Styracosaurus
Styracosaurus (/stɪˌrækəˈsɔːrəs/ sti-RAK-ə-SOR-əs; meaning "spiked lizard" from the Ancient Greek styrax/στύραξ "spike at the butt-end of a spear-shaft" and sauros/σαῦρος "lizard")[1] is a genus of herbivorous ceratopsian dinosaur from the Cretaceous Period (Campanian stage), about 75.5 to 74.5 million years ago. It had four to six long parietal spikes extending from its neck frill, a smaller jugal horn on each of its cheeks, and a single horn protruding from its nose, which may have been up to 60 centimeters (2 feet) long and 15 centimeters (6 inches) wide. The function or functions of the horns and frills have been debated for many years.
| Styracosaurus Temporal range: Late Cretaceous (Campanian), | |
|---|---|
 | |
| Holotype skeleton, Canadian Museum of Nature | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Suborder: | †Ceratopsia |
| Family: | †Ceratopsidae |
| Subfamily: | †Centrosaurinae |
| Clade: | †Eucentrosaura |
| Tribe: | †Centrosaurini |
| Genus: | †Styracosaurus Lambe, 1913 |
| Type species | |
| †Styracosaurus albertensis Lambe, 1913 | |
| Other species | |
| |
| Synonyms | |
S. albertensis synonymy
S. ovatus synonymy
| |
Styracosaurus was a relatively large dinosaur, reaching lengths of 5–5.5 metres (16–18 ft) and weighing about 1.8–2.7 metric tons (2.0–3.0 short tons). It stood about 1.8 meters (5.9 feet) tall. Styracosaurus possessed four short legs and a bulky body. Its tail was rather short. The skull had a beak and shearing cheek teeth arranged in continuous dental batteries, suggesting that the animal sliced up plants. Like other ceratopsians, this dinosaur may have been a herd animal, travelling in large groups, as suggested by bone beds.
Named by Lawrence Lambe in 1913, Styracosaurus is a member of the Centrosaurinae. One species, S. albertensis, is currently assigned to Styracosaurus. Another species, S. ovatus, named in 1930 by Charles Gilmore was reassigned to a new genus, Rubeosaurus, by Andrew McDonald and Jack Horner in 2010,[2] but it has been considered either its own species or a species of Styracosaurus (or even a specimen of S. albertensis)[3] again, since 2020.
Discoveries and species
_(20765048851).jpg.webp)
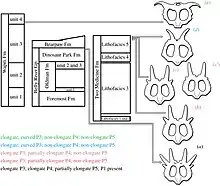
The first fossil remains of Styracosaurus were collected in Alberta, Canada by C. M. Sternberg (from an area now known as Dinosaur Provincial Park, in a formation now called the Dinosaur Park Formation) and named by Lawrence Lambe in 1913. This quarry was revisited in 1935 by a Royal Ontario Museum crew who found the missing lower jaws and most of the skeleton. These fossils indicate that S. albertensis was around 5.5–5.8 metres (18–19 ft) in length and stood about 1.65 metres (5.4 ft) high at the hips.[4] An unusual feature of this first skull is that the smallest frill spike on the left side is partially overlapped at its base by the next spike. It appears that the frill suffered a break at this point in life and was shortened by about 6 centimeters (2.4 inches). The normal shape of this area is unknown because the corresponding area of the right side of the frill was not recovered.[5]

Barnum Brown and crew, working for the American Museum of Natural History in New York, collected a nearly complete articulated skeleton with a partial skull in 1915. These fossils were also found in the Dinosaur Park Formation, near Steveville, Alberta. Brown and Erich Maren Schlaikjer compared the finds, and, though they allowed that both specimens were from the same general locality and geological formation, they considered the specimen sufficiently distinct from the holotype to warrant erecting a new species, and described the fossils as Styracosaurus parksi, named in honor of William Parks.[6] Among the differences between the specimens cited by Brown and Schlaikjer were a cheekbone quite different from that of S. albertensis, and smaller tail vertebrae. S. parksi also had a more robust jaw, a shorter dentary, and the frill differed in shape from that of the type species.[6] However, much of the skull consisted of plaster reconstruction, and the original 1937 paper did not illustrate the actual skull bones.[4] It is now accepted as a specimen of S. albertensis.[5][7]
