Senn, Helen; Ghazali, Muhammad; Karmacharya, Dibesh; Sherchan, Adarsh Man; Joshi, Jyoti; Kusi, Naresh; López-Bao, José Vincente; Rosen, Tanya; Kachel, Shannon; Sillero-Zubiri, Claudio; MacDonald, David W. (2018). "The unique genetic adaptation of the Himalayan wolf to high-altitudes and consequences for conservation". Global Ecology and Conservation. 16: e00455. doi:10.1016/j.gecco.2018.e00455.
- Morey, Darcy F.; Jeger, Rujana (2016). "From wolf to dog: Late Pleistocene ecological dynamics, altered trophic strategies, and shifting human perceptions". Historical Biology. 29 (7): 895–903. doi:10.1080/08912963.2016.1262854. S2CID 90588969.
- Morrell, Virginia (2015). "Ch2.1-From Wolf to Dog". Our Furry Friends: The Science of Pets. Scientific American. pp. 44–46. ISBN 978-1-4668-5901-2. Archived from the original on 8 October 2022. Retrieved 16 October 2021.
- Freedman, Adam H.; Schweizer, Rena M.; Ortega-Del Vecchyo, Diego; Han, Eunjung; Davis, Brian W.; Gronau, Ilan; Silva, Pedro M.; Galaverni, Marco; Fan, Zhenxin; Marx, Peter; Lorente-Galdos, Belen; Ramirez, Oscar; Hormozdiari, Farhad; Alkan, Can; Vilà, Carles; Squire, Kevin; Geffen, Eli; Kusak, Josip; Boyko, Adam R.; Parker, Heidi G.; Lee, Clarence; Tadigotla, Vasisht; Siepel, Adam; Bustamante, Carlos D.; Harkins, Timothy T.; Nelson, Stanley F.; Marques-Bonet, Tomas; Ostrander, Elaine A.; Wayne, Robert K.; Novembre, John (2016). "Demographically-Based Evaluation of Genomic Regions under Selection in Domestic Dogs". PLOS Genetics. 12 (3): e1005851. doi:10.1371/journal.pgen.1005851. PMC 4778760. PMID 26943675.
- Botigué, Laura R.; Song, Shiya; Scheu, Amelie; Gopalan, Shyamalika; Pendleton, Amanda L.; Oetjens, Matthew; Taravella, Angela M.; Seregély, Timo; Zeeb-Lanz, Andrea; Arbogast, Rose-Marie; Bobo, Dean; Daly, Kevin; Unterländer, Martina; Burger, Joachim; Kidd, Jeffrey M.; Veeramah, Krishna R. (2017). "Ancient European dog genomes reveal continuity since the Early Neolithic". Nature Communications. 8: 16082. Bibcode:2017NatCo...816082B. doi:10.1038/ncomms16082. PMC 5520058. PMID 28719574.
- Frantz, L. A. F.; Mullin, V. E.; Pionnier-Capitan, M.; Lebrasseur, O.; Ollivier, M.; Perri, A.; Linderholm, A.; Mattiangeli, V.; Teasdale, M. D.; Dimopoulos, E. A.; Tresset, A.; Duffraisse, M.; McCormick, F.; Bartosiewicz, L.; Gal, E.; Nyerges, E. A.; Sablin, M. V.; Brehard, S.; Mashkour, M.; b l Escu, A.; Gillet, B.; Hughes, S.; Chassaing, O.; Hitte, C.; Vigne, J.-D.; Dobney, K.; Hanni, C.; Bradley, D. G.; Larson, G. (2016). "Genomic and archaeological evidence suggest a dual origin of domestic dogs". Science. 352 (6290): 1228–31. Bibcode:2016Sci...352.1228F. doi:10.1126/science.aaf3161. PMID 27257259. S2CID 206647686. Archived from the original on 4 July 2017. Retrieved 22 February 2019.
- Schleidt, Wolfgang M.; Shalter, Michael D. (2018). "Dogs and Mankind: Coevolution on the Move – an Update". Human Ethology Bulletin. 33: 15–38. doi:10.22330/heb/331/015-038.
- Silva, Pedro; Galaverni, Marco; Ortega-Del Vecchyo, Diego; Fan, Zhenxin; Caniglia, Romolo; Fabbri, Elena; Randi, Ettore; Wayne, Robert; Godinho, Raquel (2020). "Genomic evidence for the Old divergence of Southern European wolf populations". Proceedings of the Royal Society B: Biological Sciences. 287 (1931). doi:10.1098/rspb.2020.1206. PMC 7423677. PMID 32693716.
- Savolainen, P. (2002). "Genetic evidence for an East Asian origin of domestic dogs". Science. 298 (5598): 1610–3. Bibcode:2002Sci...298.1610S. doi:10.1126/science.1073906. PMID 12446907. S2CID 32583311.
- Pang, J. (2009). "mtDNA data indicate a single origin for dogs south of Yangtze River, less than 16,300 years ago, from numerous wolves". Molecular Biology and Evolution. 26 (12): 2849–64. doi:10.1093/molbev/msp195. PMC 2775109. PMID 19723671.