In the summer of 2006, Darren Tanke of the Royal Tyrrell Museum of Palaeontology in Drumheller, Alberta relocated the long lost S. parksi site.[5] Pieces of the skull, evidently abandoned by the 1915 crew, were found in the quarry. These were collected and it is hoped more pieces will be found, perhaps enough to warrant a redescription of the skull and test whether S. albertensis and S. parksi are the same. The Tyrrell Museum has also collected several partial Styracosaurus skulls.[8] At least one confirmed bone bed (bonebed 42) in Dinosaur Provincial Park has also been explored (other proposed Styracosaurus bone beds instead have fossils from a mix of animals, and nondiagnostic ceratopsian remains). Bonebed 42 is known to contain numerous pieces of skulls such as horncores, jaws and frill pieces.[5]
Several other species which were assigned to Styracosaurus have since been assigned to other genera. S. sphenocerus, described by Edward Drinker Cope in 1890 as a species of Monoclonius and based on a nasal bone with a broken Styracosaurus-like straight nose horn, was attributed to Styracosaurus in 1915.[9] "S. makeli", mentioned informally by amateur paleontologists Stephen and Sylvia Czerkas in 1990 in a caption to an illustration, is an early name for Einiosaurus.[10] "S. borealis" is an early informal name for S. parksi.[11]
Styracosaurus ovatus

A species, Styracosaurus ovatus, from the Two Medicine Formation of Montana, was described by Gilmore in 1930, named for a partial parietal under the accession number USNM 11869. Unlike S. albertensis, the longest parietal spikes converge towards their tips, instead of projecting parallel behind the frill. There also may only have been two sets of spikes on each side of the frill, instead of three. As estimated from the preserved material, the spikes are much shorter than in S. albertensis, with the longest only 295 millimeters (11.6 inches) long.[12] An additional specimen from the Two Medicine Formation was referred to Styracosaurus ovatus in 2010 by Andrew McDonald and John Horner, having been found earlier in 1986 but not described until that year. Known from a premaxilla, the nasal bones and their horncore, a postorbital bone and a parietal, the specimen Museum of the Rockies 492 was considered to share the medially-converging parietal spikes with the only other specimen of S. ovatus, the holotype. Following this additional material, the species was added to a phylogenetic analysis where it was found to group not with Styracosaurus albertensis, but in a clade including Pachyrhinosaurus, Einiosaurus and Achelousaurus, and therefore McDonald and Horner gave the species the new genus name Rubeosaurus.[13] Another specimen, the partial immature skull USNM 14768, which was earlier referred to the undiagnostic genus Brachyceratops, was also referred to Rubeosaurus ovatus by McDonald and colleagues in 2011. While the medial spikes of USNM 14768 were too incomplete to show if it shared the convergence seen in other R. ovatus specimens, it was considered to be the same species as it was also found in the older deposits of the Two Medicine Formation, and had a unique combination of parietal features only shared completely with the other specimens of the species.[14]
Though it was originally found to nest closer to Einiosaurus and later centrosaurines by McDonald and colleagues in both 2010 and 2011, revisions of phylogenetic analyses in 2013 by Scott Sampson and colleagues, and further expansions and modifications of the same dataset, instead placed Rubeosaurus ovatus as the sister taxon of Styracosaurus albertensis, as had been originally considered when the species was first named, though the two species were not moved into the same genus as originally named. A review of the variability within known Styracosaurus specimens by Robert Holmes and colleagues in 2020 found that USNM 11869, the type specimen of Rubeosaurus ovatus, fell within the variation seen in other specimens from the older deposits of the Dinosaur Park Formation S. albertensis is known from. While no phylogenetic analysis was conducted, previous results of updated analyses showed that Rubeosaurus ovatus and Styracosaurus albertensis were not distantly related, so the justification for naming the genus Rubeosaurus was not present, and the variability in Styracosaurus albertensis specimens also did not support the distinction of Styracosaurus ovatus, with Holmes et al. considering the latter a junior synonym of the former.[3] The conclusion of Holmes and colleagues was supported by a later 2020 study authored by Caleb Brown, Holmes, and Philip J. Currie, who described a new juvenile Styracosaurus specimen and determined that there were several specimens that are otherwise consistent with S. albertensis have been found with inward angled midline frill spikes, though not the same degree as S. ovatus. Though they considered that S. ovatus represented an extreme end of the S. albertensis variation not only in morphology but also as it was stratigraphically younger, they cautioned that at the least the current diagnosis of S. ovatus was inadequate.[15]
Later in 2020, the supposed specimen MOR 492 was redescribed by John Wilson and colleagues, who reinterpreted its anatomy in a way that contrasted McDonald and Horner who referred it to Styracosaurus ovatus. While Wilson et al. agreed that the close relationship between S. albertensis and S. ovatus meant that the genus name Rubeosaurus should be abandoned, they cautioned against synonymization. MOR 492 was moved into its own taxon, Stellasaurus ancellae, which nested alongside Einiosaurus, Achelousaurus and Pachyrhinosaurus in a similar result to McDonald and Horner when the specimen was included as part of the S. ovatus hypodigm. Wilson and colleagues also suggested that the new taxon may have been ancestral to the later forms it was found related to, suggesting that gradual evolution through anagenesis could be the reason for the intermediate morphologies of many specimens and species found in the Two Medicine Formation, possibly also including S. ovatus. As the holotype of Styracosaurus ovatus was found in deposits much younger than the remainder of Styracosaurus specimens, and was considered to have the most extreme morphology while still falling within plausible variation as Holmes et al. had concluded, Wilson and colleagues advised that S. ovatus was retained as a separate, probably directly descended from S. albertensis, species of Styracosaurus. The immature specimen USNM 14768, referred to S. ovatus by McDonald et al. in 2011, was considered too immature to be diagnostic, and thus S. ovatus was limited to its holotype USNM 11869.[16]
Description
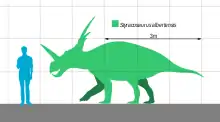
Individuals of the genus Styracosaurus were approximately 5–5.5 metres (16–18 ft) long as adults and weighed about 1.8–2.7 metric tons (2.0–3.0 short tons).[17][18] The skull was massive, with a large nostril, a tall straight nose horn, and a parietal squamosal frill (a neck frill) crowned with at least four large spikes. Each of the four longest frill spines was comparable in length to the nose horn, at 50 to 55 centimeters (20 to 22 inches) long.[4] The nasal horn was estimated by Lambe at 57 centimeters (22 inches) long in the type specimen,[19] but the tip had not been preserved. Based on other nasal horn cores from Styracosaurus and Centrosaurus, this horn may have come to a more rounded point at around half of that length.[5]

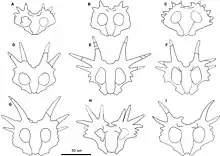
Aside from the large nasal horn and four long frill spikes, the cranial ornamentation was variable. Some individuals had small hook-like projections and knobs at the posterior margin of the frill, similar to but smaller than those in Centrosaurus. Others had less prominent tabs. Some, like the type individual, had a third pair of long frill spikes. Others had much smaller projections, and small points are found on the side margins of some but not all specimens. Modest pyramid-shaped brow horns were present in subadults, but were replaced by pits in adults.[5] Like most ceratopsids, Styracosaurus had large fenestrae (skull openings) in its frill. The front of the mouth had a toothless beak.