- Ardalan, A (2011). "Comprehensive study of mtDNA among Southwest Asian dogs contradicts independent domestication of wolf, but implies dog–wolf hybridization". Ecology and Evolution. 1 (3): 373–385. doi:10.1002/ece3.35. PMC 3287314. PMID 22393507.
- Brown, Sarah K.; Pedersen, Niels C.; Jafarishorijeh, Sardar; Bannasch, Danika L.; Ahrens, Kristen D.; Wu, Jui-Te; Okon, Michaella; Sacks, Benjamin N. (2011). "Phylogenetic Distinctiveness of Middle Eastern and Southeast Asian Village Dog y Chromosomes Illuminates Dog Origins". PLOS ONE. 6 (12): e28496. Bibcode:2011PLoSO...628496B. doi:10.1371/journal.pone.0028496. PMC 3237445. PMID 22194840.
- Ding, Z. (2011). "Origins of domestic dog in Southern East Asia is supported by analysis of Y-chromosome DNA". Heredity. 108 (5): 507–14. doi:10.1038/hdy.2011.114. PMC 3330686. PMID 22108628.
- Wang, Guo-Dong; Zhai, Weiwei; Yang, He-Chuan; Wang, Lu; Zhong, Li; Liu, Yan-Hu; Fan, Ruo-Xi; Yin, Ting-Ting; Zhu, Chun-Ling; Poyarkov, Andrei D.; Irwin, David M.; Hytönen, Marjo K.; Lohi, Hannes; Wu, Chung-I; Savolainen, Peter; Zhang, Ya-Ping (2016). "Out of southern East Asia: The natural history of domestic dogs across the world". Cell Research. 26 (1): 21–33. doi:10.1038/cr.2015.147. PMC 4816135. PMID 26667385.
- Boyko, A. (2009). "Complex population structure in African village dogs and its implications for inferring dog domestication history". Proceedings of the National Academy of Sciences. 106 (33): 13903–13908. Bibcode:2009PNAS..10613903B. doi:10.1073/pnas.0902129106. PMC 2728993. PMID 19666600.
- Larson G (2012). "Rethinking dog domestication by integrating genetics, archeology, and biogeography". PNAS. 109 (23): 8878–8883. Bibcode:2012PNAS..109.8878L. doi:10.1073/pnas.1203005109. PMC 3384140. PMID 22615366.
- Shannon, Laura M.; Boyko, Ryan H.; Castelhano, Marta; Corey, Elizabeth; Hayward, Jessica J.; McLean, Corin; White, Michelle E.; Abi Said, Mounir; Anita, Baddley A.; Bondjengo, Nono Ikombe; Calero, Jorge; Galov, Ana; Hedimbi, Marius; Imam, Bulu; Khalap, Rajashree; Lally, Douglas; Masta, Andrew; Oliveira, Kyle C.; Pérez, Lucía; Randall, Julia; Tam, Nguyen Minh; Trujillo-Cornejo, Francisco J.; Valeriano, Carlos; Sutter, Nathan B.; Todhunter, Rory J.; Bustamante, Carlos D.; Boyko, Adam R. (2015). "Genetic structure in village dogs reveals a Central Asian domestication origin". Proceedings of the National Academy of Sciences. 112 (44): 13639–13644. Bibcode:2015PNAS..11213639S. doi:10.1073/pnas.1516215112. PMC 4640804. PMID 26483491.
- vonHoldt, B. (2010). "Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication". Nature. 464 (7290): 898–902. Bibcode:2010Natur.464..898V. doi:10.1038/nature08837. PMC 3494089. PMID 20237475.
- Freedman, A. (2014). "Genome sequencing highlights the dynamic early history of dogs". PLOS Genetics. 10 (1): e1004016. doi:10.1371/journal.pgen.1004016. PMC 3894170. PMID 24453982.
- Wayne, Robert K.; Vonholdt, Bridgett M. (2012). "Evolutionary genomics of dog domestication". Mammalian Genome. 23 (1–2): 3–18. doi:10.1007/s00335-011-9386-7. PMID 22270221. S2CID 16003335.
- Lee, E. (2015). "Ancient DNA analysis of the oldest canid species from the Siberian Arctic and genetic contribution to the domestic dog". PLOS ONE. 10 (5): e0125759. Bibcode:2015PLoSO..1025759L. doi:10.1371/journal.pone.0125759. PMC 4446326. PMID 26018528.
- Irizarry, Kristopher J. L.; Vasconcelos, Elton J. R. (2018). "Population Genomics of Domestication and Breed Development in Canines in the Context of Cognitive, Social, Behavioral, and Disease Traits". In Rajora, O. (ed.). Population Genomics. pp. 755–806. doi:10.1007/13836_2018_43. ISBN 978-3-030-04587-6.
- Druzhkova, Anna S.; Thalmann, Olaf; Trifonov, Vladimir A.; Leonard, Jennifer A.; Vorobieva, Nadezhda V.; Ovodov, Nikolai D.; Graphodatsky, Alexander S.; Wayne, Robert K. (2013). "Ancient DNA Analysis Affirms the Canid from Altai as a Primitive Dog". PLOS ONE. 8 (3): e57754. Bibcode:2013PLoSO...857754D. doi:10.1371/journal.pone.0057754. PMC 3590291. PMID 23483925.