The bulky body of Styracosaurus resembled that of a rhinoceros. It had powerful shoulders which may have been useful in intraspecies combat. Styracosaurus had a relatively short tail. Each toe bore a hooflike ungual which was sheathed in horn.[17]
Various limb positions have been proposed for Styracosaurus and ceratopsids in general, including forelegs which were held underneath the body, or, alternatively, held in a sprawling position. The most recent work has put forward an intermediate crouched position as most likely.[20]
Classification
Styracosaurus is a member of the Centrosaurinae. Other members of the clade include Centrosaurus (from which the group takes its name),[21][22] Pachyrhinosaurus,[21][23] Avaceratops,[21] Einiosaurus,[23][24] Albertaceratops,[24] Achelousaurus,[23] Brachyceratops,[7] and Monoclonius,[21] although these last two are dubious. Because of the variation between species and even individual specimens of centrosaurines, there has been much debate over which genera and species are valid, particularly whether Centrosaurus and/or Monoclonius are valid genera, undiagnosable, or possibly members of the opposite sex. In 1996, Peter Dodson found enough variation between Centrosaurus, Styracosaurus, and Monoclonius to warrant separate genera, and that Styracosaurus resembled Centrosaurus more closely than either resembled Monoclonius. Dodson also believed one species of Monoclonius, M. nasicornis, may actually have been a female Styracosaurus.[25] However, most other researchers have not accepted Monoclonius nasicornis as a female Styracosaurus, instead regarding it as a synonym of Centrosaurus apertus.[5][26] While sexual dimorphism has been proposed for an earlier ceratopsian, Protoceratops,[27] there is no firm evidence for sexual dimorphism in any ceratopsid.[28][29][30]

_(20758118955).jpg.webp)
The cladogram depicted below represents a phylogenetic analysis by Chiba et al. (2017):[31]
| Centrosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Origins and evolution
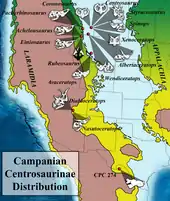
The evolutionary origins of Styracosaurus were not understood for many years because fossil evidence for early ceratopsians was sparse. The discovery of Protoceratops, in 1922, shed light on early ceratopsid relationships,[32] but several decades passed before additional finds filled in more of the blanks. Fresh discoveries in the late 1990s and 2000s, including Zuniceratops, the earliest known ceratopsian with brow horns, and Yinlong, the first-known Jurassic ceratopsian, indicate what the ancestors of Styracosaurus may have looked like. These new discoveries have been important in illuminating the origins of horned dinosaurs in general, and suggest that the group originated during the Jurassic in Asia, with the appearance of true horned ceratopsians occurring by the beginning of the late Cretaceous in North America.[7]
Goodwin and colleagues proposed in 1992 that Styracosaurus was part of the lineage leading to Einiosaurus, Achelousaurus and Pachyrhinosaurus. This was based on a series of fossil skulls from the Two Medicine Formation of Montana.[33] The position of Styracosaurus in this lineage is now equivocal, as the remains that were thought to represent Styracosaurus have been transferred to the genus Rubeosaurus.[13]
Styracosaurus is known from a higher position in the formation (relating specifically to its own genus) than the closely related Centrosaurus, suggesting that Styracosaurus displaced Centrosaurus as the environment changed over time and/or dimension.[26] It has been suggested that Styracosaurus albertensis is a direct descendant of Centrosaurus (C. apertus or C. nasicornis), and that it in turn evolved directly into the slightly later species Rubeosaurus ovatus. Subtle changes can be traced in the arrangement of the horns through this lineage, leading from Rubeosaurus to Einiosaurus, to Achelousaurus and Pachyrhinosaurus. However, the lineage may not be a simple, straight line, as a pachyrhinosaur-like species has been reported from the same time and place as Styracosaurus albertensis.[2]
In 2020, during the description of Stellasaurus, Wilson et al. found Styracosaurus (including S. ovatus) to be the earliest member of a single evolutionary lineage that eventually developed into Stellasaurus, Achelousaurus, and Pachyrhinosaurus.[14]
Paleobiology

Styracosaurus and other horned dinosaurs are often depicted in popular culture as herd animals. A bonebed composed of Styracosaurus remains is known from the Dinosaur Park Formation of Alberta, about halfway up the formation. This bonebed is associated with different types of river deposits.[8][34] The mass deaths may have been a result of otherwise non-herding animals congregating around a waterhole in a period of drought, with evidence suggesting the environment may have been seasonal and semi-arid.[35]
Paleontologists Gregory Paul and Per Christiansen proposed that large ceratopsians such as Styracosaurus were able to run faster than an elephant, based on possible ceratopsian trackways which did not exhibit signs of sprawling forelimbs.[36]
Dentition and diet
Styracosaurs were herbivorous dinosaurs; they probably fed mostly on low growth because of the position of the head. They may, however, have been able to knock down taller plants with their horns, beak, and bulk.[7][37] The jaws were tipped with a deep, narrow beak, believed to have been better at grasping and plucking than biting.[38]
Ceratopsid teeth, including those of Styracosaurus, were arranged in groups called batteries. Older teeth on top were continually replaced by the teeth underneath them. Unlike hadrosaurids, which also had dental batteries, ceratopsid teeth sliced but did not grind.[7] Some scientists have suggested that ceratopsids like Styracosaurus ate palms and cycads,[39] while others have suggested ferns.[40] Dodson has proposed that Late Cretaceous ceratopsians may have knocked down angiosperm trees and then sheared off leaves and twigs.[41]
Horns and frill

The large nasal horns and frills of Styracosaurus are among the most distinctive facial adornments of all dinosaurs. Their function has been the subject of debate since the first horned dinosaurs were discovered.