- Miklosi, Adam (2018). "1-Evolution & Ecology". The Dog: A Natural History. Princeton University Press. pp. 13–39. ISBN 978-0-691-17693-2. Archived from the original on 8 October 2022. Retrieved 17 March 2018.
- Vila, C. (1997). "Multiple and ancient origins of the domestic dog". Science. 276 (5319): 1687–9. doi:10.1126/science.276.5319.1687. PMID 9180076.
- Shipman 2015, pp. 149
- Frantz, L. (2015). "Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes". Nature Genetics. 47 (10): 1141–1148. doi:10.1038/ng.3394. PMID 26323058. S2CID 205350534.
- Schnitzler, Annick; Patou-Mathis, Marylène (2017). "Wolf (Canis lupus Linnaeus, 1758) domestication: Why did it occur so late and at such high latitude? A hypothesis". Anthropozoologica. 52 (2): 149. doi:10.5252/az2017n2a1. S2CID 134153466. Archived from the original on 14 April 2021. Retrieved 7 February 2021.
- Ciucani, Marta Maria; Palumbo, Davide; Galaverni, Marco; Serventi, Patrizia; Fabbri, Elena; Ravegnini, Gloria; Angelini, Sabrina; Maini, Elena; Persico, Davide; Caniglia, Romolo; Cilli, Elisabetta (2019). "Old wild wolves: Ancient DNA survey unveils population dynamics in Late Pleistocene and Holocene Italian remains". PeerJ. 7: e6424. doi:10.7717/peerj.6424. PMC 6441319. PMID 30944772.
- Koupadi, Kyriaki; Fontani, Francesco; Ciucani, Marta Maria; Maini, Elena; De Fanti, Sara; Cattani, Maurizio; Curci, Antonio; Nenzioni, Gabriele; Reggiani, Paolo; Andrews, Adam J.; Sarno, Stefania; Bini, Carla; Pelotti, Susi; Caniglia, Romolo; Luiselli, Donata; Cilli, Elisabetta (2020). "Population Dynamics in Italian Canids between the Late Pleistocene and Bronze Age". Genes. 11 (12): 1409. doi:10.3390/genes11121409. PMC 7761486. PMID 33256122.
- Boschin, Francesco; Bernardini, Federico; Pilli, Elena; Vai, Stefania; Zanolli, Clément; Tagliacozzo, Antonio; Fico, Rosario; Fedi, Mariaelena; Corny, Julien; Dreossi, Diego; Lari, Martina; Modi, Alessandra; Vergata, Chiara; Tuniz, Claudio; Moroni, Adriana; Boscato, Paolo; Caramelli, David; Ronchitelli, Annamaria (2020). "The first evidence for Late Pleistocene dogs in Italy". Scientific Reports. 10 (1): 13313. Bibcode:2020NatSR..1013313B. doi:10.1038/s41598-020-69940-w. PMC 7414845. PMID 32770100. S2CID 221019303.
- Bergström, A., Stanton, D.W.G., Taron, U.H. et al. Grey wolf genomic history reveals a dual ancestry of dogs. Nature 607, 313–320 (2022). https://doi.org/10.1038/s41586-022-04824-9 Archived 8 October 2022 at the Wayback Machine
- Ed Yong (2016). "A New Origin Story for Dogs - Interview with Greger Larson". The Atlantic Monthly Group. Archived from the original on 30 March 2017. Retrieved 10 March 2017.
- Grimm, David (2015). "Feature: Solving the mystery of dog domestication". Science. doi:10.1126/science.aab2477. quoting Greger Larson
- Pierotti & Fogg 2017, pp. 192–193
- Jans, N. (2014). A wolf called Romeo. Houghton Mifflin Harcourt. ISBN 978-0-547-85819-7. Archived from the original on 8 October 2022. Retrieved 5 July 2018.
- Derr 2011, pp. 85–98
- Arctic Wolf: The High Arctic by Laura DeLallo. Bearport Publishing, New York 2011
- Arctic wildlife in a warming world Archived 2 January 2015 at the Wayback Machine by Michael Becker. BBC Two, 2014.
- Ellesmere Island Journal & Field Notes Archived 19 October 2016 at the Wayback Machine by Henry Beston 2006. International Wolf Centre.
- Arctic Wolves and Their Prey Archived 19 October 2016 at the Wayback Machine by L. David Mech. National Ocean and Atmospheric Administration, Pacific Marine Environment Laboratory, Arctic Zone. 2004
- Wang, Xiaoming; Tedford, Richard H.; Dogs: Their Fossil Relatives and Evolutionary History. New York: Columbia University Press, 2008. pp. 166
- Schleidt, W. (2003). "Co-evolution of humans and canids: An alternative view of dog domestication: Homo homini lupus?" (PDF). Evolution and Cognition. 9 (1): 57–72. Archived (PDF) from the original on 8 October 2022. Retrieved 15 December 2016.
- Frans de Waal (2006). Primates and Philosophers: How Morality Evolved. Princeton University Press. p. 3. ISBN 978-0-691-12447-6.
- Olsen, S. J. (1985). Origins of the domestic dog: the fossil record. Univ. of Arizona Press, Tucson, US. pp. 88–89.
- Zeder MA (2012). "The domestication of animals". Journal of Anthropological Research. 68 (2): 161–190. doi:10.3998/jar.0521004.0068.201. S2CID 85348232.
- Schleidt, W. M. (1998). "Is humaneness canine?". Human Ethology Bulletin. 13 (4): 1–4.
- Andrew Brown Smith (2005). African Herders: Emergence of Pastoral Traditions. Walnut Creek: Altamira Press. p. 27.
- Musiani M, Leonard JA, Cluff H, Gates CC, Mariani S, et al. (2007). "Differentiation of tundra/taiga and boreal coniferous forest wolves: genetics, coat colour and association with migratory caribou". Mol. Ecol. 16 (19): 4149–70. doi:10.1111/j.1365-294x.2007.03458.x. PMID 17725575. S2CID 14459019.
- Leonard, J. (2007). "Megafaunal extinctions and the disappearance of a specialized wolf ecomorph". Current Biology. 17 (13): 1146–50. doi:10.1016/j.cub.2007.05.072. hdl:10261/61282. PMID 17583509. S2CID 14039133.
- Lahtinen, Maria; Clinnick, David; Mannermaa, Kristiina; Salonen, J. Sakari; Viranta, Suvi (2021). "Excess protein enabled dog domestication during severe Ice Age winters". Scientific Reports. 11 (1): 7. Bibcode:2021NatSR..11....7L. doi:10.1038/s41598-020-78214-4. PMC 7790815. PMID 33414490.
 Text was copied/adapted from this source, which is available under a Creative Commons Attribution 4.0 International License Archived 16 October 2017 at the Wayback Machine.
Text was copied/adapted from this source, which is available under a Creative Commons Attribution 4.0 International License Archived 16 October 2017 at the Wayback Machine. - Bannasch, Danika L.; et al. (2021). "Dog colour patterns explained by modular promoters of ancient canid origin". Nature Ecology & Evolution. 5 (10): 1415–1423. doi:10.1038/s41559-021-01524-x. PMC 8484016. PMID 34385618.
- Shipman, Pat (2015). "How do you kill 86 mammoths? Taphonomic investigations of mammoth megasites". Quaternary International. 359–360: 38–46. Bibcode:2015QuInt.359...38S. doi:10.1016/j.quaint.2014.04.048.
- Zimov, S.A.; Zimov, N.S.; Tikhonov, A.N.; Chapin, F.S. (2012). "Mammoth steppe: A high-productivity phenomenon". Quaternary Science Reviews. 57: 26–45. Bibcode:2012QSRv...57...26Z. doi:10.1016/j.quascirev.2012.10.005. S2CID 14078430.
- Li, Y. (2014). "Domestication of the dog from the wolf was promoted by enhanced excitatory synaptic plasticity: A hypothesis". Genome Biology and Evolution. 6 (11): 3115–21. doi:10.1093/gbe/evu245. PMC 4255776. PMID 25377939.
- Serpell J, Duffy D. Dog Breeds and Their Behavior. In: Domestic Dog Cognition and Behavior. Berlin, Heidelberg: Springer; 2014
- Cagan, Alex; Blass, Torsten (2016). "Identification of genomic variants putatively targeted by selection during dog domestication". BMC Evolutionary Biology. 16: 10. doi:10.1186/s12862-015-0579-7. PMC 4710014. PMID 26754411.
- Almada RC, Coimbra NC. Recruitment of striatonigral disinhibitory and nigrotectal inhibitory GABAergic pathways during the organization of defensive behavior by mice in a dangerous environment with the venomous snake Bothrops alternatus [ Reptilia, Viperidae ] Synapse 2015:n/a–n/a
- Coppinger R, Schneider R: Evolution of working dogs. The domestic dog: Its evolution, behaviour and interactions with people. Cambridge: Cambridge University press, 1995
- Cimarelli, Giulia; Virányi, Zsófia (2019). "Social Behaviour of Pet Dogs is Associated with Peripheral OXTR Methylation". In Topál, József; Kis, Anna; Oliva, Jessica; Virányi, Zsófia (eds.). Oxytocin and Social Behaviour in Dogs and Other (Self-)Domesticated Species Methodological Caveats and Promising Perspectives. Frontiers Media. pp. 52–53. ISBN 978-2-88945-913-1. Archived from the original on 15 August 2021. Retrieved 8 October 2022.
- Sundman, Ann-Sofie; Pértille, Fábio; Lehmann Coutinho, Luiz; Jazin, Elena; Guerrero-Bosagna, Carlos; Jensen, Per (2020). "DNA methylation in canine brains is related to domestication and dog-breed formation". PLOS ONE. 15 (10): e0240787. Bibcode:2020PLoSO..1540787S. doi:10.1371/journal.pone.0240787. PMC 7595415. PMID 33119634.