Early in the 20th century, paleontologist R. S. Lull proposed that the frills of ceratopsian dinosaurs acted as anchor points for their jaw muscles.[42] He later noted that for Styracosaurus, the spikes would have given it a formidable appearance.[43] In 1996, Dodson supported the idea of muscle attachments in part and created detailed diagrams of possible muscle attachments in the frills of Styracosaurus and Chasmosaurus, but did not subscribe to the idea that they completely filled in the fenestrae.[44] C. A. Forster, however, found no evidence of large muscle attachments on the frill bones.[28]
It was long believed that ceratopsians like Styracosaurus used their frills and horns in defence against the large predatory dinosaurs of the time. Although pitting, holes, lesions, and other damage on ceratopsid skulls are often attributed to horn damage in combat, a 2006 study found no evidence for horn thrust injuries causing these forms of damage (for example, there is no evidence of infection or healing). Instead, non-pathological bone resorption, or unknown bone diseases, are suggested as causes.[45]
However, a newer study compared incidence rates of skull lesions in Triceratops and Centrosaurus and showed that these were consistent with Triceratops using its horns in combat and the frill being adapted as a protective structure, while lower pathology rates in Centrosaurus may indicate visual rather than physical use of cranial ornamentation, or a form of combat focused on the body rather than the head;[46] as Centrosaurus was more closely related to Styracosaurus and both genera had long nasal horns, the results for this genus would be more applicable for Styracosaurus. The researchers also concluded that the damage found on the specimens in the study was often too localized to be caused by bone disease.[47]
The large frill on Styracosaurus and related genera also may have helped to increase body area to regulate body temperature,[48] like the ears of the modern elephant. A similar theory has been proposed regarding the plates of Stegosaurus,[49] although this use alone would not account for the bizarre and extravagant variation seen in different members of the Ceratopsidae.[7] This observation is highly suggestive of what is now believed to be the primary function, display.
The theory of frill use in sexual display was first proposed in 1961 by Davitashvili. This theory has gained increasing acceptance.[28][50] Evidence that visual display was important, either in courtship or in other social behavior, can be seen in the fact that horned dinosaurs differ markedly in their adornments, making each species highly distinctive. Also, modern living creatures with such displays of horns and adornments use them in similar behavior.[51]
The use of the exaggerated structures in dinosaurs as species identification has been questioned, as no such function exists in vast majority of modern species of tetrapods (terrestrial vertebrates).[52]
A skull discovered in 2015 from a Styracosaurus indicates that individual variation was likely commonplace in the genus. The asymmetrical nature of the horns in the specimen has been compared to deer, which often have asymmetrical antlers in various individuals. The study carried out may also indicate that the genus Rubeosaurus may be synonymous with Styracosaurus as a result.[3]
Paleoecology

Styracosaurus is known from the Dinosaur Park Formation, and was a member of a diverse and well-documented fauna of prehistoric animals that included horned relatives such as Centrosaurus and Chasmosaurus, duckbills such as Prosaurolophus, Lambeosaurus, Gryposaurus, Corythosaurus, and Parasaurolophus, tyrannosaurids Gorgosaurus, Daspletosaurus, and armored Edmontonia and Euoplocephalus.[53]
The Dinosaur Park Formation is interpreted as a low-relief setting of rivers and floodplains that became more swampy and influenced by marine conditions over time as the Western Interior Seaway transgressed westward.[54] The climate was warmer than present-day Alberta, without frost, but with wetter and drier seasons. Conifers were apparently the dominant canopy plants, with an understory of ferns, tree ferns, and angiosperms.[55]
In popular culture
Styracosaurus was featured in the lost pit scene in "King Kong" (1933). Although the original footage is now lost, a similar, fictional ceratopsian called Ferrucutus based on Styracosaurus and Pachyrhinosaurus can be seen in Peter Jackson's recreation.
See also
- Timeline of ceratopsian research
References
- Liddell & Scott (1980). Greek-English Lexicon, Abridged Edition. Oxford University Press, Oxford, UK. ISBN 978-0-19-910207-5.