- Pörtl, Daniela; Jung, Christoph (2019). "Physiological pathways to rapid prosocial evolution" (PDF). Biologia Futura. 70 (2): 93–102. doi:10.1556/019.70.2019.12. PMID 34554422. S2CID 201392789. Archived (PDF) from the original on 21 January 2022. Retrieved 3 December 2021.
- Palagi, Elisabetta; Cordoni, Giada (2020). "Intraspecific Motor and Emotional Alignment in Dogs and Wolves: The Basic Building Blocks of Dog–Human Affective Connectedness". Animals. 10 (2): 241. doi:10.3390/ani10020241. PMC 7070632. PMID 32028648.
- Pongrácz, Péter (2019). "The future of biology from a canine perspective". Biologia Futura. 70 (2): 89–92. doi:10.1556/019.70.2019.11. PMID 34554423.
- Hare, Brian (2013). The Genius of Dogs. Penguin Publishing Group. p. 60.
- Hare, Brian (2005). "Human-like social skills in dogs?". Trends in Cognitive Sciences. 9 (9): 439–44. doi:10.1016/j.tics.2005.07.003. PMID 16061417. S2CID 9311402.
- Lakatos, G. (2009). "A comparative approach to dogs' (Canis familiaris) and human infants' comprehension of various forms of pointing gestures". Animal Cognition. 12 (4): 621–31. doi:10.1007/s10071-009-0221-4. PMID 19343382. S2CID 18078591.
- Muller, C. (2015). "Dogs can discriminate the emotional expressions of human faces". Current Biology. 25 (5): 601–5. doi:10.1016/j.cub.2014.12.055. PMID 25683806.
- Hare, Brian (2013). "What Are Dogs Saying When They Bark?". Scientific American. Archived from the original on 15 March 2015. Retrieved 17 March 2015.
- Sanderson, K. (2008). "Humans can judge a dog by its growl". Nature. doi:10.1038/news.2008.852.
- Nagasawa, M. (2015). "Oxytocin-gaze positive loop and the coevolution of human-dog bonds". Science. 348 (6232): 333–336. Bibcode:2015Sci...348..333N. doi:10.1126/science.1261022. PMID 25883356. S2CID 5399803.
- Derr 2011, pp. 40
- Taçon, Paul; Pardoe, Colin (2002). "Dogs make us human". Nature Australia. Australian Museum. 27 (4): 52–61. Archived from the original on 30 January 2022. Retrieved 3 December 2017. also available: https://www.researchgate.net/publication/29464691_Dogs_make_us_human Archived 7 March 2017 at the Wayback Machine
- Grandin, Temple; Johnson, Catherine (2005). Animals in Translation: Using the Mysteries of Autism to Decode Animal Behavior (PDF). New York: Harcourt, Inc. p. 305. Archived (PDF) from the original on 8 January 2016. Retrieved 22 February 2016.
- Kirkpatrick, Kathryn (2014). Jeanne Dubino; Ziba Rashidian; Andrew Smyth (eds.). Representing the Modern Animal in Culture. Palgrave Macmillan New York. p. 75.
- Fogg, Brandy R.; Howe, Nimachia; Pierotti, Raymond (2015). "Relationships Between Indigenous American Peoples and Wolves 1: Wolves as Teachers and Guides". Journal of Ethnobiology. 35 (2): 262–285. doi:10.2993/etbi-35-02-262-285.1. S2CID 86236996.
- Duleba, Anna; Skonieczna, Katarzyna; Bogdanowicz, Wiesław; Malyarchuk, Boris; Grzybowski, Tomasz (2015). "Complete mitochondrial genome database and standardized classification system for Canis lupus familiaris". Forensic Science International: Genetics. 19: 123–129. doi:10.1016/j.fsigen.2015.06.014. PMID 26218982.
- Nikolskiy, P. A.; Sotnikova, M.V.; Nikolskii, A.A.; Pitulko, V.V. (2018). "Predomestication and Wolf-Human Relationships in the Arctic Siberia of 30,000 Years Ago: Evidence from the Yana Palaeolithic Site". Stratum Plus (1): 231–262. Archived from the original on 14 April 2021. Retrieved 24 February 2021.
- Schweizer, Rena M; Durvasula, Arun; Smith, Joel; Vohr, Samuel H; Stahler, Daniel R; Galaverni, Marco; Thalmann, Olaf; Smith, Douglas W; Randi, Ettore; Ostrander, Elaine A; Green, Richard E; Lohmueller, Kirk E; Novembre, John; Wayne, Robert K (2018). "Natural Selection and Origin of a Melanistic Allele in North American Gray Wolves". Molecular Biology and Evolution. 35 (5): 1190–1209. doi:10.1093/molbev/msy031. PMC 6455901. PMID 29688543.
- Wang, Ming-Shan; Wang, Sheng; Li, Yan; Jhala, Yadvendradev; Thakur, Mukesh; Otecko, Newton O.; Si, Jing-Fang; Chen, Hong-Man; Shapiro, Beth; Nielsen, Rasmus; Zhang, Ya-Ping; Wu, Dong-Dong (2020). "Ancient Hybridization with an Unknown Population Facilitated High-Altitude Adaptation of Canids". Molecular Biology and Evolution. 37 (9): 2616–2629. doi:10.1093/molbev/msaa113. PMID 32384152.
- Janssens, Luc; Giemsch, Liane; Schmitz, Ralf; Street, Martin; Van Dongen, Stefan; Crombé, Philippe (2018). "A new look at an old dog: Bonn-Oberkassel reconsidered". Journal of Archaeological Science. 92: 126–138. doi:10.1016/j.jas.2018.01.004. hdl:1854/LU-8550758. Archived from the original on 24 February 2021. Retrieved 9 February 2020.
- Bryce, Authors: Caleb M.; Davis, Michael S.; Gompper, Matthew E.; Hurt, Aimee; Koster, Jeremy M.; Larson, Greger; Ostrander, Elaine A.; Udell, Monique A R.; Urfer, Silvan; Wirsing, Aaron J.; Jimenez, Ana G. (2021). "Biology's best friend: Bridging disciplinary gaps to advance canine science". Integrative and Comparative Biology. doi:10.1093/icb/icab072.
- Verworn, M.; Bonnet, R.; Steinmann, G. (1914). "Diluviale Menschenfunde in Obercassel bei Bonn" [Diluvial People found in Obercassel near Bonn]. Naturwissenschaften. 2 (27): 645–650. Bibcode:1914NW......2..645V. doi:10.1007/bf01495289. S2CID 34155217.
- Verworn, M., R. Bonnet, G. Steinmann. 1919. Der diluviale Menschenfund von Obercassel bei Bonn. Wiesbaden. [The diluvial People found in Obercassel near Bonn]
- Nobis, G. 1979. Der älteste Haushund lebte vor 14 000 Jahren. Umschau 79 (19): 610.
- Nobis, G. 1981. Aus Bonn: Das älteste Haustier des Menschen. Unterkiefer eines Hundes aus dem Magdaleniengrab von Bonn-Oberkassel. Das Rheinische Landesmuseum Bonn: Berichte aus der Arbeit des Museums 4/81: 49–50.
- Benecke, Norbert (1987). "Studies on early dog remains from Northern Europe". Journal of Archaeological Science. 14: 31–49. doi:10.1016/S0305-4403(87)80004-3.
- Street, Martin & Janssens, Luc & Napierala, Hannes. (2015). Street, M., Napierala, H. & Janssens, L. 2015: The late Palaeolithic dog from Bonn-Oberkassel in context. In: The Late Glacial Burial from Oberkassel Revisited (L. Giemsch / R. W. Schmitz eds.), Rheinische Ausgrabungen 72, 253-274. ISBN 978-3-8053-4970-3. Rheinische Ausgrabungen.
- Feuerborn, Tatiana R.; Carmagnini, Alberto; Losey, Robert J.; Nomokonova, Tatiana; Askeyev, Arthur; Askeyev, Igor; Askeyev, Oleg; Antipina, Ekaterina E.; Appelt, Martin; Bachura, Olga P.; Beglane, Fiona; Bradley, Daniel G.; Daly, Kevin G.; Gopalakrishnan, Shyam; Murphy Gregersen, Kristian; Guo, Chunxue; Gusev, Andrei V.; Jones, Carleton; Kosintsev, Pavel A.; Kuzmin, Yaroslav V.; Mattiangeli, Valeria; Perri, Angela R.; Plekhanov, Andrei V.; Ramos-Madrigal, Jazmín; Schmidt, Anne Lisbeth; Shaymuratova, Dilyara; Smith, Oliver; Yavorskaya, Lilia V.; Zhang, Guojie; Willerslev, Eske; Meldgaard, Morten; Gilbert, M. Thomas P.; Larson, Greger; Dalén, Love; Hansen, Anders J.; Sinding, Mikkel-Holger S.; Frantz, Laurent (2021). "Modern Siberian dog ancestry was shaped by several thousand years of Eurasian-wide trade and human dispersal". Proceedings of the National Academy of Sciences. 118 (39): e2100338118. Bibcode:2021PNAS..11800338F. doi:10.1073/pnas.2100338118. PMC 8488619. PMID 34544854. S2CID 237584023.
- Bergström, Anders; et al. (2022). "Grey wolf genomic history reveals a dual ancestry of dogs". Nature. 607: 313–320. doi:10.1038/s41586-022-04824-9.
- Coppinger R; Feinstein M (2015). How Dogs Work. University of Chicago Press, Chicago. ISBN 978-0-226-12813-9.
- Davis, S (1982). "The taming of the few". New Scientist. 95: 697–700.
- Clutton-Brock, J. 1984. Dog, in I.L. Mason (ed.) Evolution of domesticated animals. London:Longman.
- Perri, Angela R. (2016). "Hunting dogs as environmental adaptations in Jōmon Japan". Antiquity. 90 (353): 1166–1180. doi:10.15184/aqy.2016.115.