- Andrew T. McDonald & John R. Horner, (2010). "New Material of "Styracosaurus" ovatus from the Two Medicine Formation of Montana". Pages 156–168 in: Michael J. Ryan, Brenda J. Chinnery-Allgeier, and David A. Eberth (eds), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Indiana University Press, Bloomington and Indianapolis, IN.
- Holmes, R.B.; Persons, W.S.; Singh Rupal, B.; Jawad Qureshi, A.; Currie, P.J. (2020). "Morphological variation and asymmetrical development in the skull of Styracosaurus albertensis". Cretaceous Research. 107: 104308. doi:10.1016/j.cretres.2019.104308. S2CID 210260909.
- Dodson, P. (1996). The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press. pp. 165–169. ISBN 978-0-691-05900-6.
- Ryan, Michael J.; Holmes, Robert; Russell, A.P. (2007). "A revision of the late Campanian centrosaurine ceratopsid genus Styracosaurus from the Western Interior of North America" (PDF). Journal of Vertebrate Paleontology. 27 (4): 944–962. doi:10.1671/0272-4634(2007)27[944:AROTLC]2.0.CO;2. S2CID 86218327. Retrieved August 19, 2010.
- Brown, Barnum; Erich Maren Schlaikjer (1937). "The skeleton of Styracosaurus with the description of a new species". American Museum Novitates (955): 12. hdl:2246/2191.
- Dodson, P., Forster, C. A, and Sampson, S. D. (2004) Ceratopsidae. In: Weishampel, D. B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria (second edition). Berkeley: University of California Press, pp. 494–513. ISBN 0-520-24209-2.
- Eberth, David A.; Getty, Michael A. (2005). "Ceratopsian bonebeds: occurrence, origins, and significance". In Currie, Phillip J.; Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. pp. 501–536. ISBN 978-0-253-34595-0.
- Lambe, L. M. (1915). "On Eoceratops canadensis, gen. nov., with remarks on other genera of Cretaceous horned dinosaurs". Canada Geological Survey Bulletin, Geological Series. 12 (24): 1–49.
- Glut, Donald F. (1997). "Einiosaurus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 396–398. ISBN 978-0-89950-917-4.
- Glut, Donald F. (1997). "Styracosaurus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 865–868. ISBN 978-0-89950-917-4.
- Gilmore, Charles W. (1930). "On dinosaurian reptiles from the Two Medicine Formation of Montana". Proceedings of the United States National Museum. 77 (16): 1–39. doi:10.5479/si.00963801.77-2839.1.
- Andrew T. McDonald & John R. Horner, (2010). "New Material of "Styracosaurus" ovatus from the Two Medicine Formation of Montana", In: Michael J. Ryan, Brenda J. Chinnery-Allgeier, and David A. Eberth (eds), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Indiana University Press, 656 pp.
- McDonald, A. T. (2011). Farke, Andrew Allen (ed.). "A Subadult Specimen of Rubeosaurus ovatus (Dinosauria: Ceratopsidae), with Observations on Other Ceratopsids from the Two Medicine Formation". PLOS ONE. 6 (8): e22710. Bibcode:2011PLoSO...622710M. doi:10.1371/journal.pone.0022710. PMC 3154267. PMID 21853043.
- Brown, C.; Holmes, R.; Currie, P. (2020). "A subadult individual of Styracosaurus albertensis (Ornithischia: Ceratopsidae) with comments on ontogeny and intraspecific variation in Styracosaurus and Centrosaurus". Vertebrate Anatomy Morphology Palaeontology. 8 (1): 67–95. doi:10.18435/vamp29361. S2CID 218945057.
- Wilson, John P.; Ryan, Michael J.; Evans, David C. (2020). "A new, transitional centrosaurine ceratopsid from the Upper Cretaceous Two Medicine Formation of Montana and the evolution of the 'Styracosaurus-line' dinosaurs". Royal Society Open Publishing. 7 (4): 200284. Bibcode:2020RSOS....700284W. doi:10.1098/rsos.200284. PMC 7211873. PMID 32431910.
- Lambert, D. (1993). The Ultimate Dinosaur Book. Dorling Kindersley: New York, 152–167. ISBN 1-56458-304-X.
- Paul, Gregory S. (2010). Princeton Field Guide to Dinosaurs. Princeton University Press. ISBN 978-0-691-13720-9.