- Guagnin, Maria; Perri, Angela R.; Petraglia, Michael D. (2018). "Pre-Neolithic evidence for dog-assisted hunting strategies in Arabia". Journal of Anthropological Archaeology. 49: 225–236. doi:10.1016/j.jaa.2017.10.003. hdl:10072/412752. S2CID 149042460. Archived from the original on 22 October 2020. Retrieved 27 August 2020.
- Sinding, Mikkel-Holger S.; Gopalakrishnan, Shyam; Ramos-Madrigal, Jazmín; De Manuel, Marc; Pitulko, Vladimir V.; Kuderna, Lukas; Feuerborn, Tatiana R.; Frantz, Laurent A. F.; Vieira, Filipe G.; Niemann, Jonas; Samaniego Castruita, Jose A.; Carøe, Christian; Andersen-Ranberg, Emilie U.; Jordan, Peter D.; Pavlova, Elena Y.; Nikolskiy, Pavel A.; Kasparov, Aleksei K.; Ivanova, Varvara V.; Willerslev, Eske; Skoglund, Pontus; Fredholm, Merete; Wennerberg, Sanne Eline; Heide-Jørgensen, Mads Peter; Dietz, Rune; Sonne, Christian; Meldgaard, Morten; Dalén, Love; Larson, Greger; Petersen, Bent; et al. (2020). "Arctic-adapted dogs emerged at the Pleistocene–Holocene transition". Science. 368 (6498): 1495–1499. Bibcode:2020Sci...368.1495S. doi:10.1126/science.aaz8599. PMC 7116267. PMID 32587022. S2CID 220072941. Archived from the original on 14 April 2021. Retrieved 4 July 2020.
- Pitulko, Vladimir V.; Kasparov, Aleksey K. (2017). "Archaeological dogs from the Early Holocene Zhokhov site in the Eastern Siberian Arctic". Journal of Archaeological Science: Reports. 13: 491–515. doi:10.1016/j.jasrep.2017.04.003.
- Da Silva Coelho, Flavio Augusto; Gill, Stephanie; Tomlin, Crystal M.; Heaton, Timothy H.; Lindqvist, Charlotte (2021). "An early dog from southeast Alaska supports a coastal route for the first dog migration into the Americas". Proceedings of the Royal Society B: Biological Sciences. 288 (1945). doi:10.1098/rspb.2020.3103. PMC 7934960. PMID 33622130. S2CID 232020982.
- Perri, Angela; Widga, Chris; Lawler, Dennis; Martin, Terrance; Loebel, Thomas; Farnsworth, Kenneth; Kohn, Luci; Buenger, Brent (2019). "New Evidence of the Earliest Domestic Dogs in the Americas". American Antiquity. 84: 68–87. doi:10.1017/aaq.2018.74.
- Ní Leathlobhair, Máire; Perri, Angela R; Irving-Pease, Evan K; Witt, Kelsey E; Linderholm, Anna; Haile, James; Lebrasseur, Ophelie; Ameen, Carly; Blick, Jeffrey; Boyko, Adam R; Brace, Selina; Cortes, Yahaira Nunes; Crockford, Susan J; Devault, Alison; Dimopoulos, Evangelos A; Eldridge, Morley; Enk, Jacob; Gopalakrishnan, Shyam; Gori, Kevin; Grimes, Vaughan; Guiry, Eric; Hansen, Anders J; Hulme-Beaman, Ardern; Johnson, John; Kitchen, Andrew; Kasparov, Aleksei K; Kwon, Young-Mi; Nikolskiy, Pavel A; Lope, Carlos Peraza; et al. (2018). "The evolutionary history of dogs in the Americas". Science. 361 (6397): 81–85. Bibcode:2018Sci...361...81N. doi:10.1126/science.aao4776. PMC 7116273. PMID 29976825.
- Ameen, Carly; Feuerborn, Tatiana R.; Brown, Sarah K.; Linderholm, Anna; Hulme-Beaman, Ardern; Lebrasseur, Ophélie; Sinding, Mikkel-Holger S.; Lounsberry, Zachary T.; Lin, Audrey T.; Appelt, Martin; Bachmann, Lutz; Betts, Matthew; Britton, Kate; Darwent, John; Dietz, Rune; Fredholm, Merete; Gopalakrishnan, Shyam; Goriunova, Olga I.; Grønnow, Bjarne; Haile, James; Hallsson, Jón Hallsteinn; Harrison, Ramona; Heide-Jørgensen, Mads Peter; Knecht, Rick; Losey, Robert J.; Masson-Maclean, Edouard; McGovern, Thomas H.; McManus-Fry, Ellen; Meldgaard, Morten; et al. (2019). "Specialized sledge dogs accompanied Inuit dispersal across the North American Arctic". Proceedings of the Royal Society B: Biological Sciences. 286 (1916): 20191929. doi:10.1098/rspb.2019.1929. PMC 6939252. PMID 31771471.
- Skoglund, Pontus; Ersmark, Erik; Palkopoulou, Eleftheria; Dalén, Love (2015). "Ancient Wolf Genome Reveals an Early Divergence of Domestic Dog Ancestors and Admixture into High-Latitude Breeds". Current Biology. 25 (11): 1515–1519. doi:10.1016/j.cub.2015.04.019. PMID 26004765.
- Ramos-Madrigal, Jazmín; Sinding, Mikkel-Holger S.; Carøe, Christian; Mak, Sarah S.T.; Niemann, Jonas; Samaniego Castruita, José A.; Fedorov, Sergey; Kandyba, Alexander; Germonpré, Mietje; Bocherens, Hervé; Feuerborn, Tatiana R.; Pitulko, Vladimir V.; Pavlova, Elena Y.; Nikolskiy, Pavel A.; Kasparov, Aleksei K.; Ivanova, Varvara V.; Larson, Greger; Frantz, Laurent A.F.; Willerslev, Eske; Meldgaard, Morten; Petersen, Bent; Sicheritz-Ponten, Thomas; Bachmann, Lutz; Wiig, Øystein; Hansen, Anders J.; Gilbert, M. Thomas P.; Gopalakrishnan, Shyam (2021). "Genomes of Pleistocene Siberian Wolves Uncover Multiple Extinct Wolf Lineages". Current Biology. 31 (1): 198–206.e8. doi:10.1016/j.cub.2020.10.002. PMC 7809626. PMID 33125870.
- Walker, Brett (2008). The Lost Wolves of Japan. University of Washington Press, Seattle. p. 51. ISBN 978-0-295-98814-6. Archived from the original on 15 April 2021. Retrieved 8 October 2022.
- Ikeya, K. 1994. Hunting with dogs among the San in the Central Kalahari. African Study Monographs 15:119–34.
- Gron, O. & M.G. Turov. 2007. Resource 'pooling' and resource management. An ethno-archaeological study of the Evenk hunter-gatherers, Katanga County, Siberia, in B. Hardh, K. Jennbert & D. Olausson (ed.) On the road: studies in honour of Lars Larsson (Acta Archaeologica Lundensia 26):67–72. Stockholm: Almqvist & Wiksell.
- Koler-Matznick, Janice; Brisbin, I. Lehr Jr.; Yates, S; Bulmer, Susan (2007). "The New Guinea singing dog: its status and scientific importance". The Journal of the Australian Mammal Society. 29 (1): 47–56. CiteSeerX 10.1.1.627.5761. doi:10.1071/AM07005.
- Olowo Ojoade, J. 1990. Nigerian cultural attitudes to the dog, in R. Willis (ed.) Signifying animals: human meaning in the natural world: 215–21. London: Routledge.
- Mizoguchi, K. 2002. An archaeological history of Japan: 10,000 B.C. to A.D. 700. Philadelphia: University of Pennsylvania Press.
- Bourque, B.J. 1975. Comments on the late Archaic populations of central Maine: the view from the urner Farm. Arctic Anthropology 12: 35–45.
- Larsson, L. 1990. Dogs in fraction—symbols in action, in P.M. Vermeersch & P. Van Peer (ed.) Contributions to the Mesolithic in Europe: 153–60. Leuven: Leuven University Press.
- Morey, D.F. (1992). "Early Holocene domestic dog burials from the North American Midwest". Current Anthropology. 33 (2): 224–29. doi:10.1086/204059. S2CID 144485292.
- Gautier, Achilles (2001). "The Early to Late Neolithic Archeofaunas from Nabta and Bir Kiseiba". Holocene Settlement of the Egyptian Sahara. pp. 609–635. doi:10.1007/978-1-4615-0653-9_23. ISBN 978-1-4613-5178-8. Refer to page 620.
- Clutton, Juliet; Driscoll, Carlos A. (2016). "1-Origins of the dog:The archaeological evidence". In James Serpell (ed.). The Domestic Dog: Its Evolution, Behavior and Interactions with People (2 ed.). Cambridge University Press. p. 17. ISBN 978-1-107-02414-4. Archived from the original on 8 October 2022. Retrieved 29 May 2020.
- Zhang, Ming; Sun, Guoping; Ren, Lele; Yuan, Haibing; Dong, Guanghui; Zhang, Lizhao; Liu, Feng; Cao, Peng; Ko, Albert Min-Shan; Yang, Melinda A.; Hu, Songmei; Wang, Guo-Dong; Fu, Qiaomei (2020). "Ancient DNA evidence from China reveals the expansion of Pacific dogs". Molecular Biology and Evolution. 37 (5): 1462–1469. doi:10.1093/molbev/msz311. PMC 7182212. PMID 31913480.
- Pierotti & Fogg 2017, pp. 5–6
Bibliography
- Derr, Mark (2011). How the Dog Became the Dog: From Wolves to Our Best Friends. Penguin Group. ISBN 978-1-4683-0269-1.
- Pierotti, R.; Fogg, B. (2017). The First Domestication: How Wolves and Humans Coevolved. Yale University Press. ISBN 978-0-300-22616-4.
- Shipman, P. (2015). The Invaders:How humans and their dogs drove Neanderthals to extinction. Harvard University Press. ISBN 978-0-674-73676-4.
| Types |
|
|---|---|
| Breeds |
|
| Roles |
|
| Behavior |
|
| Human–dog interaction |
|
| Health |
|
| Training |
|
| Related |
|
| |