- Lambe, L.M. (1913). "A new genus and species from the Belly River Formation of Alberta". Ottawa Naturalist. 27: 109–116.
- Thompson, Stefan; Holmes, Robert (April 2007). "Forelimb stance and step cycle in Chasmosaurus irvinensis (Dinosauria:Neoceratopsia". Palaeontologia Electronica. Retrieved May 28, 2007.
- Dodson, P. (1990). "On the status of the ceratopsids Monoclonius and Centrosaurus". In Carpenter, K.; Currie, P.J. (eds.). Dinosaur Systematics: Perspectives and Approaches. Cambridge: Cambridge University Press. pp. 231–243. ISBN 978-0-521-36672-4.
- Ryan, M.J.; A.P. Russell (2003). "New centrosaurine ceratopsids from the late Campanian of Alberta and Montana and a review of contemporaneous and regional patterns of centrosaurine evolution". Journal of Vertebrate Paleontology. 23 (3): 91A. doi:10.1080/02724634.2003.10010538. S2CID 220410105.
- Ryan, M.J.; A.P. Russell (2005). "A new centrosaurine ceratopsid from the Oldman Formation of Alberta and its implications for centrosaurine taxonomy and systematics". Canadian Journal of Earth Sciences. 42 (7): 1369–1387. Bibcode:2005CaJES..42.1369R. doi:10.1139/e05-029. hdl:1880/47001.
- Ryan, M.J. (2007). "A new basal centrosaurine ceratopsid from the Oldman Formation, southeastern Alberta". Journal of Paleontology. 81 (2): 376–396. doi:10.1666/0022-3360(2007)81[376:ANBCCF]2.0.CO;2. S2CID 130607301.
- Dodson, P. (1996). The Horned Dinosaurs: A Natural History. Princeton University Press: Princeton, New Jersey, pp. 197–199. ISBN 0-691-02882-6.
- Ryan, Michael J.; Evans, David C. (2005). "Ornithischian Dinosaurs". In Currie, Phillip J.; Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. pp. 312–348. ISBN 978-0-253-34595-0.
- Dodson, P. "Quantitative aspects of relative growth and sexual dimorphism in Protoceratops". Journal of Paleontology. 50: 929–940.
- Forster, C. A. (1990). The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny. Ph.D. Dissertation. University of Pennsylvania, Philadelphia. 227 pp. OCLC 61500040
- Lehman, T. M. (1998). "A gigantic skull and skeleton of the horned dinosaur Pentaceratops sternbergi from New Mexico". Journal of Paleontology. 72 (5): 894–906. doi:10.1017/S0022336000027220. S2CID 132807103.
- Sampson, S. D.; Ryan, M.J.; Tanke, D.H. (1997). "Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taphonomic and behavioral phylogenetic implications". Zoological Journal of the Linnean Society. 121 (3): 293–337. doi:10.1111/j.1096-3642.1997.tb00340.x.
- Kentaro Chiba; Michael J. Ryan; Federico Fanti; Mark A. Loewen; David C. Evans (2018). "New material and systematic re-evaluation of Medusaceratops lokii (Dinosauria, Ceratopsidae) from the Judith River Formation (Campanian, Montana)". Journal of Paleontology. 92 (2): 272–288. doi:10.1017/jpa.2017.62. S2CID 134031275.
- Dodson, P. (1996). The Horned Dinosaurs: A Natural History. Princeton University Press: Princeton, New Jersey, p. 244. ISBN 0-691-02882-6.
- Goodwin, M.J.; Varricchio, D.J.; Horner, J.R. (1992). "Marine transgressions and the evolution of Cretaceous dinosaurs". Nature. 358 (6381): 59–61. Bibcode:1992Natur.358...59H. doi:10.1038/358059a0. S2CID 4283438.
- Although this article mentioned two bonebeds, including BB 156, the recent review by Ryan et al. only accepted BB 42.
- Rogers, R. R. (1990). "Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation, northwestern Montana: Evidence for drought-related mortality". PALAIOS. 5 (5): 394–41. Bibcode:1990Palai...5..394R. doi:10.2307/3514834. JSTOR 3514834.
- Paul, Gregory S; Per Christiansen (September 2000). "Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion". Paleobiology. 26 (3): 450–465. doi:10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2. S2CID 85280946.
- Tait, J.; Brown, B. (1928). "How the Ceratopsia carried and used their head". Transactions of the Royal Society of Canada. 22: 13–23.
- Ostrom, J. H. (1966). "Functional morphology and evolution of the ceratopsian dinosaurs". Evolution. 20 (3): 290–308. doi:10.2307/2406631. JSTOR 2406631. PMID 28562975.
- Weishampel, D. B. (1984). Evolution of jaw mechanisms in ornithopod dinosaurs. Advances in Anatomy Embryology and Cell Biology. Vol. 87. pp. 1–110. doi:10.1007/978-3-642-69533-9. ISBN 978-3-540-13114-4. PMID 6464809. S2CID 12547312.
- Coe, M. J., Dilcher, D. L., Farlow, J. O., Jarzen, D. M., and Russell, D. A. (1987). Dinosaurs and land plants. In: Friis, E. M., Chaloner, W. G., and Crane, P. R. (eds.) The Origins of Angiosperms and their Biological Consequences Cambridge University Press, pp. 225–258. ISBN 0-521-32357-6.
- Dodson, P. (1996). The Horned Dinosaurs: A Natural History. Princeton University Press: Princeton, New Jersey, p. 266. ISBN 0-691-02882-6.
- Lull, R.S. (1908). "The cranial musculature and the origin of the frill in the ceratopsian dinosaurs". American Journal of Science. 4 (25): 387–399. Bibcode:1908AmJS...25..387L. doi:10.2475/ajs.s4-25.149.387.
- Lull, R.S. (1933). "A revision of the Ceratopsia or horned dinosaurs". Memoirs of the Peabody Museum of Natural History. 3 (3): 1–175. doi:10.5962/bhl.title.5716.
- Dodson, P. (1996). The Horned Dinosaurs: A Natural History. Princeton University Press: Princeton, New Jersey, p. 269. ISBN 0-691-02882-6.
- Tanke, D. H, and Farke, A. A. (2006). Bone resorption, bone lesions, and extracranial fenestrae in ceratopsid dinosaurs: a preliminary assessment. in: Carpenter, K. (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs Indiana University Press: Bloomington. pp. 319–347. ISBN 0-253-34817-X.
- Farke, A. A.; Wolff, E. D. S.; Tanke, D. H.; Sereno, Paul (2009). Sereno, Paul (ed.). "Evidence of Combat in Triceratops". PLOS ONE. 4 (1): e4252. Bibcode:2009PLoSO...4.4252F. doi:10.1371/journal.pone.0004252. PMC 2617760. PMID 19172995.
- Wall, Michael (January 27, 2009). "Scars Reveal How Triceratops Fought –". Wired.com. Retrieved August 3, 2010.
- Wheeler, P.E. (1978). "Elaborate CNS cooling structures in large dinosaurs". Nature. 275 (5679): 441–443. Bibcode:1978Natur.275..441W. doi:10.1038/275441a0. PMID 692723. S2CID 4160470.
- Farlow, J. O., Thompson, C. V., and Rosner, D. E. (1976). "Plates of the dinosaur Stegosaurus: Forced convection heat loss fins?". Science. 192 (4244): 1123–1125. Bibcode:1976Sci...192.1123F. doi:10.1126/science.192.4244.1123. PMID 17748675. S2CID 44506996.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Davitashvili L (1961). The Theory of sexual selection. Izdatel'stvo Akademii Nauk SSSR. p. 538.
- Farlow, J. O. & Dodson, P. (1975). "The behavioral significance of frill and horn morphology in ceratopsian dinosaurs". Evolution. 29 (2): 353–361. doi:10.2307/2407222. JSTOR 2407222. PMID 28555861.
- Hone, D. W. E.; Naish, D. (2013). "The 'species recognition hypothesis' does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs". Journal of Zoology. 290 (3): 172–180. doi:10.1111/jzo.12035. S2CID 17827309.
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). "Dinosaur Distribution", in The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606. ISBN 0-520-24209-2
- Eberth, David A. "The geology", in Dinosaur Provincial Park, pp. 54–82.
- Braman, Dennis R., and Koppelhus, Eva B. "Campanian palynomorphs", in Dinosaur Provincial Park, pp. 101–130.
Marginocephalia | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||

.png.webp)


